
Threshold Responses of Canopy Cover and Tree Growth to Drought and Siberian silk Moth Outbreak in Southern Taiga Picea obovata Forests



Abstract
:1. Introduction
2. Materials and Methods
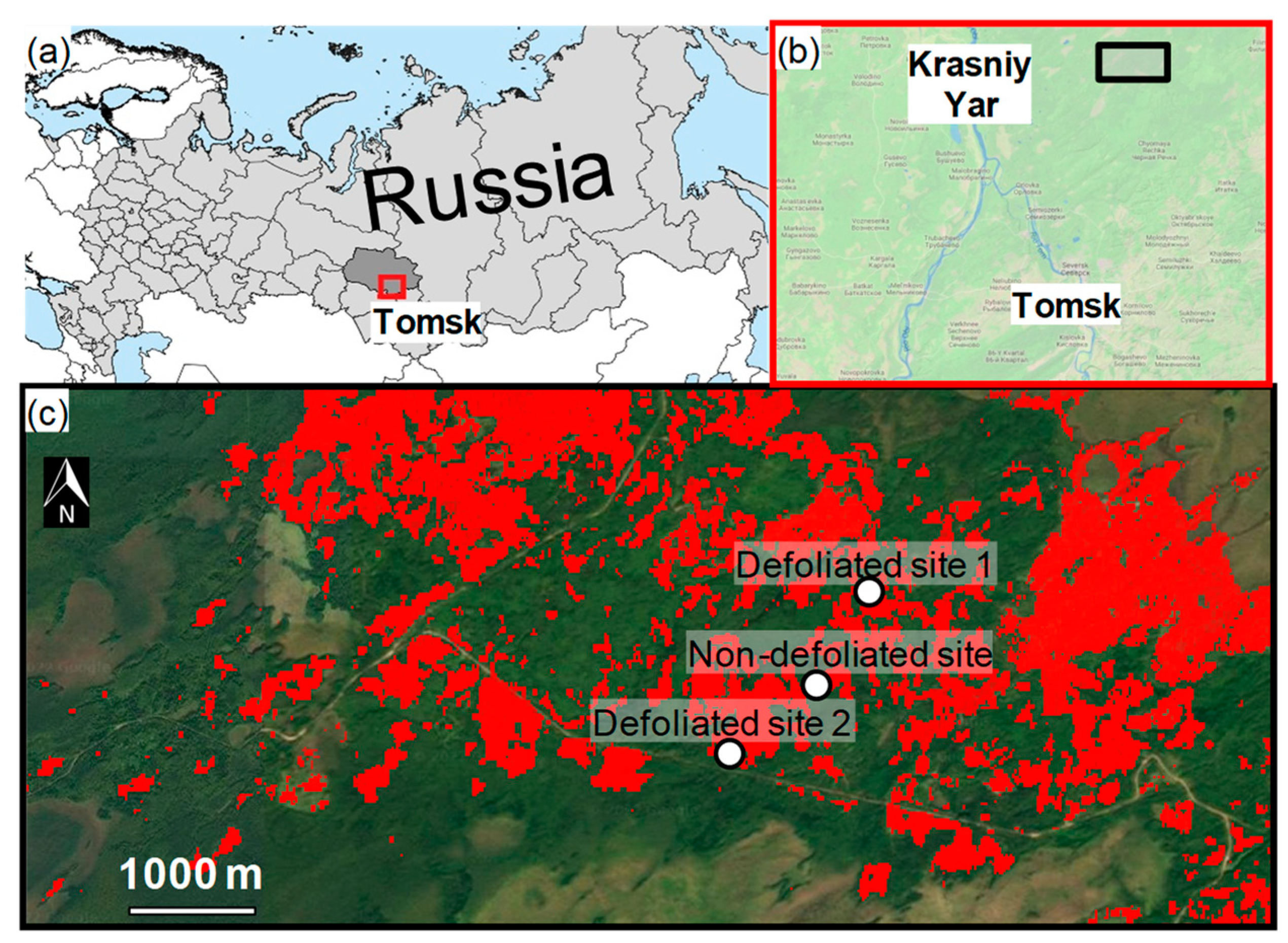
2.1. Study Sites and Species
2.2. Field Sampling
2.3. Tree-Ring Data Processing
2.4. Remote Sensing Data
2.5. Statistical Analyses
3. Results
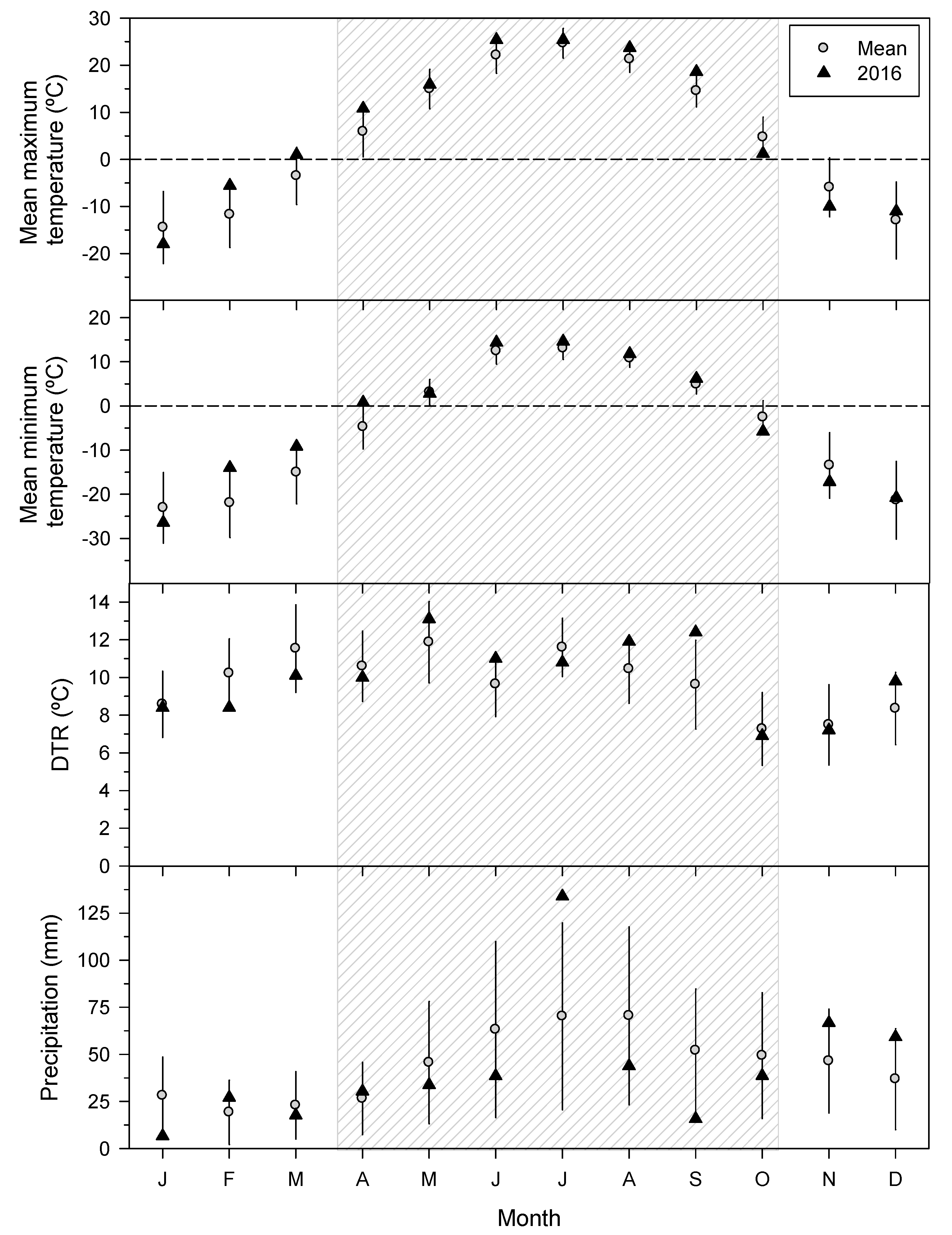
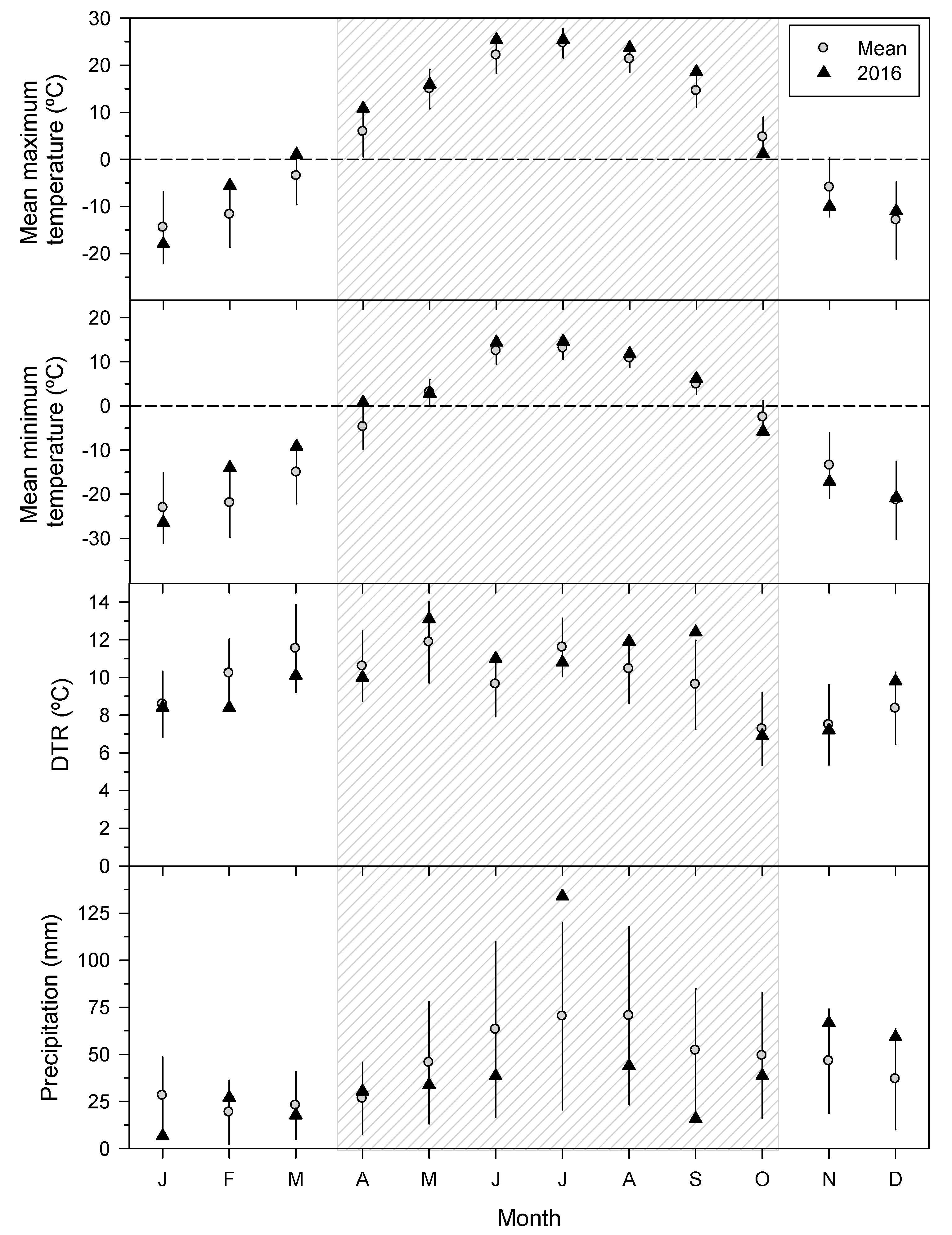
3.1. Climatic Conditions Prior to the SSM Outbreak
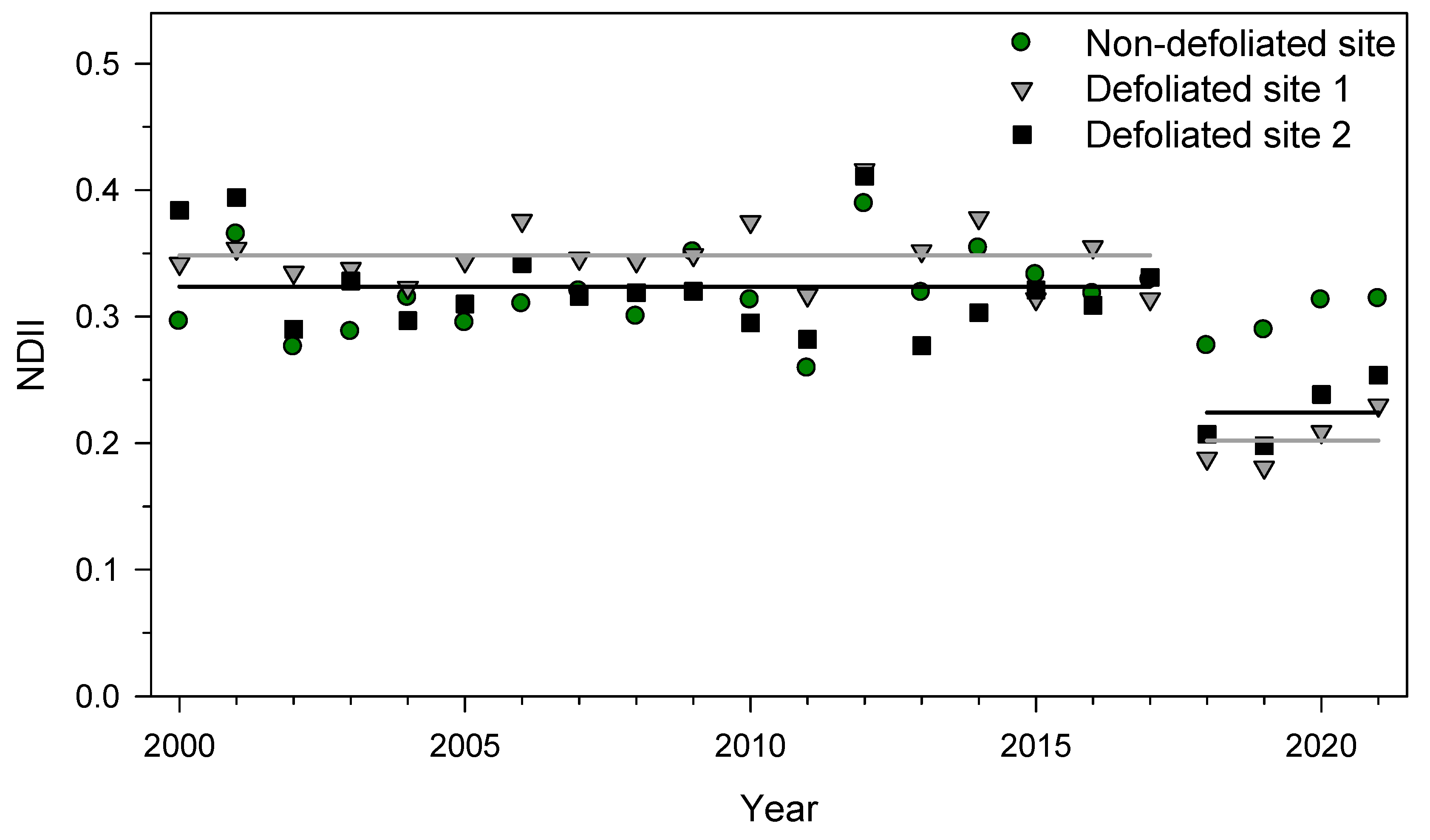
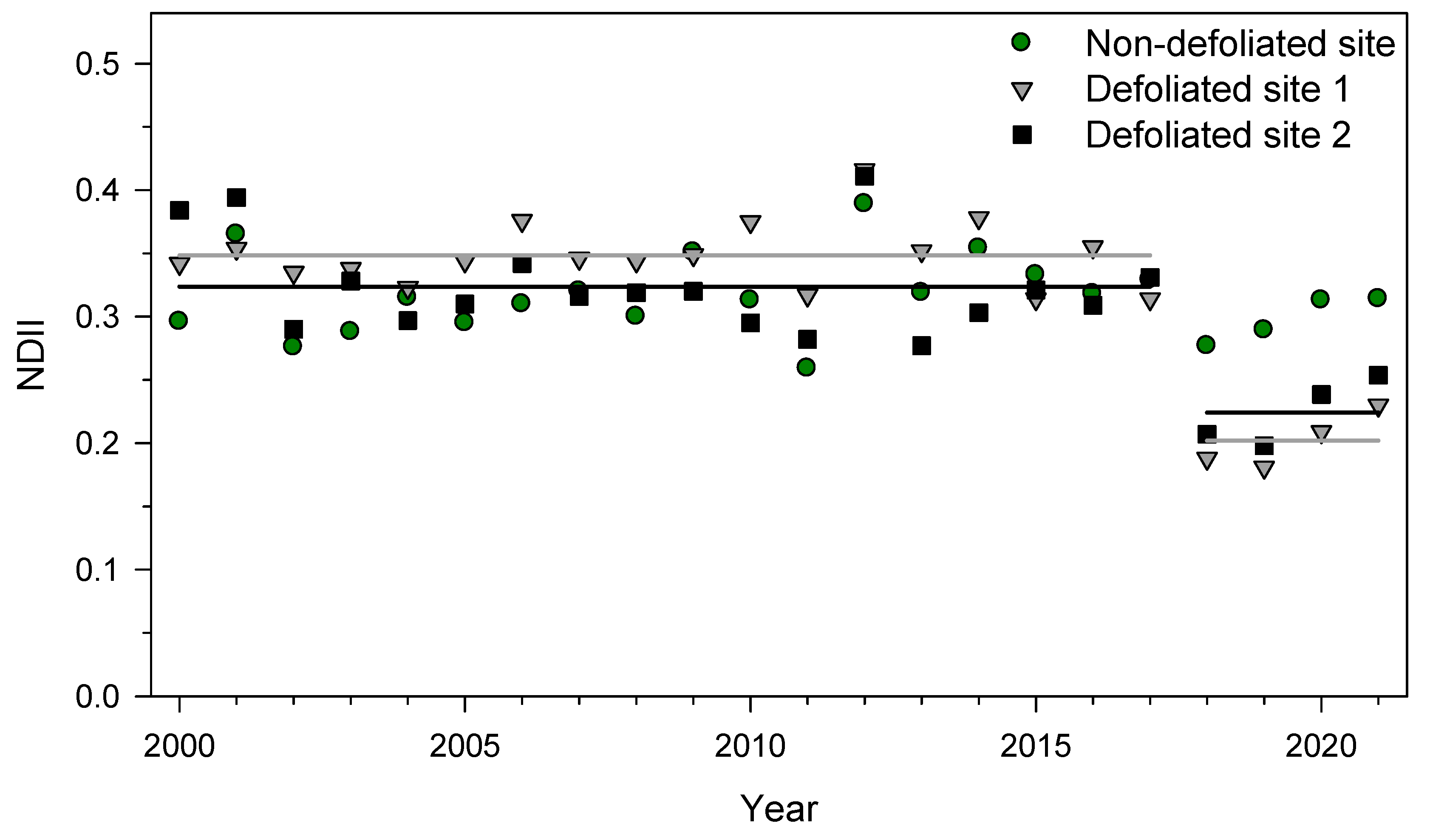
3.2. NDII Changes Due to SSM Defoliation and Drought
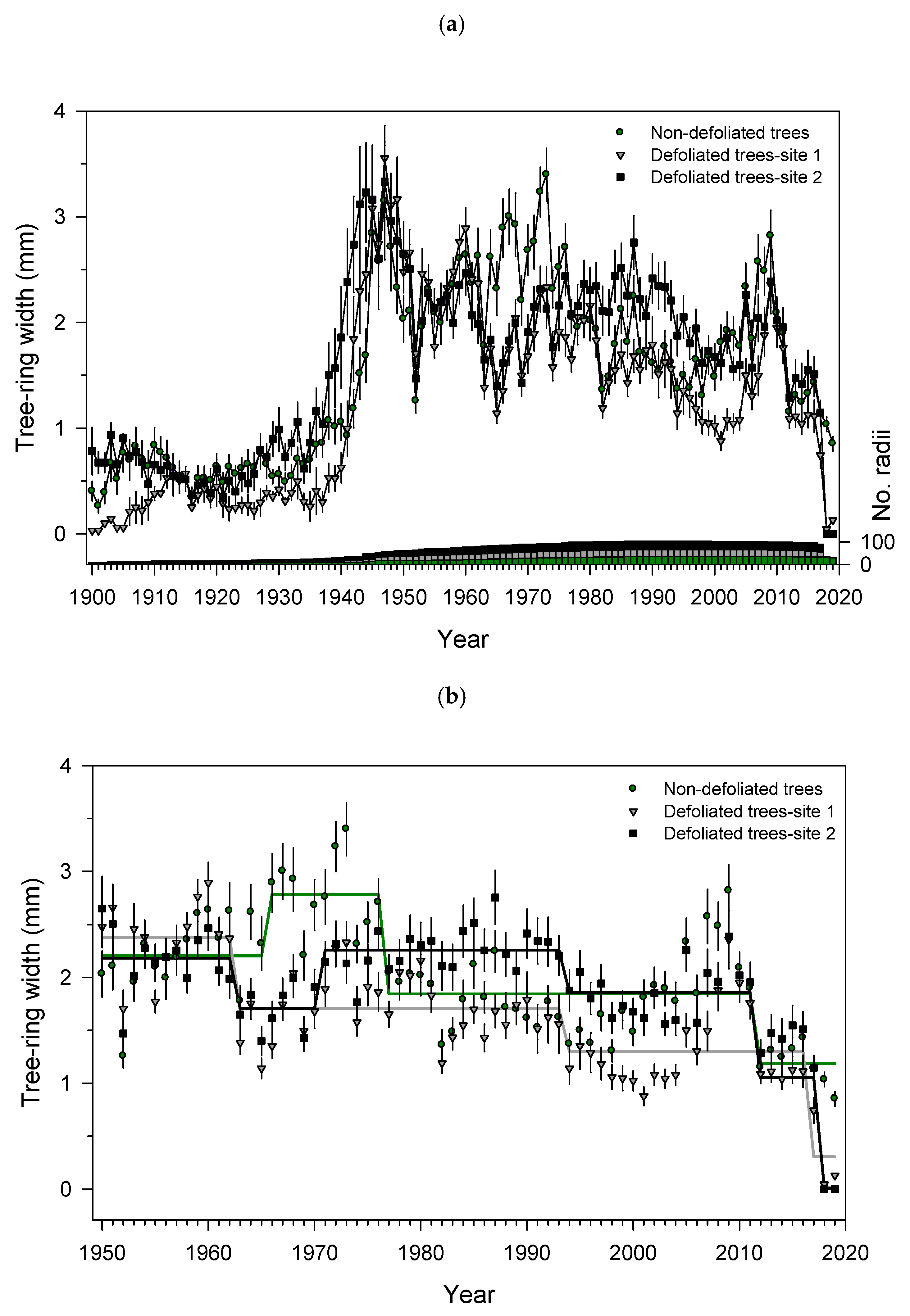
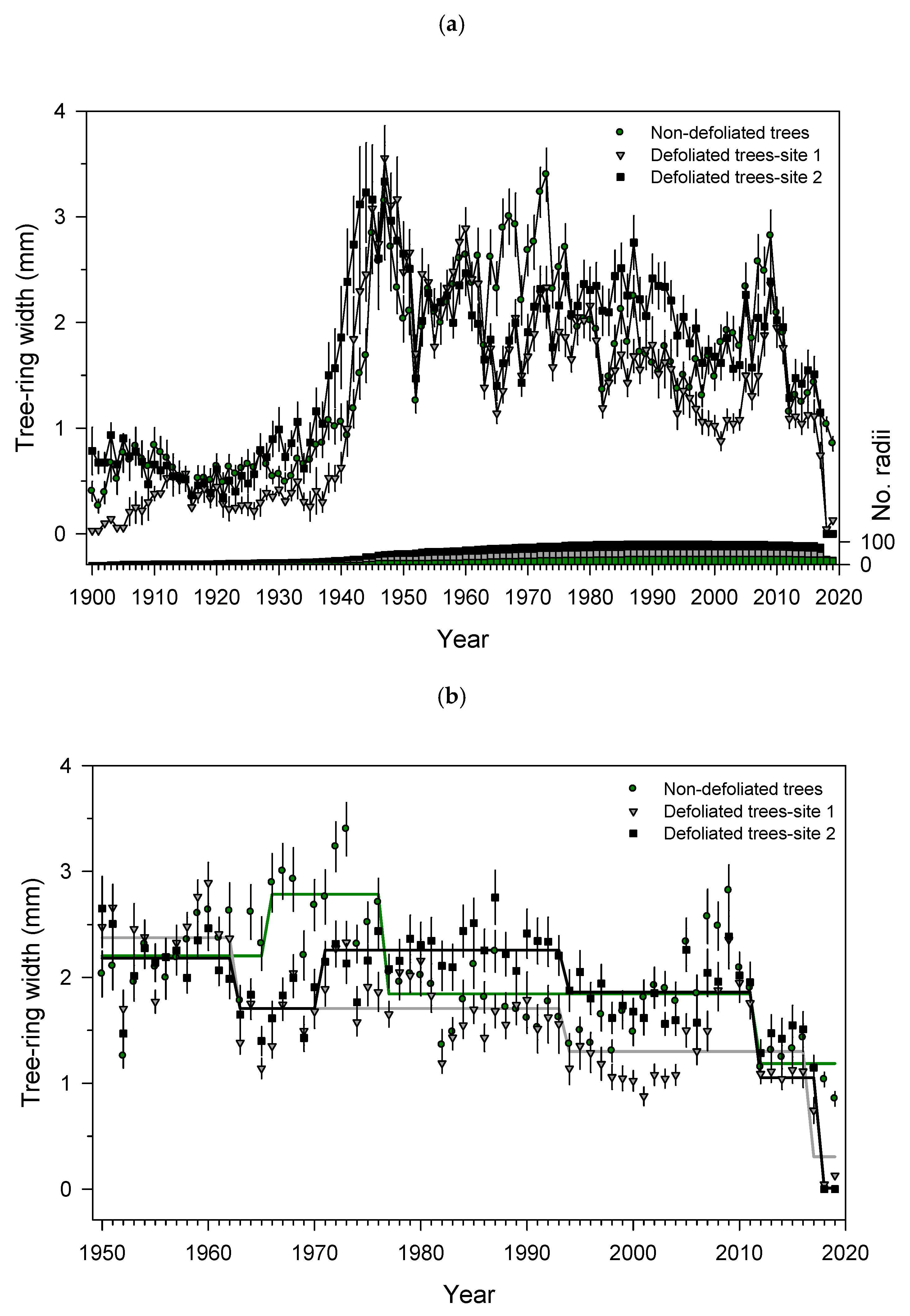
3.3. Growth Patterns
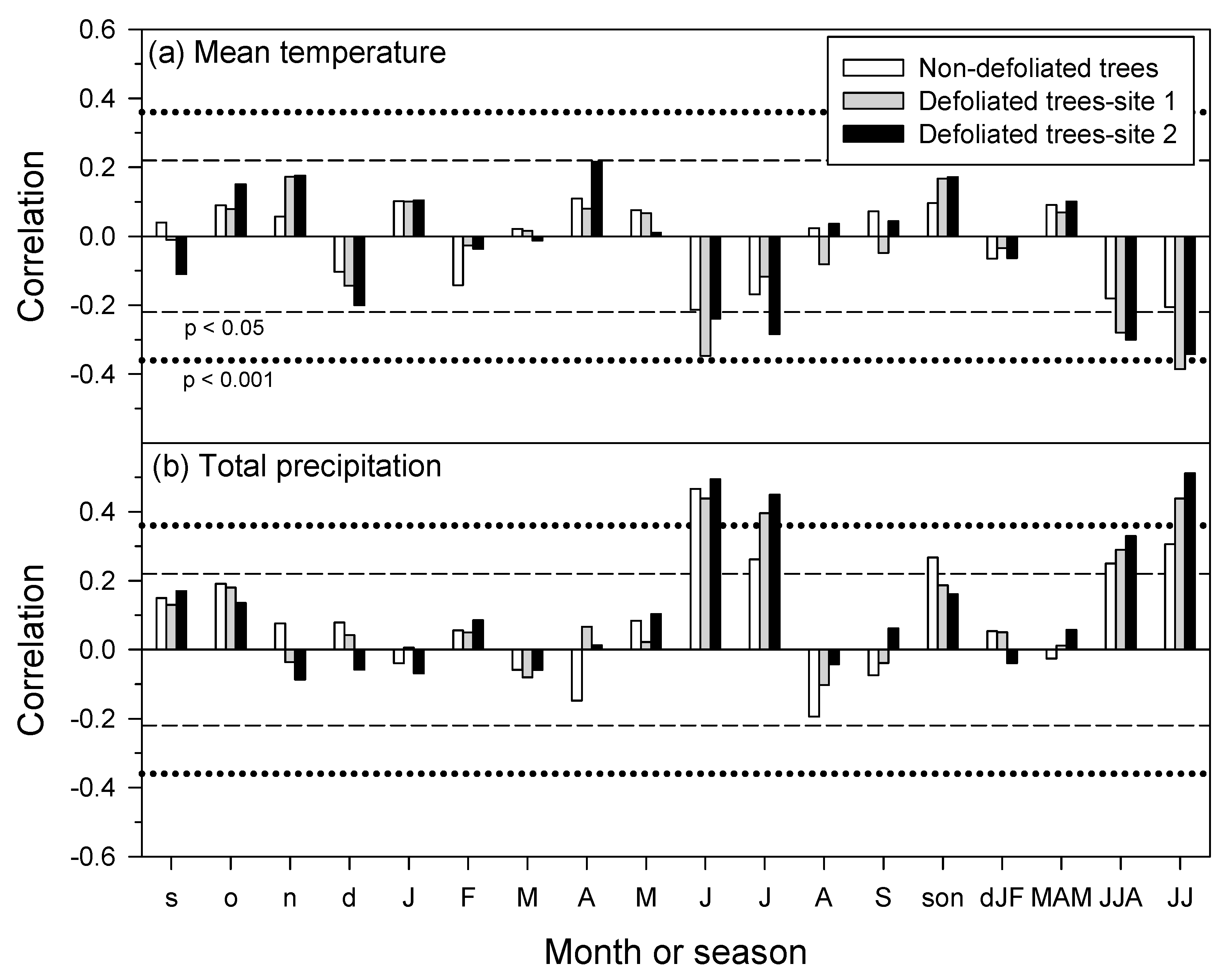
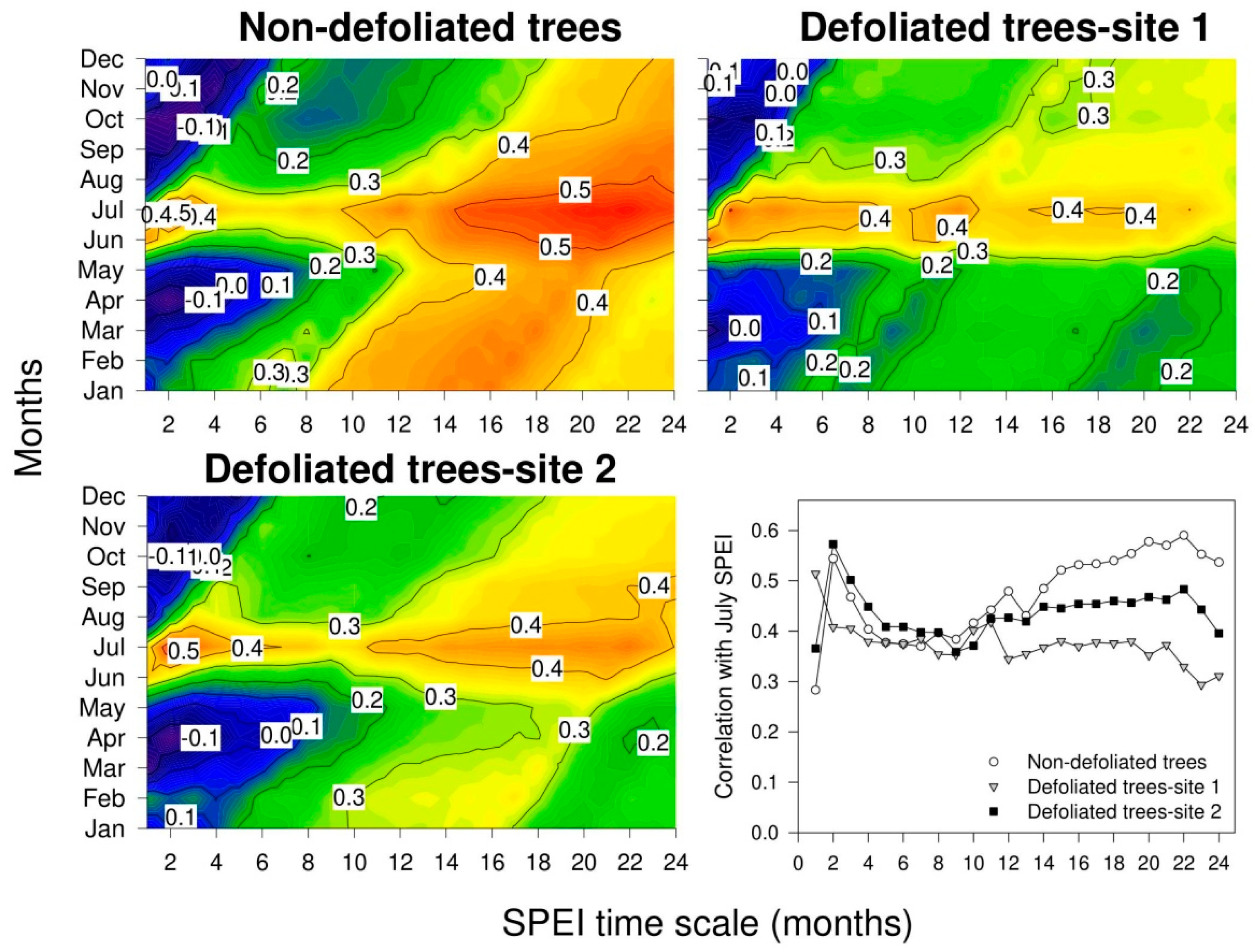
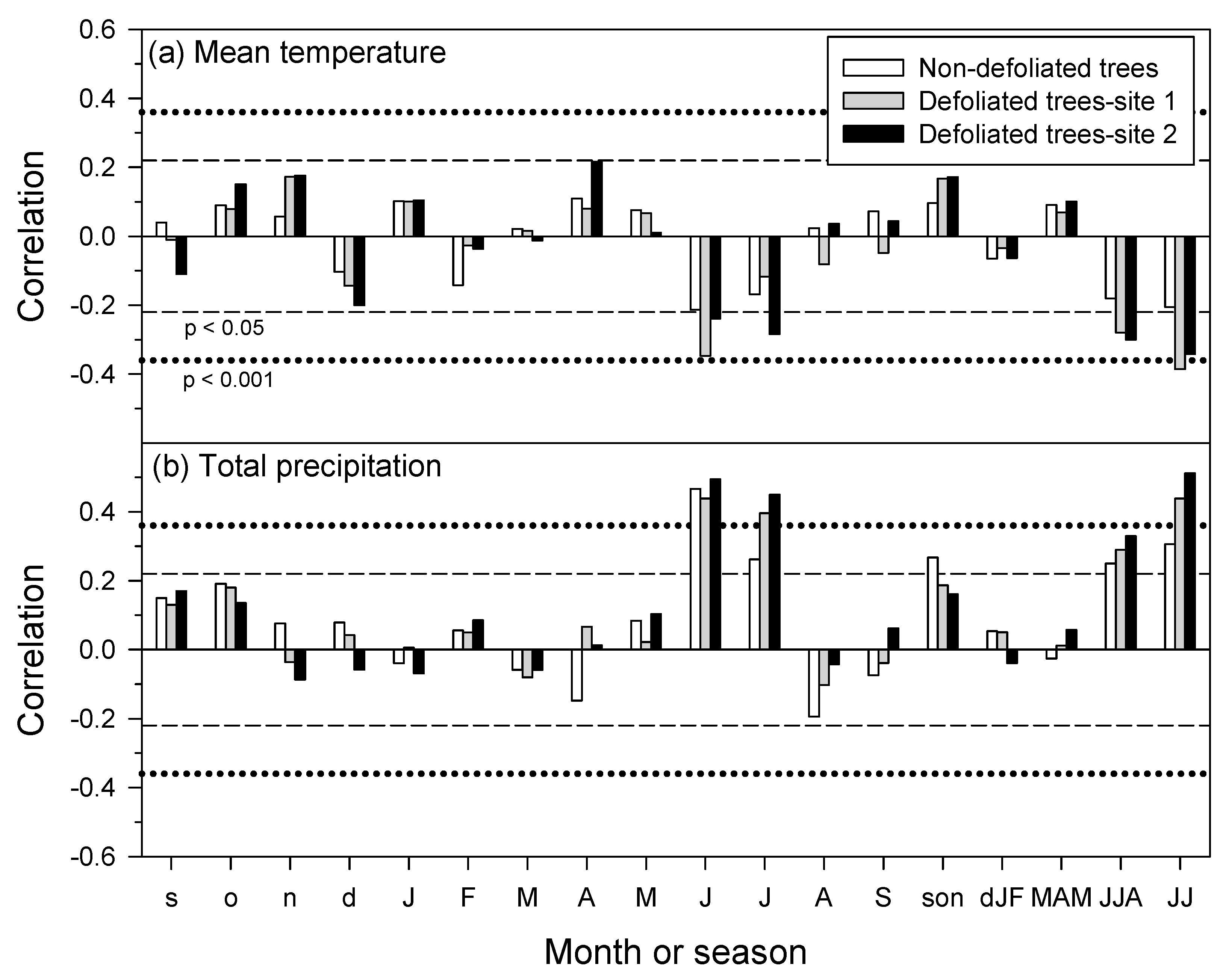
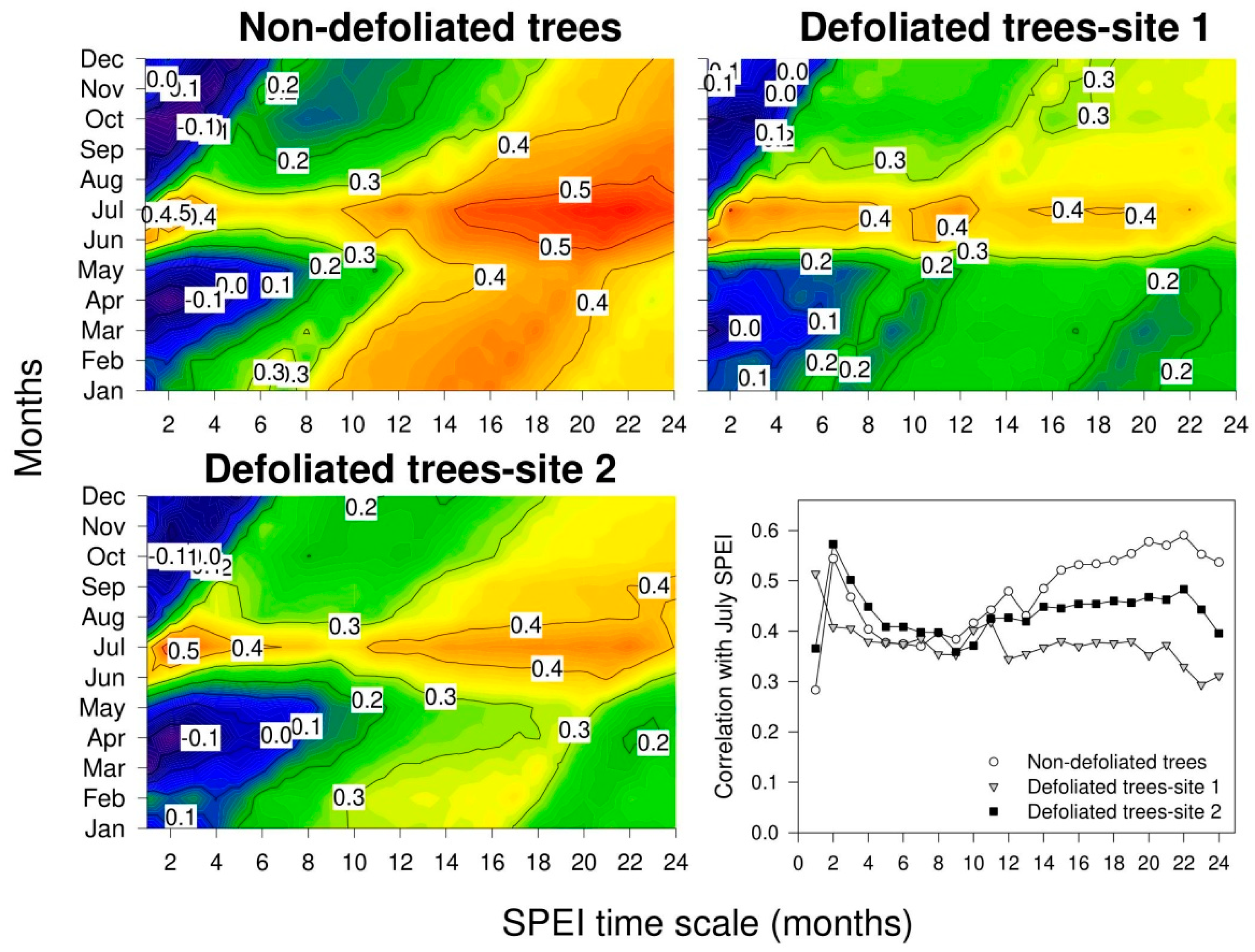
3.4. Growth Responses to Climate Variability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Rojkov, A.S. Siberian Silk Moth Outbreak and Pest Control; Nauka: Moskow, Russia, 1965. (In Russian) [Google Scholar]
- Pureswaran, D.; Grandpré, L.; Paré, D.; Taylor, A.; Barrette, M.; Morin, H.; Régnière, J.; Kneeshaw, D.D. Climate–induced changes in host tree-insect phenology may drive ecological state-shift in boreal forests. Ecology 2015, 96, 1480–1491. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Demidko, D.A.; Fedotova, E.V.; Dvinskaya, M.L.; Budnik, U.A. Spatial and temporal dynamics of Siberian silk moth large-scale outbreak in dark-needle coniferous tree stands in Altai. Contemp. Probl. Ecol. 2016, 9, 711–720. [Google Scholar] [CrossRef]
- Kolb, T.E.; Fettig, C.J.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and anticipated impacts of drought on forests insects and diseases in the United States. For. Ecol. Manag. 2016, 380, 321–334. [Google Scholar] [CrossRef]
- Kondakov, Y.P. Siberian silk moth outbreaks in Krasnoyarskii krai. In Entomology Researches in Siberia; KF REO: Krasnoyarsk, Russia, 2002; pp. 25–74. (In Russian) [Google Scholar]
- Kharuk, V.I.; Im, S.T.; Yagunov, M.N. Migration of the northern boundary of the Siberian silk moth. Contemp. Probl. Ecol. 2018, 11, 26–34. [Google Scholar] [CrossRef]
- Kondakov, Y.P. Patterns of Siberian silk moth outbreaks. In Ecology of Forest Animal Populations in Siberia; Nauka: Novosibirsk, Russia, 1974; pp. 206–264. (In Russian) [Google Scholar]
- Kharuk, V.I.; Ranson, K.J.; Kuz’michev, V.V. Landsat-based analysis of insect outbreaks in southern Siberia. Can. J. Rem. Sens. 2003, 29, 286–297. [Google Scholar] [CrossRef]
- EPPO. Data sheets on quarantine pests: Dendrolimus sibiricus and Dendrolimus superans. EPPO Bull. 2005, 35, 390–395. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.M.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Field, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Kharuk, V.I.; Im, S.T.; Soldatov, V.V. Siberian silkmoth outbreaks surpassed geoclimatic barrier in Siberian Mountains. J. Mt. Sci. 2020, 17, 8. [Google Scholar] [CrossRef]
- Serreze, M.C.; Walsh, J.E.I.; Chapin, F.S.; Osterkamp, T.; Dyurgerov, M.; Romanovsky, V.; Oechel, W.C.; Morison, J.; Zhang, T.; Barry, R.G. Observational evidence of recent change in the northern high-latitude environment. Clim. Chang. 2000, 46, 159–207. [Google Scholar] [CrossRef]
- Groisman, P.Y.; Blyakharchuk, T.A.; Chernokulsky, A.V.; Arzhanov, M.M.; Marchesini, L.B.; Bogdanova, G.; Borzenkova, I.I.; Bulygina, O.N.; Karpenko, A.A. Climate changes in Siberia. In Regional Environmental Changes in SIBERIA and Their Global Consequences; Groisman, P., Gutman, G., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 57–109. [Google Scholar]
- Baranchikov, Y.N.; Montgomery, M.E. Siberian Moth: (Dendrolimus sibiricus [Chetverikov]) (Lepidoptera: Lasiocampidae). In The Use of Classical Biological Control to Preserve Forests in North America; United States Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2014; pp. 383–391. [Google Scholar]
- Camarero, J.J.; Tardif, T.; Gazol, A.; Conciatori, F. Pine processionary moth outbreaks cause longer growth legacies than drought and are linked to the North Atlantic Oscillation. Sci. Total Environ. 2022, 819, 153041. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.; Cartensen, J.; Hernández-García, E.; Duarte, C.M. Ecological thresholds and regime shifts: Approaches to identification. Trends Ecol. Evol. 2009, 24, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart-Haëntjens, E.J.; Curtis, P.S.; Fahey, R.T.; Vogel, C.S.; Gough, C.M. Net primary production of a temperate deciduous forest exhibits a threshold response to increasing disturbance severity. Ecology 2015, 96, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Shushpanov, A.S.; Petrov, I.A.; Demidko, D.A.; Im, S.T.; Knorre, A.A. Fir (Abies sibirica Ledeb.) mortality in mountain forests of the eastern Sayan Ridge, Siberia. Contemp. Probl. Ecol. 2019, 12, 299–309. [Google Scholar] [CrossRef]
- Bréda, N.; Badeau, V. Forest tree responses to extreme drought and some biotic events: Towards a selection according to hazard tolerance? C. R. Geosci. 2008, 340, 651–662. [Google Scholar] [CrossRef]
- Rodionov, S.N. A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 2004, 31, 5. [Google Scholar] [CrossRef] [Green Version]
- González de Andrés, E.; Shestakova, T.A.; Scholten, R.C.; Delcourt, C.J.F.; Gorina, N.V.; Camarero, J.J. Changes in tree growth synchrony and resilience in Siberian Pinus sylvestris forests are modulated by fire dynamics and ecohydrological conditions. Agric. For. Meteorol. 2022, 312, 108712. [Google Scholar] [CrossRef]
- Menne, M.J.; Williams, C.N.; Gleason, B.E.; Jared Rennie, J.; Lawrimore, J.H. The global historical climatology network monthly temperature dataset, version 4. J. Clim. 2018, 31, 9835–9854. [Google Scholar] [CrossRef]
- Kovalev, A.; Soukhovolsky, V. Analysis of forest stand resistance to insect attack according to remote sensing data. Forests 2021, 12, 1188. [Google Scholar] [CrossRef]
- Kirichenko, N.I.; Baranchikov, Y.N.; Vidal, S. Performance of the potentially invasive Siberian moth Dendrolimus superans sibiricus on coniferous species in Europe. Agric. For. Entomol. 2009, 11, 247–254. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Tchebakova, N.; Kirichenko, N.; Parphenova, E.; Korezts, M.; Kenis, M. Biological invasions: The Siberian moth, Dendrolimus superans sibiricus, a potential invader in Europe? In Atlas of Biodiversity Risk; Settele, J., Penev, L., Georgiev, T., Grabaum, R., Grobelnik, V., Hammen, V., Klotz, S., Kotarac, M., Kuehn, I., Eds.; Pensoft Publ.: Sofia, Bulgaria, 2010. [Google Scholar]
- Flø, D.; Rafoss, T.; Wendell, M.; Sundheim, L. The Siberian moth (Dendrolimus sibiricus), a pest risk assessment for Norway. For. Ecosyst. 2020, 7, 48. [Google Scholar] [CrossRef]
- Sultson, S.M.; Goroshko, A.A.; Verkhovets, S.V.; Mikhaylov, P.V.; Ivanov, V.A.; Demidko, D.A.; Kulakov, S.S. Orographic factors as a predictor of the spread of the Siberian silk moth outbreak in the mountainous southern taiga Forests of Siberia. Land 2021, 10, 115. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Antamoshkina, O.A. Impact of silkmoth outbreak on taiga wildfires. Contemp. Probl. Ecol. 2017, 10, 556–562. [Google Scholar] [CrossRef]
- Dudin, V.A. Use and restoration of silkmoth-damaged forests in Tomsk oblast. Tr. Lesn. Khoz. Sib. 1958, 4, 262–268. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [Green Version]
- Fritts, H.C. Tree-Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Duncan, R.P. An evaluation of errors in tree age estimates based on increment cores in kahikatea (Dacrycarpus dacrydioides). N. Z. Nat. Sci. 1989, 16, 31–37. [Google Scholar]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program Library in R; R Package Version 1.7.1; dplR: Milperra, Australia, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 11 January 2022).
- Briffa, K.R.; Jones, P.D. Basic chronology statistics and assessment. In Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Acad. Publ.: Dordrecht, The Netherlands, 1990; pp. 137–152. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Biondi, F. Treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Hantson, S.; Chuvieco, E. Evaluation of different topographic correction methods for Landsat imagery. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 691–700. [Google Scholar] [CrossRef]
- Roy, D.P.; Kovalskyy, V.; Zhang, H.K.; Vermote, E.F.; Yan, L.; Kumar, S.S.; Egorov, A. Characterization of Landsat-7 to Landsat-8 reflective wavelength and normalized difference vegetation index continuity. Remote Sens. Environ. 2016, 185, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hardisky, M.; Klemas, V.; Smart, R. The Influences of soil salinity, growth form, and leaf moisture on the spectral reflectance of Spartina alterniflora canopies. Photogramm. Eng. Remote Sens. 1983, 49, 77–83. [Google Scholar]
- Rodionov, S.N. A brief overview of the regime shift detection methods. Large-scale disturbances (regime shifts) and recovery in aquatic ecosystems: Challenges for management toward sustainability. In Proceedings of the UNESCO-ROSTE/BAS Workshop on Regime Shifts, Varna, Bulgaria, 14–16 June 2005; pp. 17–24. [Google Scholar]
- Rodionov, S.N. Use of prewhitening in climate regime shift detection. Geophys. Res. Lett. 2006, 33, 245. [Google Scholar] [CrossRef] [Green Version]
- Stirnimann, L.; Conversi, A.; Marini, S. Detection of regime shifts in the environment: Testing “STARS” using synthetic and observed time series. ICES J. Mar. Sci 2019, 76, 2286–2296. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Bunn, A.G. Responses of the circumpolar boreal forest to 20th century climate variability. Environ. Res. Lett. 2007, 2, 045013. [Google Scholar] [CrossRef]
- Kagawa, A.; Naito, D.; Sugimoto, A.; Maximov, T.C. Effects of spatial and temporal variability in soil moisture on widths and δ13C values of eastern Siberian tree rings. J. Geophys. Res. 2003, 108, 4500. [Google Scholar] [CrossRef]
- Belokopytova, L.V.; Babushkina, E.A.; Zhirnova, D.F. Climatic response of conifer radial growth in forest-steppes of South Siberia: Comparison of three approaches. Contemp. Probl. Ecol. 2018, 11, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Shestakova, T.A.; Gutiérrez, E.; Valeriano, C.; Lapshina, E.; Voltas, J. Recent loss of sensitivity to summer temperature constrains tree growth synchrony among boreal Eurasian forests. Agric. For. Meteorol. 2019, 268, 318–330. [Google Scholar] [CrossRef]
- Tabakova, M.A.; Arzac, A.; Martínez, E.; Kirdyanov, A.V. Climatic factors controlling Pinus sylvestris radial growth along a transect of increasing continentality in southern Siberia. Dendrochronologia 2020, 62, 125709. [Google Scholar] [CrossRef]
- Kirichenko, N.; Flament, J.; Baranchikov, Y.; Gregoire, J.C. Larval performances and life cycle completion of the Siberian moth, Dendrolimus sibiricus (Lepidoptera: Lasiocampidae), on potential host plants in Europe: A laboratory study on potted trees. Eur. J. For. Res. 2011, 130, 1067–1074. [Google Scholar] [CrossRef]
- Florov, D.N. Pests of Siberian Forests; OGIX: Irkutsk, Russia, 1948; p. 143. [Google Scholar]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (Code) | Latitude N | Longitude E | Elevation (m a.s.l.) |
|---|---|---|---|
| Defoliated 1 (D1) | 57°09′01″ | 84°43′01″ | 137 |
| Defoliated 2 (D2) | 57°08′12″ | 84°42′01″ | 102 |
| Not defoliated (ND) | 57°08′33″ | 84°42′25″ | 197 |
| Site | DBH (cm) | No. Trees (No. Cores) | Age at 1.3 m (Years) | Tree-Ring Width (mm) | Timespan | AR1 | MSx | rbar |
|---|---|---|---|---|---|---|---|---|
| D1 | 29.6 ± 10.7 | 17 (34) | 72 ± 22 | 1.64 ± 1.05 | 1865–2019 | 0.46 ± 0.13 | 0.29 ± 0.04 | 0.41 ± 0.12 |
| D2 | 36.3 ± 11.0 | 18 (36) | 79 ± 24 | 1.93 ± 1.25 | 1876–2019 | 0.37 ± 0.17 | 0.27 ± 0.03 | 0.44 ± 0.14 |
| ND | 36.7 ± 8.6 | 17 (34) | 77 ± 33 | 1.76 ± 1.17 | 1876–2019 | 0.39 ± 0.16 | 0.27 ± 0.04 | 0.39 ± 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camarero, J.J.; Shestakova, T.A.; Pizarro, M. Threshold Responses of Canopy Cover and Tree Growth to Drought and Siberian silk Moth Outbreak in Southern Taiga Picea obovata Forests. Forests 2022, 13, 768. https://doi.org/10.3390/f13050768
Camarero JJ, Shestakova TA, Pizarro M. Threshold Responses of Canopy Cover and Tree Growth to Drought and Siberian silk Moth Outbreak in Southern Taiga Picea obovata Forests. Forests. 2022; 13(5):768. https://doi.org/10.3390/f13050768
Chicago/Turabian StyleCamarero, Jesús Julio, Tatiana A. Shestakova, and Manuel Pizarro. 2022. "Threshold Responses of Canopy Cover and Tree Growth to Drought and Siberian silk Moth Outbreak in Southern Taiga Picea obovata Forests" Forests 13, no. 5: 768. https://doi.org/10.3390/f13050768
APA StyleCamarero, J. J., Shestakova, T. A., & Pizarro, M. (2022). Threshold Responses of Canopy Cover and Tree Growth to Drought and Siberian silk Moth Outbreak in Southern Taiga Picea obovata Forests. Forests, 13(5), 768. https://doi.org/10.3390/f13050768