
The Influence of the Provenance and Spatial Structure on the Growth of European Silver Fir (Abies alba Mill.) of Autochthonous Origin in a Forest Plantation in the Białowieża Forest

 , , , , and
, , , , and 
Abstract
1. Introduction
- For the provenance factor:
- to evaluate the variability of the most important silvicultural parameter—DBH—and thus reveal the degree of possible differentiation both between the families of the studied silver fir half-sibs and within families;
- to identify the most productive phenotypes, which are potential sources of vegetative and seed material for effective reforestation of the autochthonous population of silver fir and objects of further genetic studies;
- For the spatial structure factor:
- to determine the parameters of the spatial structure effectively affecting the DBH,
- to determine the approaches of effective regulation of the spatial structure of the stand in the near term.
2. Materials and Methods
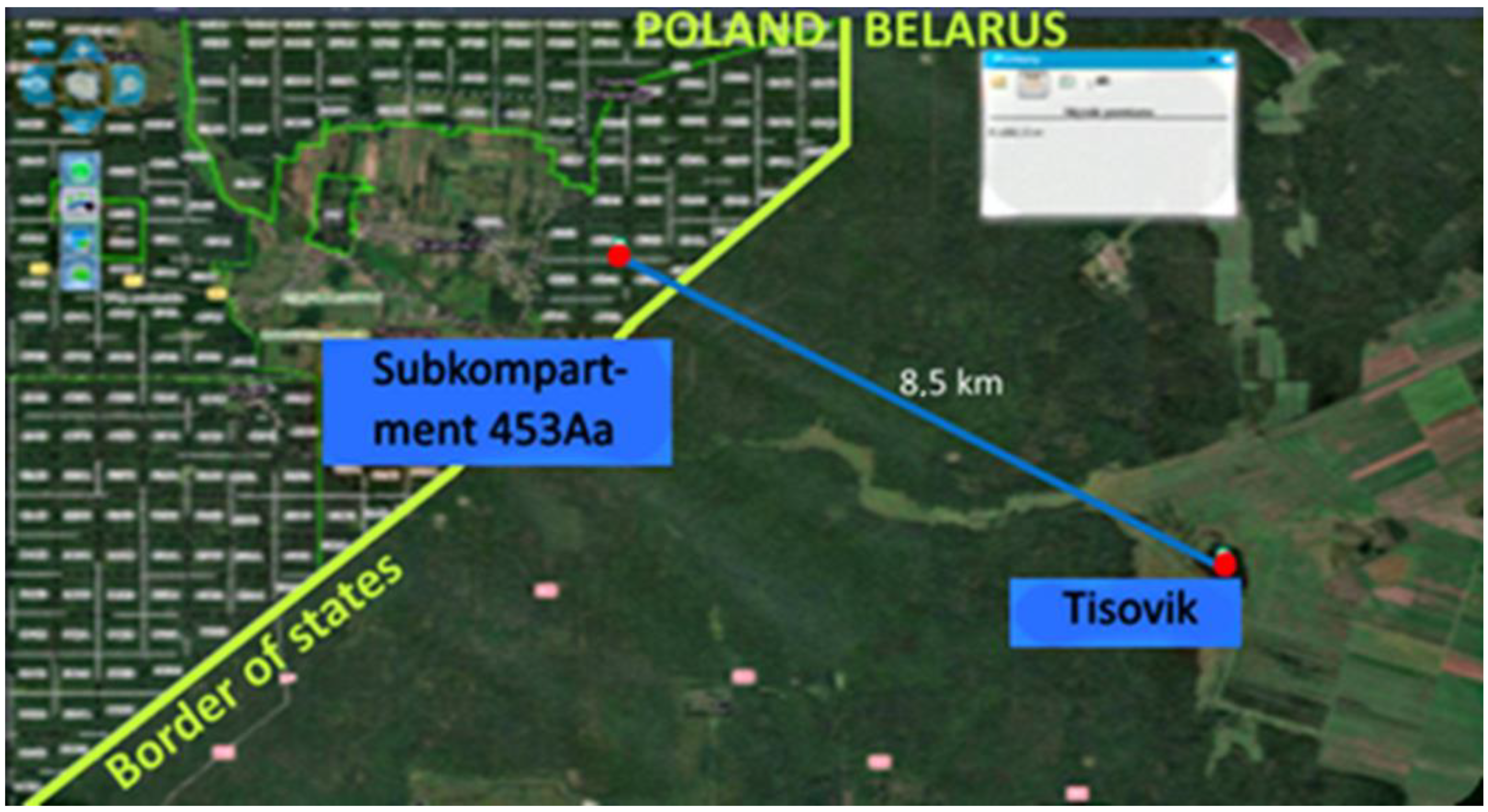
2.1. Object of Research
2.2. Collection of Field Data and Its Standard Processing
2.3. Methodological Approach for Determining the Influence of a Provenance Factor
2.4. Methodological Approach for Determining the Influence of the Factor of Spatial Structure
3. Results
3.1. The Influence of the Provenance Factor
3.2. The Influence of the Spatial Structure
3.3. Comparison of the Influence of Provenance Factor and Spatial Structure
3.4. Most Productive Phenotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brzeziecki, B.; Hilszczański, J.; Kowalski, T.; Łakomy, P.; Małek, S.; Miścicki, S.; Modrzyński, J.; Sowa, J.M.; Starzyk, J.R. Problem masowego zamierania drzewostanów świerkowych w Leśnym Kompleksie Promocyjnym “Puszcza Białowieska”. Sylwan 2018, 162, 373–386. (In Polish) [Google Scholar]
- Nowakowska, J.A.; Hsiang, T.; Patynek, P.; Stereńczak, K.; Olejarski, I.; Oszako, T. Health assessment and genetic structure of monumental Norway spruce trees during a bark beetle (Ips typographus L.) outbreak in the Białowieża Forest District, Poland. Forests 2020, 11, 647. [Google Scholar] [CrossRef]
- Szafer, W. Jodła w Puszczy Białowieskiej [Fir in the Białowieza Primeval Forest]. Sylwan 1920, 38, 65–74. (In Polish) [Google Scholar]
- Paczoski, J. Lasy Białowiezy [Forests of Białowieża]; Państwowa Rada Ochrony Przyrody: Poznań, Poland, 1930. (In Polish)
- Karpiński, J.J. Białowieża; Drukarnia państwowa w Płocku: Warszawa, Poland, 1947. (In Polish)
- Korczyk, A.F.; Kawecka, A.; Martysevič, V.V.; Strelkov, A.Z. European fir (Abies alba Mill.) natural stand in Białowieża Primeval Forest. Pr. Inst. Badaw. Leśnictwa 1997, 837, 27–62. [Google Scholar]
- Ermochin, M.V.; Barsukova, T.L.; Knys, N.V.; Byśko, V.E.; Bernackij, D.I. Dinamika i sostojanie populjacii pichty beloj v uročišče Tisovik [Dynamics and state of the silver fir’s population in the Tisovik tract]. In Belovezhskaia Pushcha. Research Compendium of Scientific Articles; Buryj, A., Arnolbik, V., Cherkas, N., Bunevich, A., Eds.; Alternativa: Brest, Belarus, 2016; pp. 65–88. (In Russian) [Google Scholar]
- Korczyk, A.F. Ocena wartości genetycznej i hodowlanej naturalnych populacji jodły pospolitej (Abies alba Mill.) ze wschodniego zasięgu w Polsce [Assessment of the genetic and breeding value of natural populations of silver fir (Abies alba Mill.) from theeastern range in Poland]. Zesz. Nauk. Akad. Rol. Im. H. Kołłątaja W Krakowie 1999, 61, 155–170. (In Polish) [Google Scholar]
- Cherkas, E.V.; Morozov, O.V. Influence of the oak red (Quercus rubra) on components of wood plantings of the SNPE «Belovezhsky dense forest». In Problemy Lesowedenija I Lesowodstwa; INL NAN Belarusi: Gomel, Belarus, 2012; Volume 72, pp. 150–156. (In Russian) [Google Scholar]
- Cherkas, E.V.; Morozov, O.V. Population of a red oak (Quercus rubra L.) in the Bialowieza Forest. In Ekologija, Lesowodstwo i Lesoohotničje Hozjajstwo; Belarusian State Technological University: Minsk, Belarus, 2012; Volume 1, pp. 127–129. (In Russian) [Google Scholar]
- Marozau, A.; Mielcarek, M.; Krok, G.; Paluch, R.; Chiliński, K. European silver fir—An alternative for the dying Norway spruce in Białowieża Forest? Folia For. Pol. 2021, 63, 150–166. [Google Scholar] [CrossRef]
- Marozau, A.; Kotsan, U.; Kalishuk, A. Reintroduction of the European silver fir (Abies alba Mill.) in Białowieża Forest. Balt. For. 2021, 27, 121–131. [Google Scholar] [CrossRef]
- Strelkov, A.Z.; Romaniuk, I.G.; Dworak, L.E. Sostojanie i perspektiwy wosproizwodstwa i ochrany pichty beloj (Abies alba Mill.) w Belovežskoj pušče. In Sochranenie Biologičeskogo Raznoobrazija Lesow Belovežskoj Pušči [Conservation of the Biological Diversity of the Forests of Belovezhskaia Pushcha]; Lučkov, A.I., Tolkač, W.N., Parfenov, W.I., Sawickij, B.P., Januško, A.D., Eds.; Izdanie: Kameniuki-Minsk, Belarus, 1996; pp. 122–130. (In Russian) [Google Scholar]
- Parfenov, V.I.; Savčuk, S.S. Abies alba Mill. Red Book of the Republic of Belarus. Rare and Threatened Species of Wild Plants; Encyklapedyja im. P. Brouki; Belarusian State Technological University: Minsk, Belarus, 2015; pp. 25–26. (In Russian) [Google Scholar]
- Gončarenko, G.G.; Padutov, A.E.; Padutov, V.E.; Silin, A.E.; Bolsun, S.I. Opredelenie genetičeskogo raznoobrazija osnownych lesoobrazujuščich i redkich drewesnych vidov GNP «Belovežskaja pušča». In Sochranenie Biologičeskogo Raznoobrazija Lesow Belovežskoj Pušči [Conservation of the Biological Diversity of the Forests of Belovezhskaia Pushcha]; Lučkov, A.I., Tolkač, W.N., Parfenov, W.I., Sawickij, B.P., Januško, A.D., Eds.; Izdanie: Kameniuki-Minsk, Belarus, 1996; pp. 141–160. (In Russian) [Google Scholar]
- Longauer, R. Genetic variation of European silver fir (Abies alba Mill.) in the Western Carpathians. J. For. Sci. 2001, 47, 429–438. [Google Scholar]
- Kempf, M.; Zarek, M.; Paluch, J. The pattern of genetic variation, survival and growth in the Abies alba Mill. Population within the introgression zone of two refugial lineages in the carpathians. Forests 2020, 11, 849. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.M.; Stoica, E.; Birsan, M.V. Intraspecific growth response to drought of Abies alba in the Southeastern Carpathians. Forests 2021, 12, 387. [Google Scholar] [CrossRef]
- Pawlaczyk, E.M.; Bączkiewicz, A.; Buczkowska, K.; Bobowicz, M.A. Genetic variation of silver fir progeny from Tisovik Reserve population determined via microsatellite and isozyme markers. Folia For. Pol. 2017, 59, 45–58. [Google Scholar] [CrossRef][Green Version]
- Mejnartowicz, L. Cisovka—The relic population of Abies alba and its relationship to man-made silver-fir stands in Białowieża Primeval Forest. Acta Soc. Bot. Pol. 1996, 65, 319–328. [Google Scholar] [CrossRef][Green Version]
- Oliver, C.D.; Larson, B.A. Forest Stand Dynamics, Update Edition; John Wiley & Sons (Copyright now held by CD Oliver & BC Larson): Hoboken, NY, USA, 1996. [Google Scholar]
- Rossi, S.; Morin, H.; Gionest, F.; Laprise, D. Spatially explicit structure of natural stands dominated by black spruce. Silva Fenn. 2013, 47, 1–14. [Google Scholar] [CrossRef]
- Goreaud, F.; Loussier, B.; Marie Ange, N.B.; Allain, R. Simulating Realistic Spatial Structure for Forest Stands: A Mimetic Point Process; Interdisciplinary Spatial Statistics Workshop: Paris, France, 2004. [Google Scholar]
- Wang, H.; Zhang, G.; Hui, G.; Li, Y.; Hu, Y.; Zhao, Z. The influence of sampling unit size and spatial arrangement patterns on neighborhood-based spatial structure analyses of forest stands. For. Syst. 2016, 25, 56. [Google Scholar] [CrossRef]
- Mason, W.L.; Connolly, T.; Pommerening, A.; Edwards, C. Spatial structure of semi-natural and plantation stands of Scots pine (Pinus sylvestris L.) in Northern Scotland. Forestry 2007, 80, 567–586. [Google Scholar] [CrossRef]
- Corral-Rivas, J.J. Models of Tree Growth and Spatial Structure for Multi-Species, Uneven-Aged Forests in Durango (Mexico)—Cuvillier Verlag. Available online: https://cuvillier.de/de/shop/publications/2111 (accessed on 23 September 2021).
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 2012, 47, 583–621. [Google Scholar] [CrossRef]
- Conover, W.J.; Johnson, M.E.; Johnson, M.M. A comparative study of tests for homogeneity of variances, with applications to the outer continental shelf bidding data. Technometrics 1981, 23, 351–361. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Hodges, J.L.; Lehmann, E.L. Estimates of location based on rank tests. Ann. Math. Stat. 1963, 34, 598–611. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; EEA: Vienna, Austria, 2021.
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. Borchers DescTools: Tools for Descriptive Statistics; R Package Version 0.99.43; R Core Team: Vienna, Austria, 2021; pp. 1–648. [Google Scholar]
- Pohlert, T. PMCMRplus: Calculate Pairwise Multiple Comparisons of Mean Rank Sums Extended; R Package Version 1.9.2; R Core Team: Vienna, Austria, 2021; pp. 1–207. [Google Scholar]
- Haze, M. (Ed.) Zasady hodowli hodowli lasu. In Pincipes of Siviculture; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2012. (In Polish) [Google Scholar]
- Sierota, Z. Criteria and methods of estimation of trees and stands health state. Pr. Inst. Badaw. Leśnictwa 1998, 854, 28. (In Polish) [Google Scholar]
- Jaworski, A.; Paluch, J. Charakterystyka cech morfologicznych jodeł w drzewostanach o strukturze przerębowej Beskidów. Leśne Pr. Badaw. 2007, 3, 7–31. (In Polish) [Google Scholar]
- Nagimov, Z.J. Evaluation of methods for determining tree growth areas. Lesa Ural. I Chozajstwo W Nich 1999, 19, 82–98. (In Russian) [Google Scholar]
- Kotsan, V.V. Classification of trees on the basis of spatial structure in appointments thinning. Tr. BGTU 2015, 1, 24–27. (In Russian) [Google Scholar]
- Rogozin, M.V. Nutrion area of tree: Analysis of methods. Bull. Sci. Pract. 2018, 4, 34–37. (In Russian) [Google Scholar]
- Rogozin, M.V. The Structure of the Forest: Competition or Partnership? Perm State University: Perm, Russia, 2019; p. 222. (In Russian) [Google Scholar]
- Bühl, A.; Zöfel, P. SPSS: Iskusstvo Obrabotki Informatsii. Analiz Statisticheskikh Dannykh I Vosstanovlenie Skrytykh Zakonomernostey [SPSS: The Art of Information Processing. Statistical Data Analysis and Mining]; DiaSoftYuP: St Petersburg, Russia, 2005. (In Russian) [Google Scholar]
- Szymkiewicz, B. Tablice Zasobności I Przyrostu Drzewostanów Ważniejszych Gatunków Drzew Leśnych [Stands Abundance and Growth Tables of The Most Important Forest Tree Species]; PWRiL: Warszawa, Poland, 2001; p. 179. (In Polish) [Google Scholar]
- Jaworski, A.; Zarzycki, K. Ekologia. In Jodła Pospolita Abies Alba Mill. Nasze Drzewa Leśne; Białobok, S., Ed.; Państwowe Wydawn. Nauk: Warszawa-Pozanań, Poland, 1983; Volume 4, pp. 317–430. (In Polish) [Google Scholar]
- Bernadzki, E. Jodła pospolita. In Ekologia—Zagrożenia—Hodowla; Fonder, W., Ed.; PWRiL: Warszawa, Poland, 2008; p. 209. (In Polish) [Google Scholar]
- Jaworski, A. Hodowla Lasu. Charakterystyka Hodowlana Drzew I Krzewów Leśnych; PWRiL: Warszawa, Poland, 2019; Volume 3, p. 696. (In Polish) [Google Scholar]
- Gonczarenko, G.G. Analysis of Population Genetic Resources (on the Example of White Fir in Belarus); Gomelskij Gosudarstwennyj Universitet Imeni F. Skoriny: Gomel, Belarus, 2002; p. 112. ISBN 978-985-6234-78-4. (In Russian) [Google Scholar]
- Jaworski, A. Hodowla Lasu. Pielęgnowanie Lasu; PWRiL: Warszawa, Poland, 2020; Volume 2, p. 400. (In Polish) [Google Scholar]
- Bardzain, W. Strategia restytucji jody pospolitej (Abies alba Mill.) w Sudetach. Sylwan 2000, 2, 63–77. (In Polish) [Google Scholar]
- Zeide, B. Optimal stand density: A solution. Can. J. For. Res. 2004, 34, 846–854. [Google Scholar] [CrossRef]
- Assmann, E. Waldertragskunde. Organische Produktion, Struktur, Zuwachs und Ertrag Von Waldbeständen; BLV Verlagsgesellschaft: München, Germany, 1961; p. 490. [Google Scholar]
- Kobendza, R. O zarastaniu ściętych pni jodłowych. Acta Soc. Bot. Pol. 1932, 9, 123–153. [Google Scholar] [CrossRef][Green Version]
- Korczyk, A.F. The ancestral conservative tillage of silver fir in the ‘Tisovik’ reserve of the Białowieża Primeval Forest. For. Res. Pap. 2015, 76, 153–167. [Google Scholar] [CrossRef][Green Version]
- Tjabera, A.P. Issues of territorial distribution in pine stands. Lesn. Žurnal 1980, 5, 5–8. [Google Scholar]
- Będkowski, M.; Czacharowski, M.; Opanowska, J.; Patro, S.; Urbańczyk, E.; Bielak, K. Effect of planting method on the growth and silvicultural quality of silver fir (Abies alba Mill.) in the Bieszczady Mountains. Sylwan 2021, 165, 542–553. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Half-Sib Family | Mean and Standard Deviation of the DBH, cm | Median | Lower Confidence Interval | Upper Confidence Interval |
|---|---|---|---|---|
| 4 | 7.71 (3.36) | 7.75 | 6.2 | 9.3 |
| 3 | 8.46 (3.55) | 8.5 | 7.1 | 9.9 |
| 21 | 8.52 (4.68) | 8.5 | 4.95 | 11.9 |
| 5 | 8.63 (3.50) | 8.5 | 7.65 | 9.35 |
| 16 | 8.64 (2.40) | 8.68 | 6.7 | 10.5 |
| 11 | 9.97 (3.28) | 9.8 | 8.7 | 11.05 |
| 15 | 10.05 (4.15) | 9.8 | 9.2 | 10.45 |
| 2 | 10.13 (4.04) | 10.05 | 9.4 | 10.7 |
| 12 | 11.25 (3.23) | 10.99 | 8.95 | 13.45 |
| 17 | 11.18 (4.52) | 11.1 | 10.4 | 11.8 |
| Class | Half-Sibs (Deviation from the Mean Value of the DBH for the Entire Sample in %) | DBH | |
|---|---|---|---|
| Mean and Standard Deviation, cm | Coefficient of Variation,% | ||
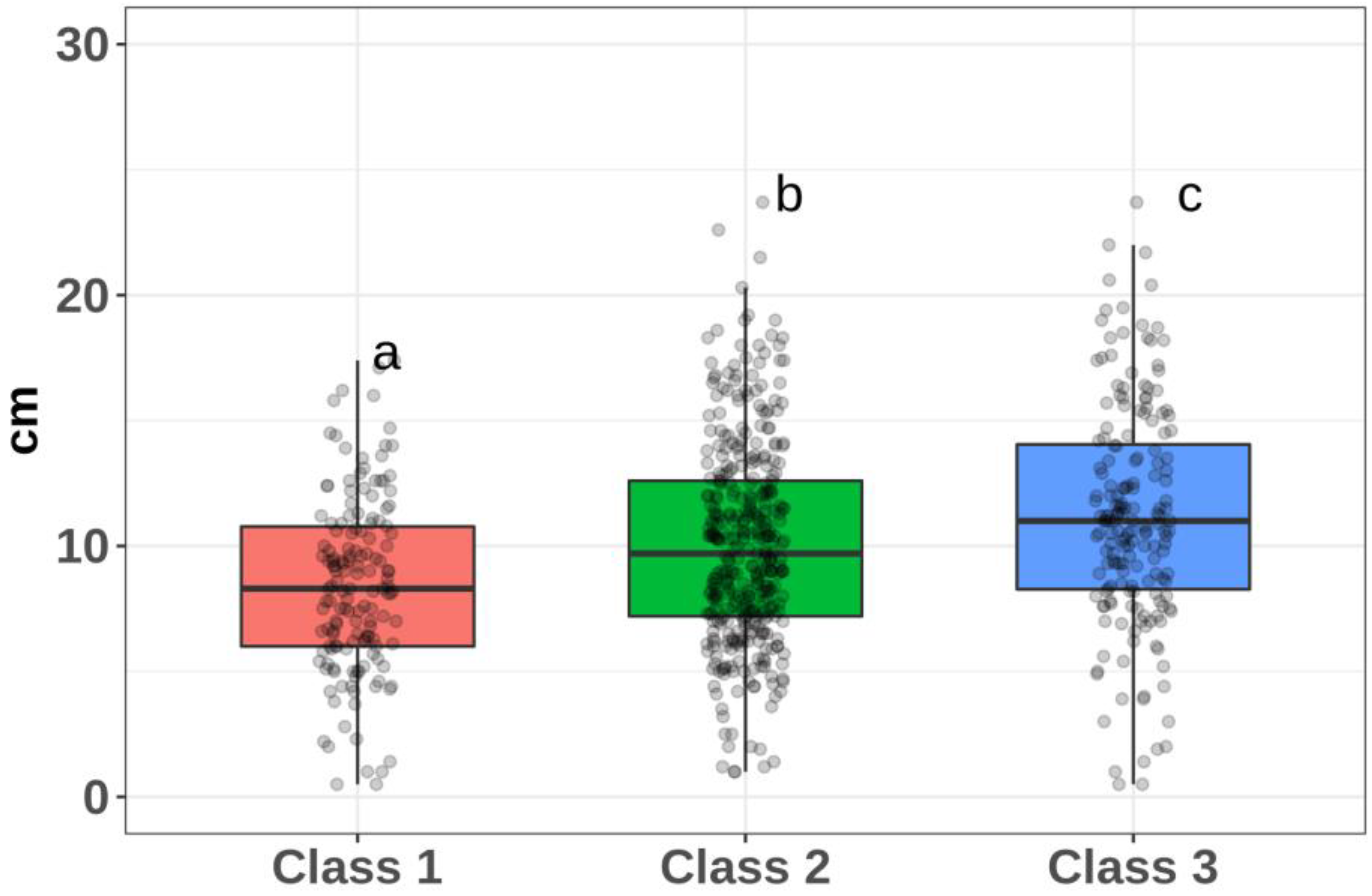
| 1 | 4 (−22.9), 3 (−15.4), 21 (−14.8), 5 (−13.7), 16 (−13.6) | 8.44 (3.49) | 41 |
| 2 | 11(−0.3), 15 (+0.5), 2 (+1.3) | 10.08 (4.02) | 40 |
| 3 | 12 (+12.5), 17 (+11.8) | 11.19 (4.43) | 37 |
| Indicator | Mean and Standard Deviation | Minimum Value | Maximum Value | Coefficient of Variation, % |
|---|---|---|---|---|
| Ln a | 2.13 (1.09) | 0.77 | 6.15 | 52 |
| Ls b | 1.94 (0.88) | 0.69 | 6.30 | 46 |
| Le c | 1.89 (0.72) | 0.58 | 3.75 | 38 |
| Lw d | 1.81 (0.73) | 0.59 | 4.26 | 41 |
| Lav e | 1.93 (0.49) | 1.10 | 3.42 | 26 |
| Class | Distance | DBH | ||||
|---|---|---|---|---|---|---|
| Class Boundaries | The Middle of the Class Interval | Mean and Standard Deviation, m | Coefficient of Variation, % | Mean and Standard Deviation, cm | Coefficient of Variation, % | |
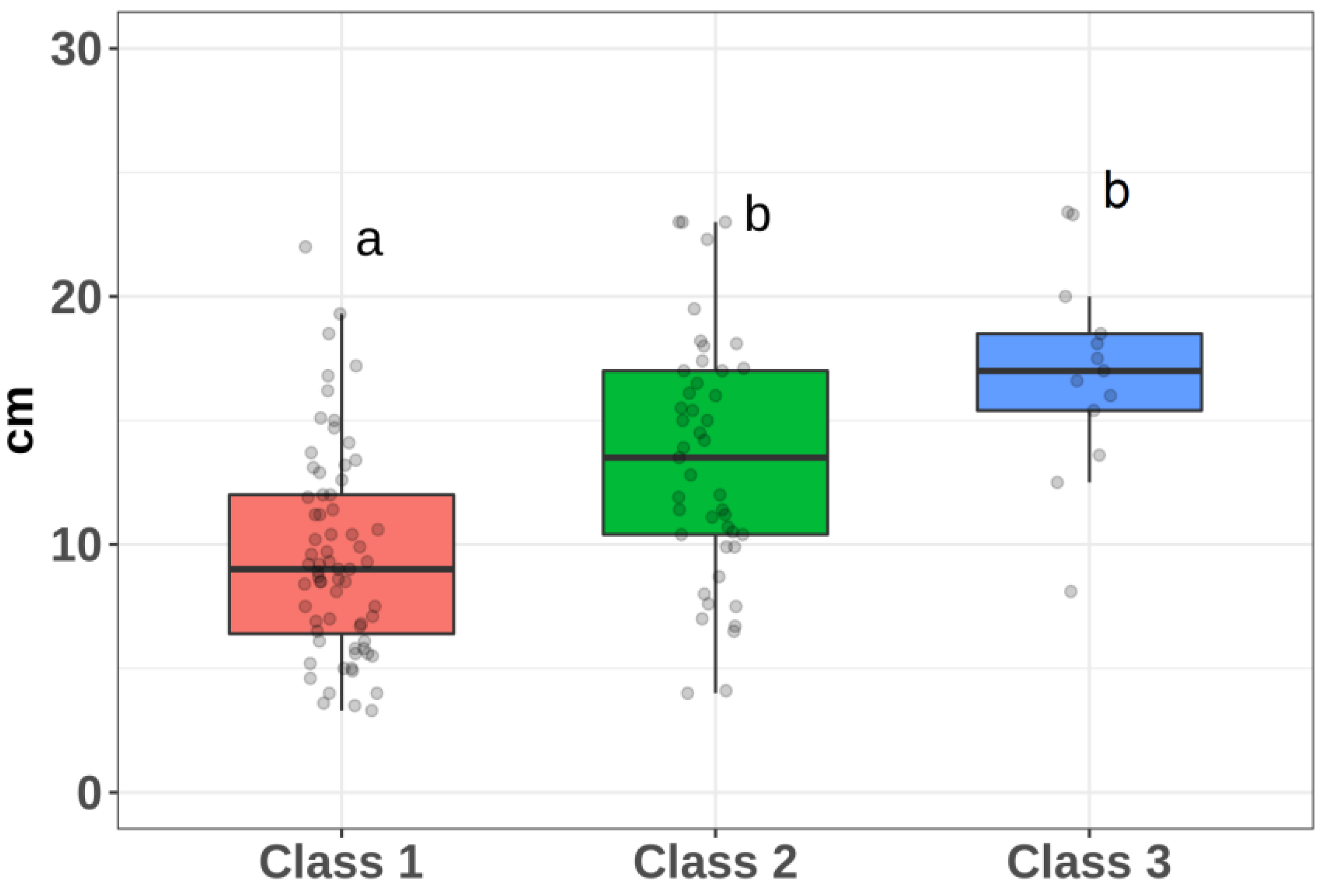
| 1 | 1.10–1.87 | 1.49 | 1.59 (0.19) | 12 | 9.6 (4.1) | 43 |
| 2 | 1.88–2.65 | 2.26 | 2.18 (0.20) | 9 | 13.4 (4.9) | 37 |
| 3 | 2.66–3.42 | 3.04 | 3.07 (0.21) | 7 | 16.9 (4.2) | 25 |
| Comparison Groups | p-Value |
|---|---|
| Class 1/Class 2 | 0.00015 |
| Class 1/Class 3 | 0.0000000072 |
| Class 2/Class 3 | 0.00227 |
| Class | N, pcs | Mode, cm | Hodges-Lehman Estimator | Lower Confidence Interval | Upper Confidence Interval |
|---|---|---|---|---|---|
| 1 | 150 | multi * | 8.4 | 7.81 | 8.99 |
| 2 | 371 | 9 | 9.9 | 9.49 | 10.35 |
| 3 | 180 | 11 | 11.1 | 10.45 | 11.75 |
| Comparison Groups | p-Value |
|---|---|
| Class 1/Class 2 | 0.000338039927 |
| Class 1/Class 3 | 0.000000019719 |
| Class 2/Class 3 | 0.12146648694 |
| Class | N, pcs | Mode, cm | Hodges-Lehman Estimator | Lower Confidence Interval | Upper Confidence Interval |
|---|---|---|---|---|---|
| 1 | 68 | 8.5 | 9.3 | 8.29 | 10.35 |
| 2 | 45 | 23 | 13.4 | 11.75 | 14.9 |
| 3 | 13 | multi * | 17.0 | 14.55 | 19.65 |
| Indicators * | Ln | Ls | Le | Lw | Lav |
|---|---|---|---|---|---|
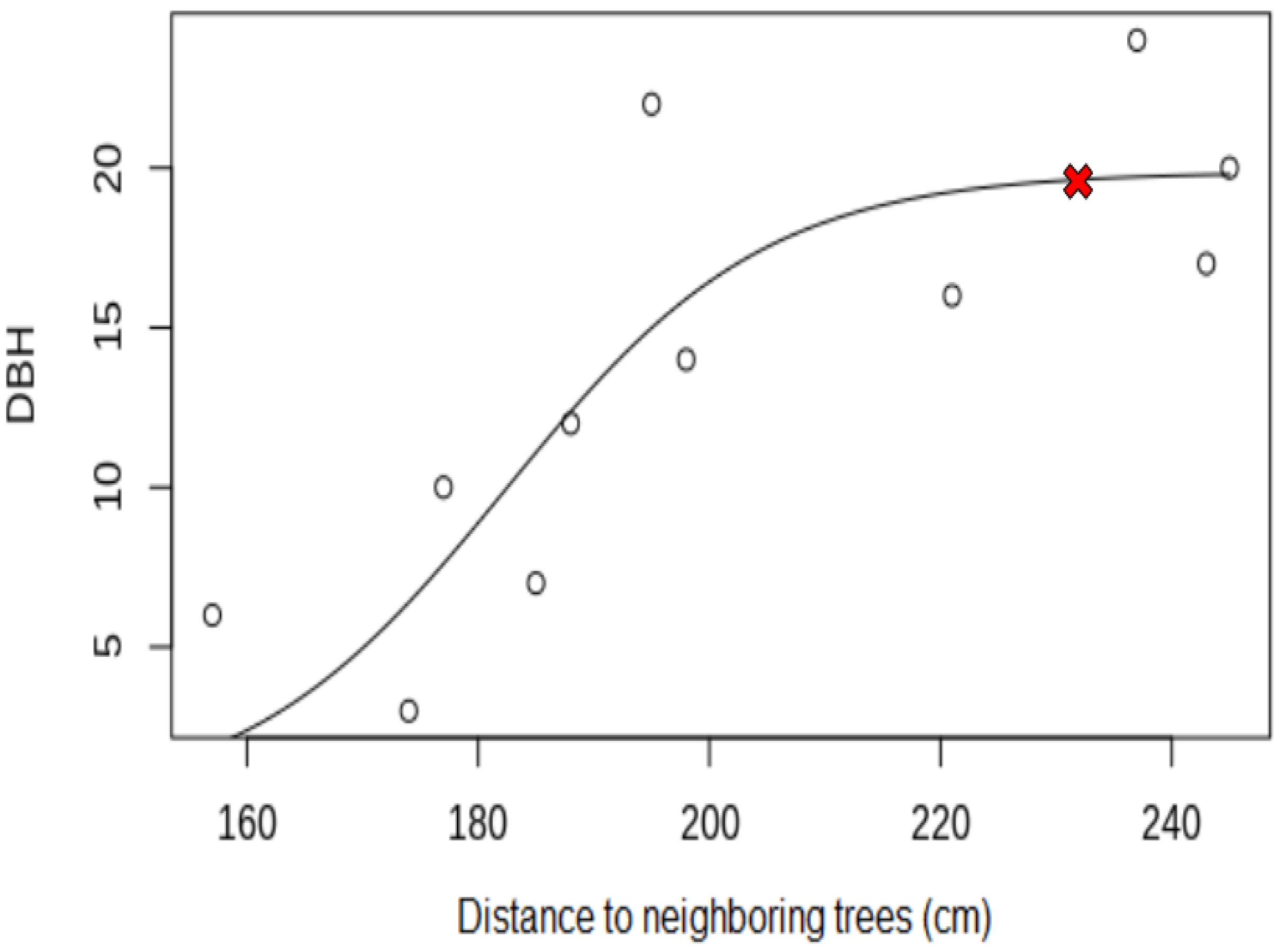
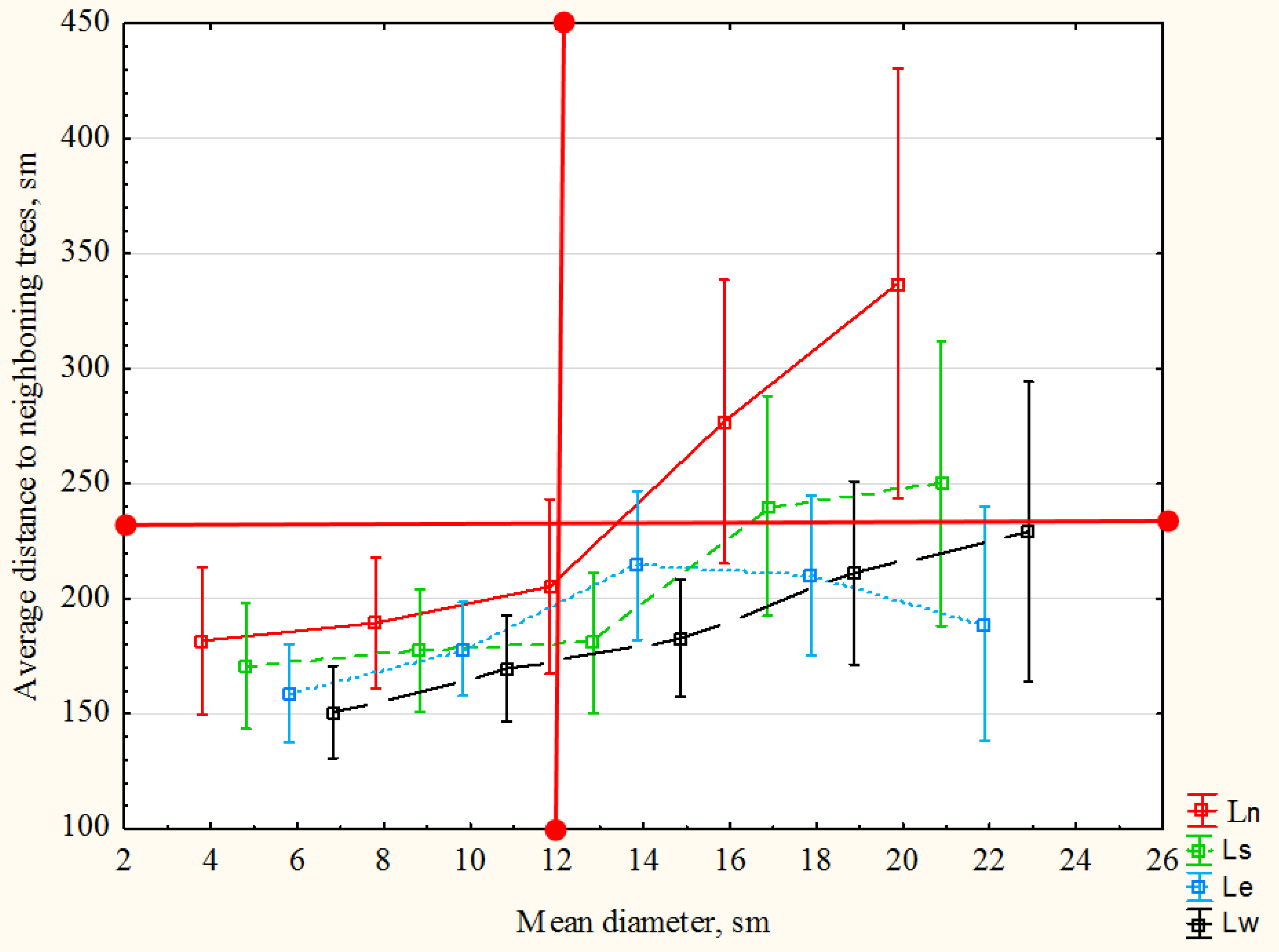
| Coefficient of correlation | +0.65 | +0.69 | +0.45 | +0.67 | +0.72 |
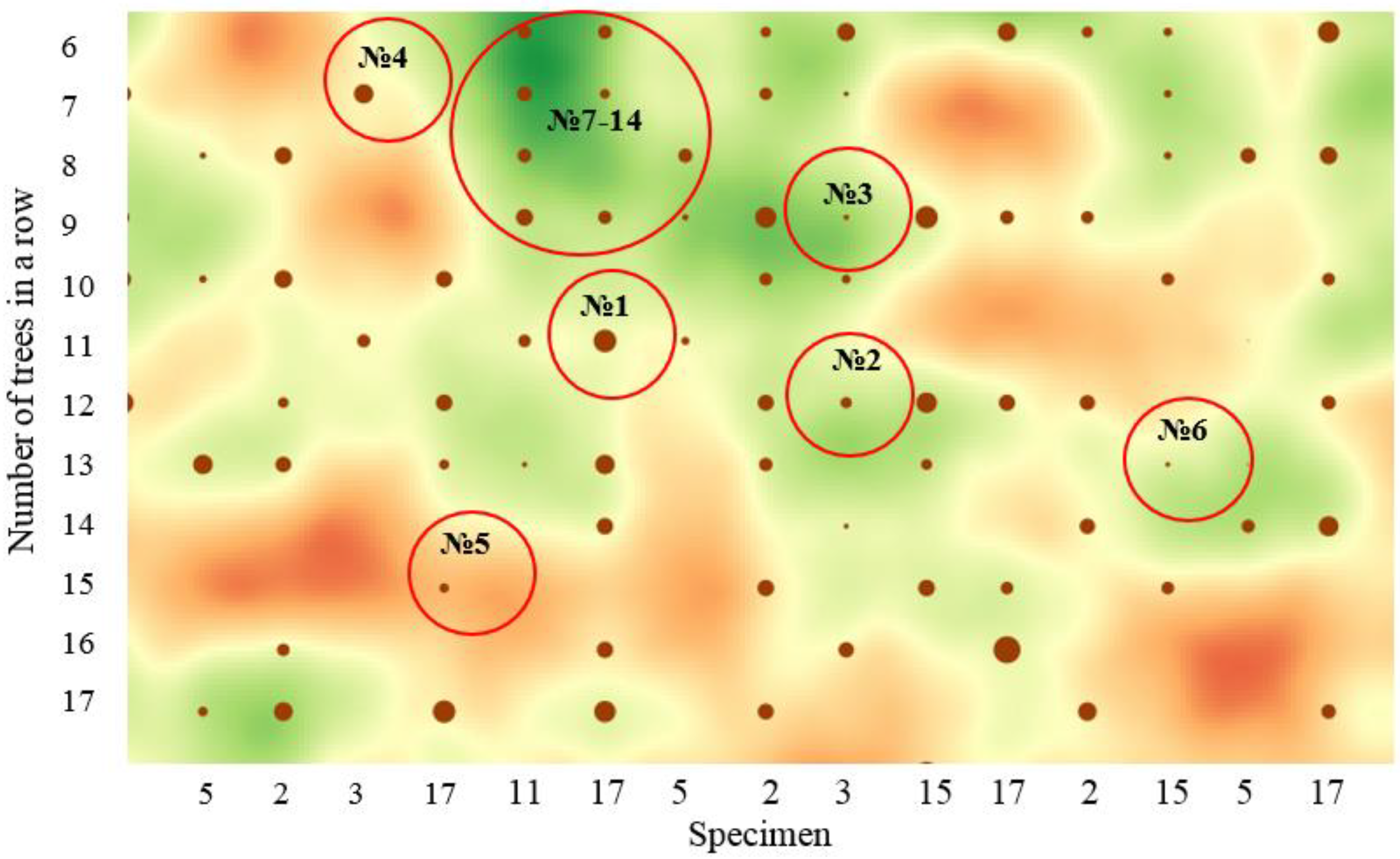
| Row | Tree Number | Half-Sib | DBH, cm | Tree Height, m | Average Distance to the Nearest Trees, m |
|---|---|---|---|---|---|
| 14 | 20 | 2 | 21.5 | 18.0 | 3.05 |
| 21 | 20 | 15 | 22.6 | 20.1 | 2.13 |
| 22 | 16 | 17 | 22.0 | 19.2 | 1.89 |
| 28 | 24 | 2 | 20.3 | 21.2 | 3.14 |
| 39 | 17 | 15 | 23.7 | 20.9 | 2.76 |
| 42 | 15 | 17 | 21.0 | 20.5 | 3.85 |
| 50 | 9 | 17 | 20.6 | 21.8 | 2.50 |
| 50 | 13 | 17 | 23.7 | 22.4 | 3.24 |
| 50 | 16 | 17 | 20.4 | 20.3 | 2.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marozau, A.; Kotszan, U.; Nowakowska, J.; Berezovska, D.; Moser, W.K.; Hsiang, T.; Oszako, T. The Influence of the Provenance and Spatial Structure on the Growth of European Silver Fir (Abies alba Mill.) of Autochthonous Origin in a Forest Plantation in the Białowieża Forest. Forests 2022, 13, 831. https://doi.org/10.3390/f13060831
Marozau A, Kotszan U, Nowakowska J, Berezovska D, Moser WK, Hsiang T, Oszako T. The Influence of the Provenance and Spatial Structure on the Growth of European Silver Fir (Abies alba Mill.) of Autochthonous Origin in a Forest Plantation in the Białowieża Forest. Forests. 2022; 13(6):831. https://doi.org/10.3390/f13060831
Chicago/Turabian StyleMarozau, Aleh, Uladzimir Kotszan, Justyna Nowakowska, Daria Berezovska, Warren Keith Moser, Tom Hsiang, and Tomasz Oszako. 2022. "The Influence of the Provenance and Spatial Structure on the Growth of European Silver Fir (Abies alba Mill.) of Autochthonous Origin in a Forest Plantation in the Białowieża Forest" Forests 13, no. 6: 831. https://doi.org/10.3390/f13060831
APA StyleMarozau, A., Kotszan, U., Nowakowska, J., Berezovska, D., Moser, W. K., Hsiang, T., & Oszako, T. (2022). The Influence of the Provenance and Spatial Structure on the Growth of European Silver Fir (Abies alba Mill.) of Autochthonous Origin in a Forest Plantation in the Białowieża Forest. Forests, 13(6), 831. https://doi.org/10.3390/f13060831