1. Introduction
Magnolia (Fam. Magnoliaceae) is a plant genus that consists of more than 300 species [
1,
2,
3,
4,
5]. This genus has a wide distribution in subtropical and tropical Asia and America [
2,
4,
5]. The Magnolia genus includes evergreen and deciduous trees and shrubs [
2,
4], and many species are prominently used as ornamental plants, timber, medicinal raw materials, cosmetics, and essential oils [
2,
3,
6,
7,
8,
9,
10]. Despite Magnolia’s crucial uses, the assessment in [
2] using the International Union for Conservation of Nature (IUCN) criteria resulted in 147 species of
Magnolia being categorized as threatened (Critically Endangered, Endangered, and Vulnerable) due to various threats such as continued deforestation, habitat destruction, and over-harvesting.
In Indonesia, there are 28 species of
Magnolia distributed in Sumatra, Java, Lesser Sunda Islands, Sulawesi, Moluccas, and Papua [
2,
7,
10]. Among these species, one species was categorized as Threatened, i.e.,
Magnolia sulawesiana Brambach, Noot., and Culmsee (Endangered); one as Near Threatened (
M. borneensis); five under Least Concern; and the rest under Data Deficiency. Indonesia is one of the countries with the least amount of information on
Magnolia, especially for the threatened taxa [
11].
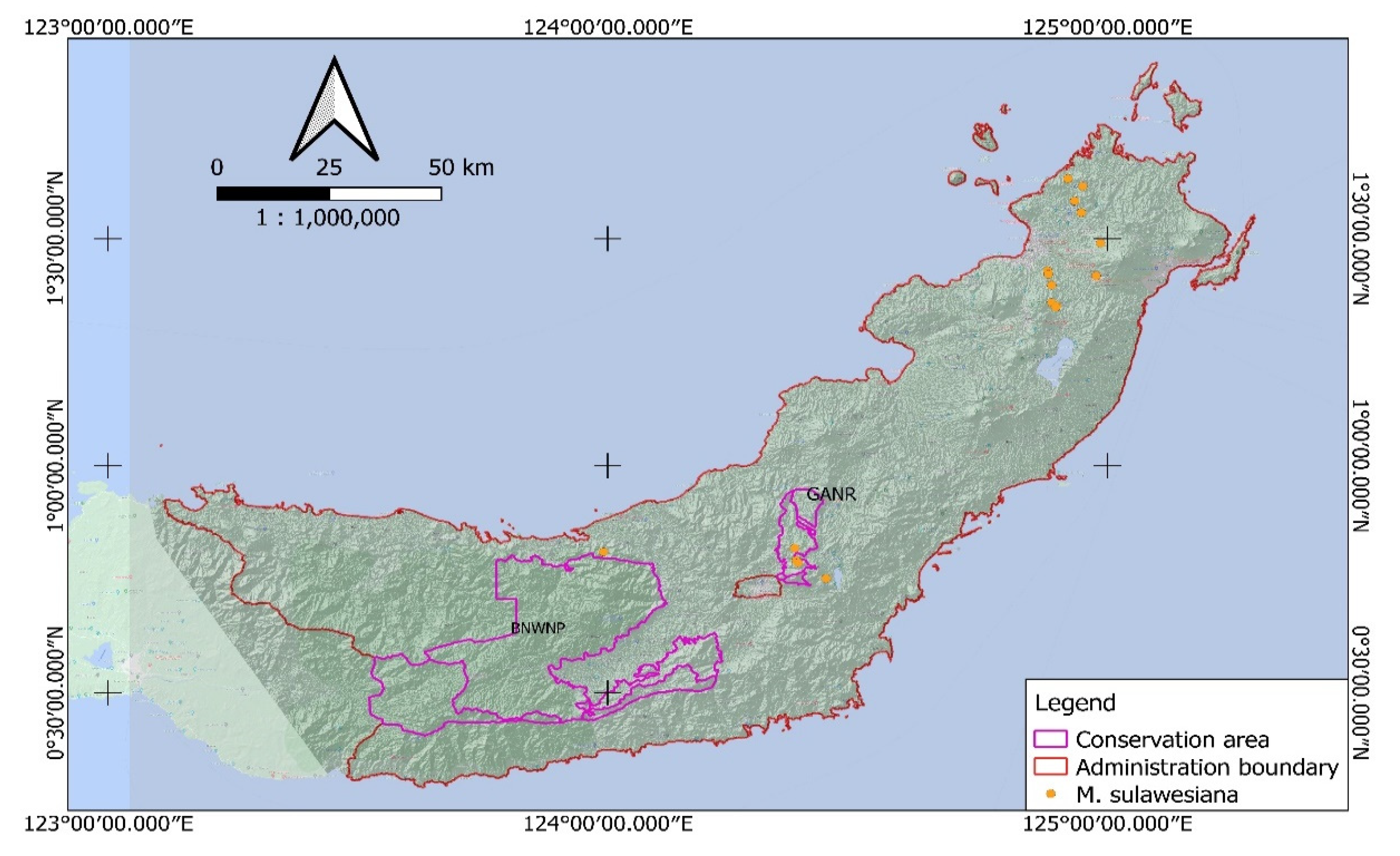
The endangered
M. sulawesiana is an endemic species that grows naturally only in three locations within the mountain range in the central part of Sulawesi [
12]. Considering the increasing rate of forest-cover loss in Sulawesi, which was 10.98% between 2000 and 2007 [
13], as well as the species’ current red-list status as Endangered [
2], it is crucial to find this endemic species in other areas of Sulawesi. The central parts of Sulawesi, including the Central and West Sulawesi Province, where
M. sulawesiana was found, face deforestation rates of 0.68 and 0.84%, respectively [
13]. Deforestation is not the only threat to
Magnolia; overharvesting, poor fruiting, and low natural regeneration [
11] also add pressure to the endemic species’ vulnerability in the wild.
In Indonesia, overharvesting might become a real threat since Magnolia species are commercially traded. This is especially true for
M. sulawesiana because it is challenging to distinguish
M. sulawesiana wood from other Magnolia woods on the market. Magnolia species have a long historical connection with the Minahasa tribe, one of the tribes in the northern part of Sulawesi. The
Magnolia woods are known as
Cempaka or
Wasian and were used as material to construct Woloan, a traditional Minahasa wooden house [
14,
15,
16]. In the 1970s, when forest concession rightsholder companies began to operate in the production forest,
Magnolia wood became prominent because of its good quality. In response to the high demand for
Magnolia wood, local communities started to plant
Magnolia species [
17,
18]. Today, the Minahasa district is known to have the largest community plantation forest containing
Magnolia species among the areas in Sulawesi [
15,
19,
20].
There is also
Magnolia cubensis in Cuba, which is a highly endangered and endemic species that requires conservation measures. The need for conservation action was determined based on the findings of studies on the influence of habitat fragmentation on the species’ population structure and genetic diversity [
21]. In Western Mexico,
Magnolia granbarrancae,
M. pugana,
M. talpana, and
M. vallartensis are also critically endangered species because their extent of occurrence (EOO) and area of occupancy (AOO) are below the limits set by the IUCN, and they also have a low genetic diversity [
22].
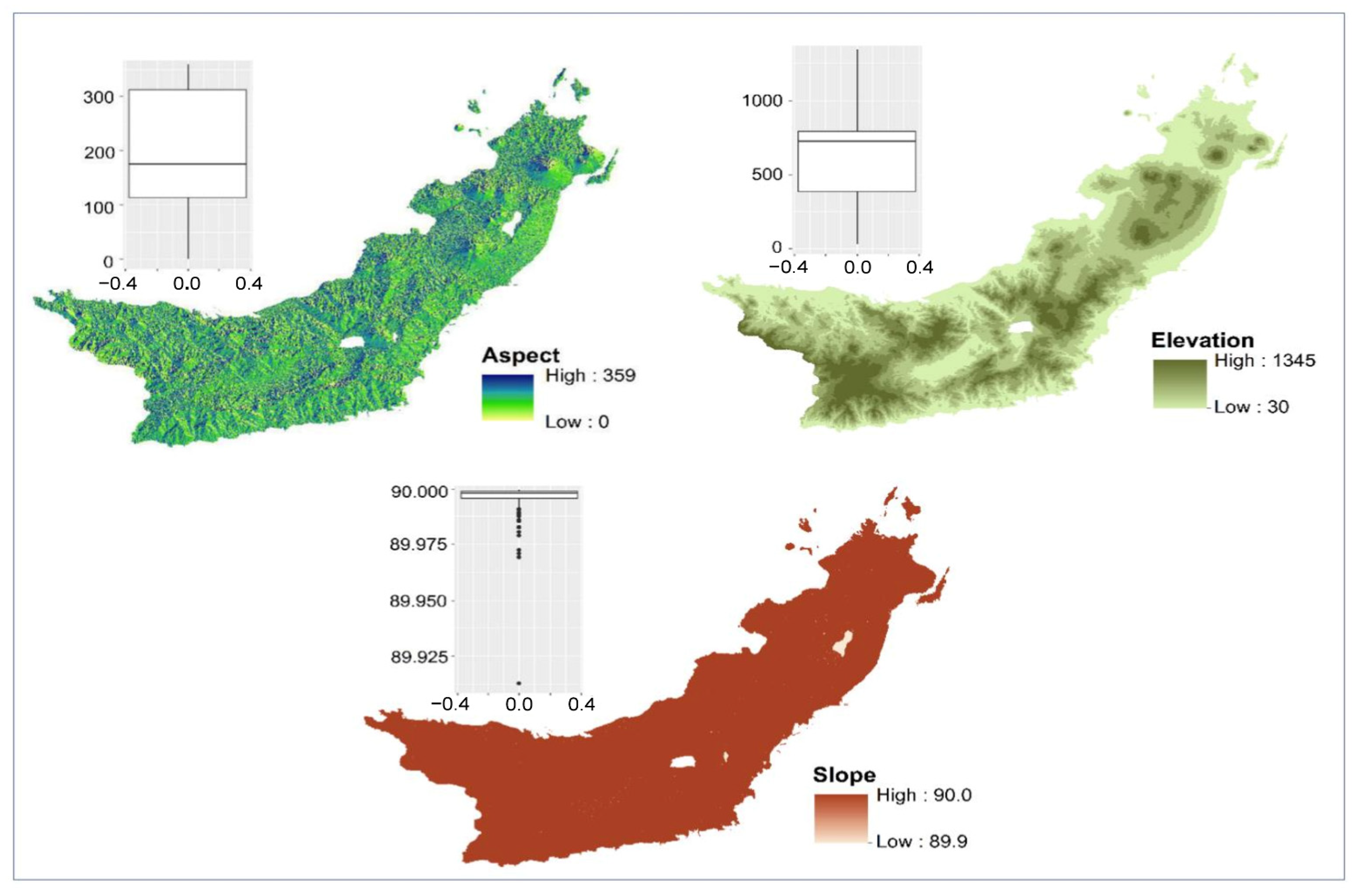
The spatial distribution of plant species is not the result of a random event but is influenced by environmental variables, especially climate and topography characteristics [
23,
24], as well as soil, temperature, hydrology, and spatial constraints, which affect plant distribution [
25,
26,
27,
28,
29]. Information on the distribution of
Magnolia spp. in North Sulawesi is crucial for conservation strategies and landscape protection. Understanding the habitat preferences and suitability of
Magnolia spp. is also important to determine the species’ functions in its surrounding community, including the associated animals [
30]. An assessment conducted in China using climate and terrain variables demonstrated differing habitat preferences among
Magnolia species [
8]. Despite its importance for conservation strategies, this type of assessment has never been used for
Magnolia species in Indonesia.
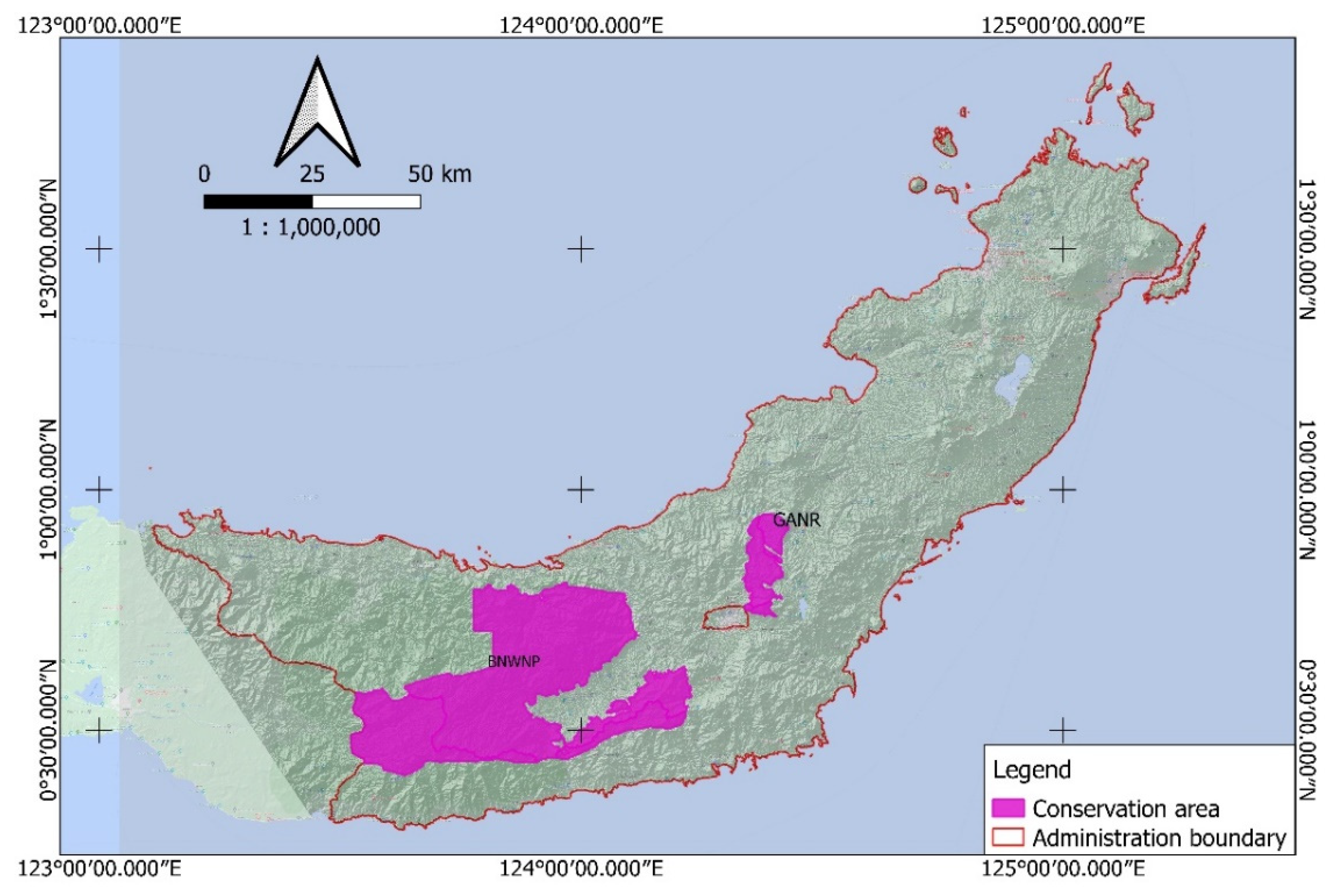
This study aims to identify the distribution of
Magnolia species in the northern part of Sulawesi, including the endemic
M. sulawesiana. We distinguished the habitat preferences of
Magnolia and combined them with spatial data to estimate the potential species distribution. The discovery of
M. sulawesiana distribution in the northern part of Sulawesi will lead to a new record of this species’ distribution in the Wallacea bioregion [
29], which features high species endemism but is still poorly understood.
4. Discussion
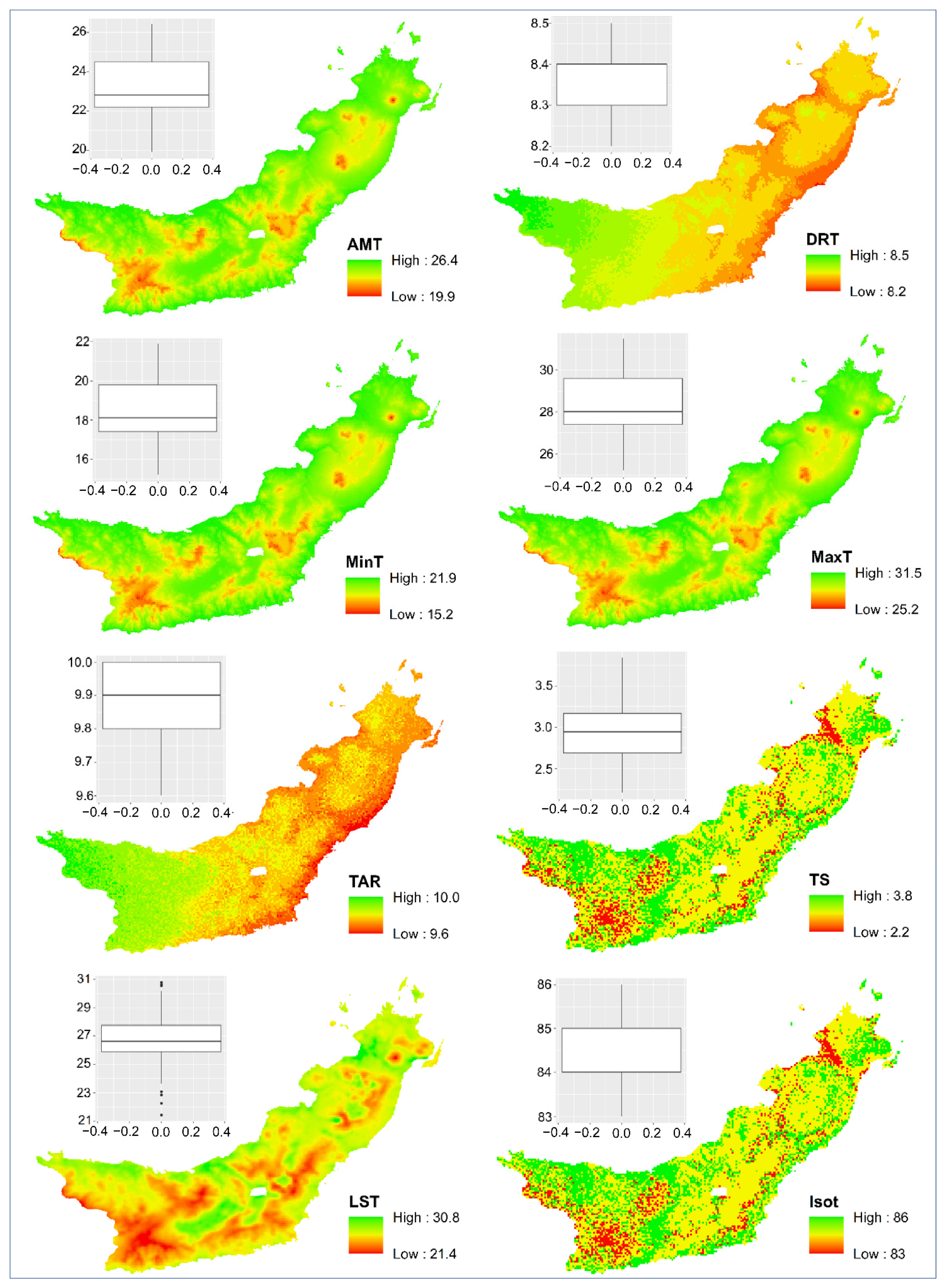
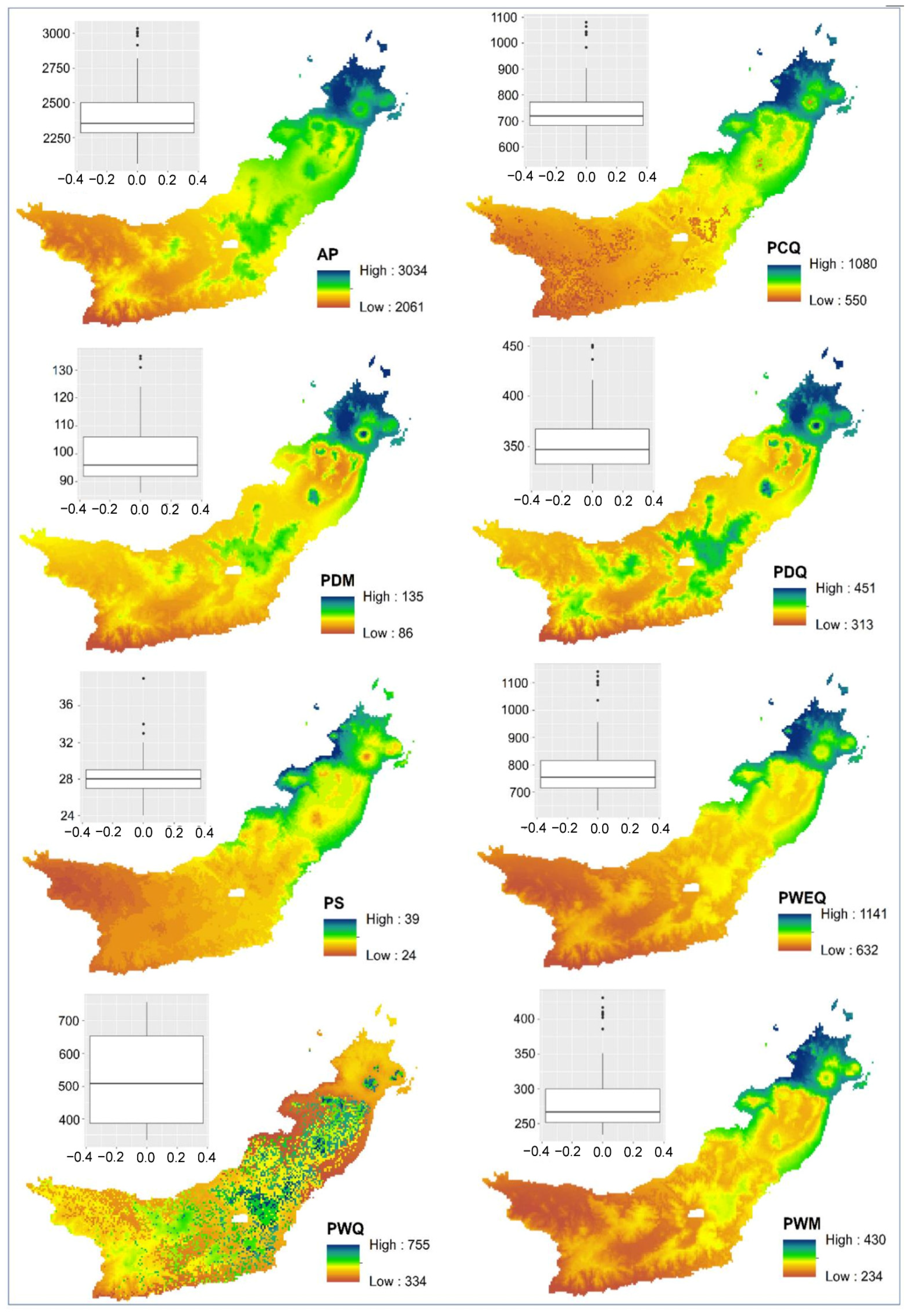
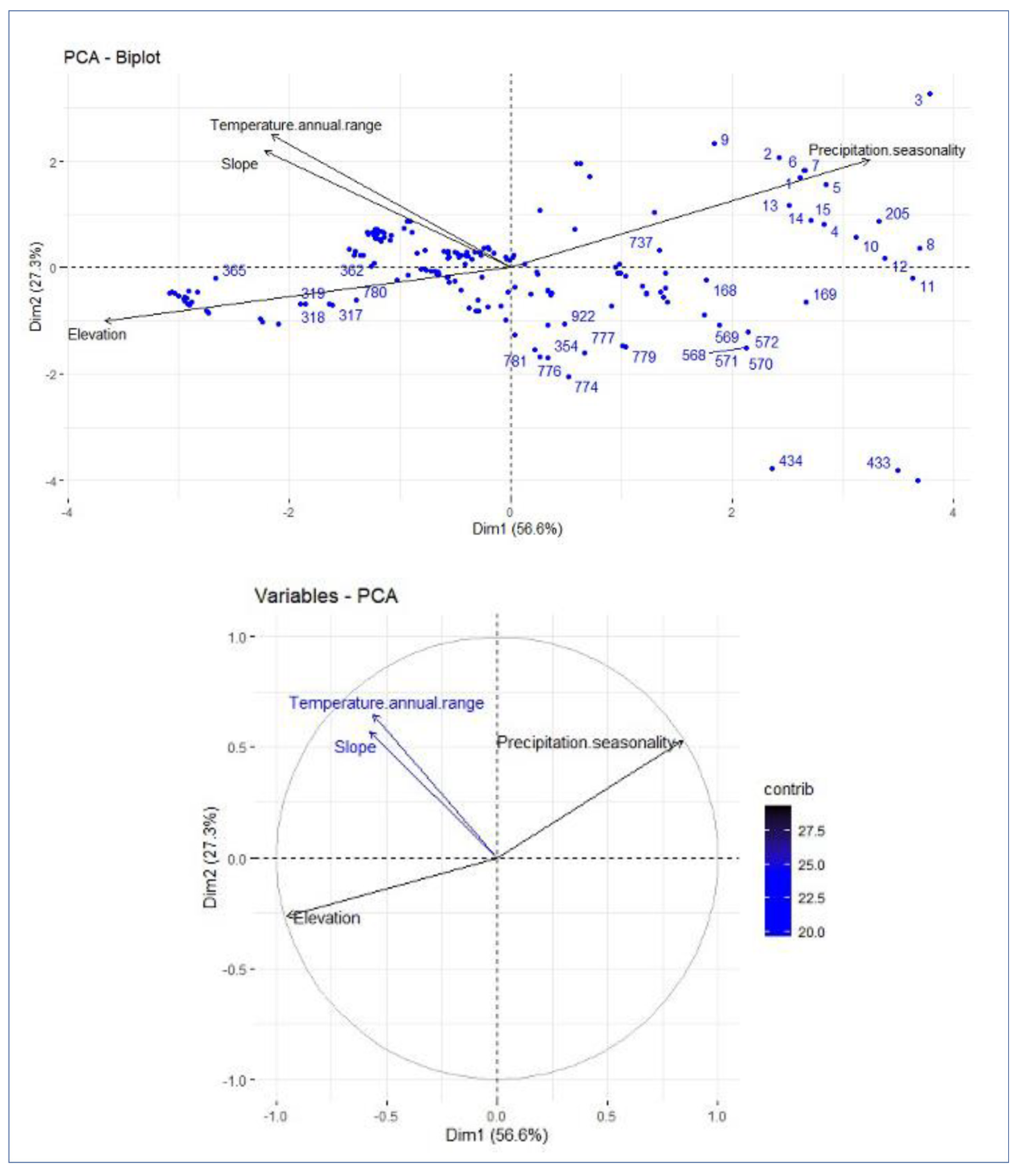
The spatial distribution of
Magnolia in North Sulawesi Province is influenced by the annual temperature range, precipitation seasonality, and elevation. At the landscape scale in this extremely varied environment, topography and climate were reported to be significant determinants of species richness, endemic richness, and endemicity [
23,
24]. The influence of climate on
Magnolia distribution demonstrates the species’ vulnerability to climate changes [
40].
Magnolia is also reported to have allopatric speciation [
3]. Thus, geographical isolation [
3], topography, and climatic factors [
26] have led to the scarcity of this genus due to a lack of specific habitat suitability. On the other hand, changes in climatic parameters such as temperature can also lead to the distribution of a species outside of its native range, as shown in [
41] for
M. grandiflora.
Based on our results, despite the importance of climatic factors, elevation seems to have the largest influence on
Magnolia species distribution in North Sulawesi Province. Elevation can have a 10% to 50% effect on plant distribution [
27,
42]. For
M. sulawesiana, we found that this species has a wider elevation range due to the presence of outlier data. One individual was found at 175 m asl, while the rest of the individuals, including the individual recorded in [
12], were found from 1100 to 2000 m asl. This is a very interesting result because the individual was identified as
M. sulawesiana based on morphological characteristics [
12] and located inside GANP, where it remains unclear if this species grows naturally. A closer study needs to be conducted to explain this phenomenon. There might be other factors that determine species distribution other than climatic and elevation factors. Other research shows that most of the endemic and endangered species of
Magnolia are naturally found in tropical mountain forests and at high elevations. Examples include
M. schiedeana in Mexico [
43],
M. sinica in Yunan (1339–1707 m asl) [
44],
M. vovidesii in Mexico (1520–1550 m asl) [
21], and
M. granbarrancae (1073–1215 m asl) [
22].
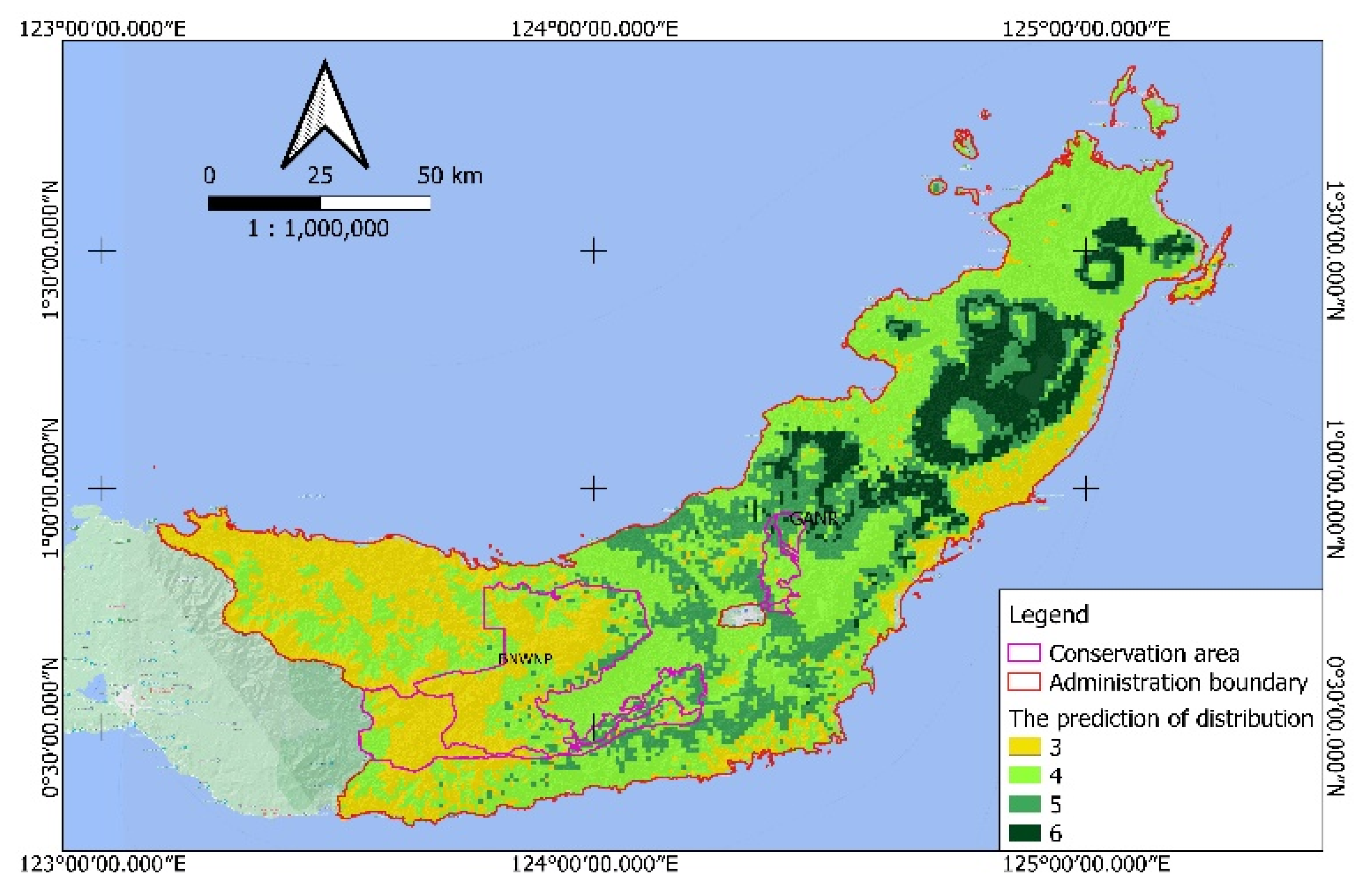
The growing demand for
Magnolia trees in the lumber market throughout the year has led to consequences such as the increasing rarity of this genus. To meet these needs, wood is harvested not only from
Magnolia plantations but also from the species’ natural habitat. This study demonstrated that the distribution of
Magnolia spp. in production forests is decreasing. Without any effort to create a community development program to maintain the balance between different needs,
Magnolia species will gradually become rare and, eventually, extinct. As a result, the existence of this genus is threatened. Scattered, small-scale
Magnolia plantations managed by local people still exist in several areas in North Sulawesi. Examples include Rumoong Atas Village, South Minahasa District [
17], and Kawatak Village, Minahasa District [
18]. The plantation in Rumoong Atas village has existed for decades, and the Cempaka trees in the region were planted on inheritance land [
17]. The plantations typically cover about 1–2 ha and is managed from generation to generation. While the local community in Kawatak village developed plantations under the Community Forestry Program, the local people planted several
Magnolia species, including the endemic species,
M. sulawesiana.Habitat preference data will serve as the basic information for landscape management approaches to ensure the survival of the genus, including in-situ and ex-situ conservation. This approach is also expected to maintain the remaining natural population in the protected area while ensuring sustainable use through plantation and community forests. The
Magnolia species, especially the endemic species, found inside conservation areas, need to be protected in-situ [
22]. As an endemic and endangered species,
M. sulawesiana also needs to be considered as a protected species. For this reason, conservation efforts were conducted at both the habitat and species level. The other
Magnolia species found outside the conservation areas could also be proposed for protection to maintain their sustainability. Additionally, the area could be designated as a buffer zone. Using the same information, the ex-situ conservation of the species could be conducted through the development of plantations or community forests within the most suitable habitat preferences in collaboration with the local people. Community forest development could act as a buffer for the natural habitat of the species in the conservation area. Ex-situ conservation could also lessen the risk of extinction for threatened species and support in-situ conservation efforts [
22]. The remaining forest in Sulawesi plays a crucial role as a life support system due to its geographical conditions, with extreme faults being prone to landslides. Rapid changes in land use create further difficulties for conservation efforts on this island. A spatial distribution map is crucial for the local forest district to develop landscape-scale protection [
22], which is important not only for the targeted species but also for Wallace’s unique wildlife and Sulawesi’s fragile ecosystems more broadly [
45].
To facilitate the effective implementation of conservation, especially for endemic species with unique habitats [
46], further research needs to be conducted to determine the genetic diversity of all populations, and inbreeding and genetic diversity levels [
22,
46] could be used to determine protection priorities, especially at the landscape level. Information on the population size, phenological patterns, morphological variations [
47,
48], and population genetic diversity [
49] in natural habitats, including the populations in the northern and central part of Sulawesi, will determine the actions that should be taken concerning conservation in natural habitats (in-situ). This conservation should involve the indigenous knowledge of “
Eluren Eng Kayobaan” (keeping and maintaining the Earth) [
50] to encourage the planting of Wasian trees on community lands.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}