Abstract
In order to determine the relationships between bird assemblages and forest habitat, we conducted surveys for bird assemblages in different forest habitats in the Khentii Mountain region, Northern Mongolia. A total of 1730 individuals belonging to 71 species from 23 families of 11 orders were recorded. Our findings revealed that passeriformes are the most species-rich order, accounting for 86.2% of the total species. The dominant species were Anthus hodgsoni, Parus major, Poecile palustris, and Sitta europaea in study area. Non-metric multidimensional scaling (NMDS) and permutation multivariate analysis of variance (PERMANOVA) showed that bird assemblages were affected by forest habitat types. Our findings also showed significant relationships between bird assemblages and canopy height and ground cover vegetation structure, whereas there were no relationships between altitude and other habitat variables. Thus, maintaining diverse forest habitats or restoring forest would play a key role in bird conservation and sustainable management of forest areas.
1. Introduction
The boreal biome is located in the Northern Hemispheres, between 50° and 60° N latitudes [1]. The boreal forest covers northern Europe and Asia, and stretches from Far East Siberia to Scandinavia in the west [2]. Temperate conifers with varying proportions of deciduous trees dominate the boreal forest landscapes [2]. Mongolia is one of the world’s least forested regions [3]. Forest covers approximately 12% of Mongolia; 84% of the forest is mostly coniferous and deciduous, but also 16% saxaul forest [4,5,6]. In Mongolia, boreal forests mostly exist in the northern parts, which form a transitional zone between the Siberian Boreal Taiga and the Central Asian steppe [6]. These forests have relatively few tree species and are composed mainly of Siberian larch (Larix sibirica Ledeb.), Scots pine (Pinus sylvestris L.), Siberian pine (Pinus sibirica Du Tour.), Siberian fir (Abies sibirica Ledeb.), and Birch (Betula spp.), along with some deciduous tree species [5]. Mongolian boreal forest is characterized by a low human population density and a relatively low level of anthropogenic impact compared to other countries with Boreal forest regions [5]. Nevertheless, human population increases have led to an ever-increasing demand for forest products, timber harvesting, forest fires, increases in livestock numbers, degradation, and pests progressively depleting the forest cover [7,8]. Forest depletion totaled four million hectares in the last three decades, and the rate of deforestation increased to approximately 60,000 ha per year [5]. Deforestation by legal and illegal logging for especially conifer trees—sawmilling targets the largest trees—increased from the beginning of the 1970s [9,10]. Studies in the Boreal biome and tropical deforestation show that global bird diversity is tending to go down [11,12,13,14,15], and reforestation is a key action for bird conservation [16]. The relationships between bird assemblages and forest habitat have been the focus of many studies worldwide, and it is evident that forest habitat is an important determinant of the condition of bird assemblages in the boreal forest biome [17]. Bird species diversity has been shown to increases as forest tree diversity increases [18,19]. In addition, environmental variables such as habitat heterogeneity, canopy cover, tree size, and seasonal and climate changes are important effects on the growth and reproduction of the forest bird assemblages [20].
Most of the research is bird composition on boreal forest has concentrated on countries with boreal biomes, such as those in Europe, North America, and several Russian regions [2]. In Mongolia, the study of forest habitat and its biological communities (especially birds) has just started, and a limited number of studies on forest birds and their habitats are available. Most studies on bird diversity and biological communities have focused on waterfowl and threatened bird species, but there are only a few studies on the relationship between forest habitat and bird assemblages [10,21]. Unfortunately, ecological studies of the forest bird in the area are rare or have been mostly published in the Russian or Mongolian journals in their native languages and thus are hardly accessible to other scientists. Moreover, as noted by Bold’s (2003) [22], the early ecological finds from the forest bird assemblages of different habitats were reported based on bird species distribution as a result of a Russian–Mongolian joint expedition. His research focuses on bird species distribution and avifauna (Bold, 1973) [23]. In 1969, Bold [24] described some additional ecological and behavioral characteristics of some forest birds in different habitats of Mongolia. [25,26]. However, there are research gaps in Mongolia, especially regarding biological and ecological characteristics, such as forest bird species density, population dynamics, and the heterogeneity of micro habitat variables [10,21]. This his study aimed to quantify the forest structure, bird species composition, community assemblages, and occurrences of birds in different habitats of boreal forests, and to investigate the relationships between the avian community and forest conditions.
2. Materials and Methods
2.1. Study Sites
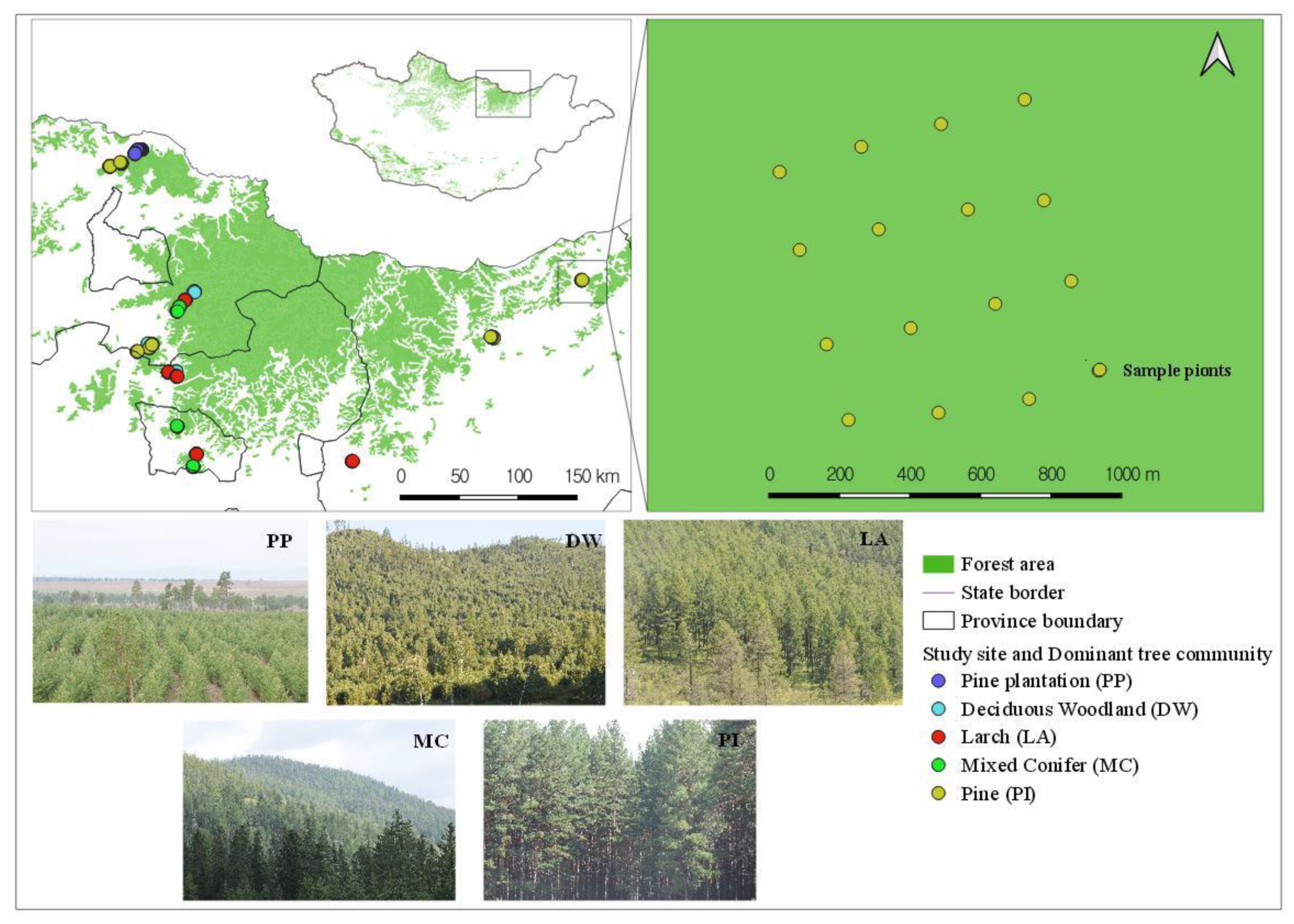
The research mainly focused on the surrounding regions of the Khentii Mountains (Figure 1). The Khentii Mountains are known as a forest region that is in northern Mongolia [27]. The annual average precipitation is 250–320 mm [3,28], and approximately 50–60% of precipitation is recorded in the summer [29]. The average annual temperature varies between −1.9 and −3.8 °C [30]. The forest of Khentii Mountains accounts for 33.5% (3755.2 ha, thousand) of the total forest area of Mongolia [24]. In the Khentii region, the western Siberian dark taiga forests with Spruce-Picea obovata, Fir-Abies sibirica, Pinus sibirica, and Siberian larch-Larix sibirica meet the eastern Siberian light taiga forests composed of species such as Birch-Betula platyphylla and related species, Larch-Larix sp. and Scotch pine-Pinus sylvestris [3,31,32,33]. Tree species of Khentii Mountain, especially mixed conifer and deciduous trees, and the plants, are relatively different in ecological regions from the other parts of Mongolia [34].
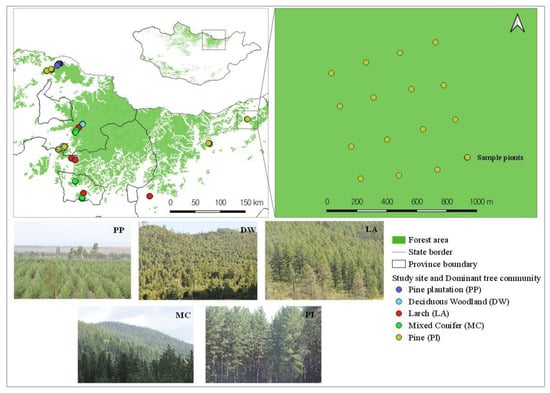
Figure 1.
Study area location and sampling sites in a forested area of the Khentii Mountains.
2.2. Sample Collection and Analyses
The survey was carried out at twenty plot sites on Kentii Mountain for native forested areas and five plots within a pine plantation area (Tujiin Nars). Five plots at plantation areas were selected based on the restorative time period since reforestation [35]. Bird surveys were conducted from June to August in 2019, a period recognized as breeding season for most species in the region [24]. All birds were counted and recorded via visual observation [36,37]. Each species was assigned to several functional groups based on migrations and diet preferences (Table 1) [21]. Field surveys were carried out from 6:00 to 11:00 AM during suitable weather conditions (without rain and with wind less than 5 m/s) and birds were recorded in point-counts of 10 min with a 50 or 80 m radius [36,37]. Survey plots were 1 km2 area with a 4 × 4 grid of 16 points, and a total of 400 point-counts were completed (Figure 1) [38,39]. The distance between points within plots was 250 m [37,38], and when we selected points at the forest’s edge, the distance between a point and the interior of the forest was 50 m. The occurrences of bird species and study sites were recorded using a global positioning system (GPS; German map62), and these positions were used to map in QGIS version 3.22 [40].

Table 1.
List of bird species and their abundances and occurrences in different habitat types in the forest of the Khentii Mountains.
We estimated forest tree (height, diameter at breast height—dbh) cover, and ground-cover variables (percentage of vegetation cover, bare ground, and down wood) of each point in a 50 m radius [39], and snag samples were 20 m in radius. Tree height was measured at ≥10 dominant trees at each point (around the center points). These data were grouped as mean values at the point-count level. The forest vegetation was sampled immediately after the end of bird counts. Forest characteristics were described as forest pattern, average tree height, cover, and tree component in the study area, the habitat was classified into five groups: the forest patterns and structure variables of each habitat type are shown in Table 2.
Table 2.
Structural variables were used to characterize the forest habitats of sampling sites where bird assemblages were described.
The relative abundance (RA) of a bird species was determined using the following expression: (number of individuals for species n/N total number of individuals) × 100%. To find differences in environmental factors and bird community attributes (i.e., species richness, abundance, and forest structure variables) among different habitat types, we performed analysis of variance (ANOVA) and Tukey’s post hoc test using the function aov in R software [41].
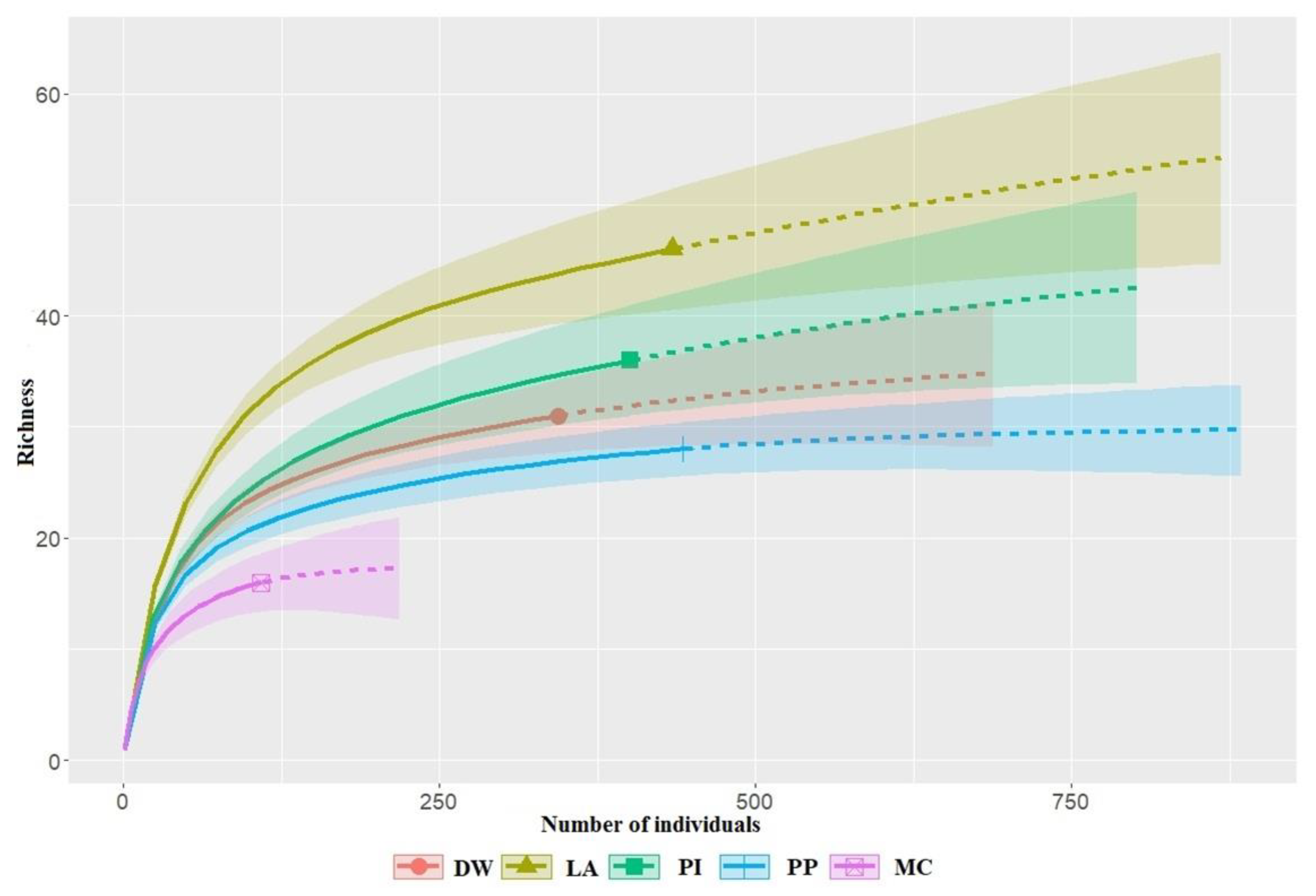
Differences in bird species richness among forest habitats were assessed through the rarefaction and extrapolation method based on sample coverage [42]. The species richness was calculated for each forest habitat based on the lowest sample coverage among the five habitats obtained within the 95% confidence intervals. This analysis was performed with the iNEXT and devtools package, using R [43,44].
To visualize the differences in bird assemblage composition between habitat types, we used permutational multivariate analysis of variance (PERMANOVA). Before performing PERMANOVA, the multivariate homogeneity of group dispersions was tested by the function betadisper, which indicated that there was a difference in dispersion between groups (F = 4.95, p < 0.01). The habitat type was used as an explanatory factor for PERMANOVA, which was tested using the function Adonis in the R package vegan [45]. The five forest types were considered a fixed effect in the analysis. Differences in species composition among samples collected in each site and forest type are presented in non-metric multidimensional scaling (NMDS) using the function metaMDS in the R package vegan 2.6-2 [46]. Moreover, similarity percentage (SIMPER) was also used to determine species that contributed most to the dissimilarities observed. All recorded species were included in the analysis. For SIMPER, we reported species that contributed to 83% of the bird community assemblages [45].
Redundancy analysis (RDA) was used as a direct gradient approach in order to determine how much variation in bird assemblages could be explained by environmental variables. Then, bird abundance data with total species were Hellinger transformed [47] using the function decosdtand in the R package vegan, in order to reduce the weights of abundant species while preserving the BrayCurtis index between samples in multidimensional space [45]. We performed RDA using the function RDA and tested the significance using the function ANOVA. In order to reduce the number of environmental variables entering the RDA, we used forward selection to get a parsimonious model. The forward selection was performed using the function ordiR2step with a permutation test (999 permutations) via the R package vegan [45]. The level of significance for all results was set to p < 0.05. All statistical analyses were performed using R version 3.5.1 [41].
3. Results
3.1. Bird Assemblage Composition
A total of 1730 birds were recorded during this study, belonging to 71 species from 23 families and 11 orders (Table 2). Of these, Passeriformes was the most species-rich order, accounting for 89% of the total species. The dominant species were Anthus hodgsoni—olive backed pipit (12.72%), Parus major—great tit (8.32%), Poecile palustris—marsh tit (7.34%), Sitta europaea—wood nuthatch (6.88%), Emberiza leucocephalos—pine bunting (6.07%), and Poecile montanus—willow tit (5.66%) which comprised 46.99% of the total bird count in this study. The dominant species varied among habitat types. For instance, A. hodgsoni, P. major, and P. montanus were generalists, which were dominant species in the most habitats. Moreover, E. leucocephalos was dominant in larix, pine, and pine plantation habitats, whereas P. palusris and S. europaea were dominant species in most habitats, but these species were not recorded in habitats with pine plantations. The high values of estimated sample coverage (ranges from 0.97 in LA and MC to 0.99 in PP) indicate that the sampling was sufficient to detect most species (Figure 2 and Table 3). In the forest habitats, species richness was higher in the larch (46 species) and pine-dominated habitats (31 species), compared to the pine plantation (28 species) and mixed conifer (16 species). We grouped the 71 species into four guilds: insectivores (38 species), omnivores (14), carnivores (13), and granivores (6) (Table 3). Insectivores were the most abundant group (68.09%), followed by granivores (14.85%), omnivores (12.2%), and carnivores (4.86%).
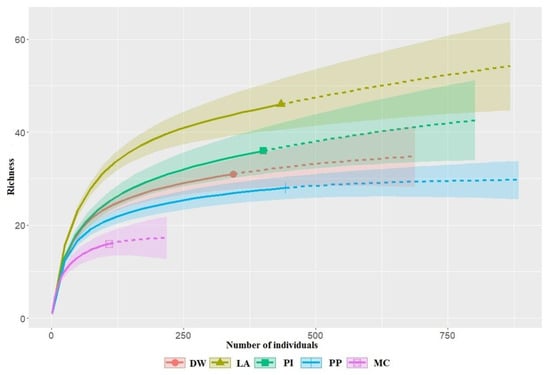
Figure 2.
Species richness of birds was recorded during this survey in the Khentii Mountains. Habitat variables are deciduous woodland (DW), larch (LA), pine (PI), pine plantation (PP), and mixed conifer (MC). Solid and dashed lines are interpolated and extrapolated data, respectively, based on rarefaction and extrapolation methods, with their associated 95% confidence intervals.
Table 3.
Bird richness in areas with different forest habitats in Khentii Mountains.
3.2. Correlations between Bird Assemblage and Habitat Types
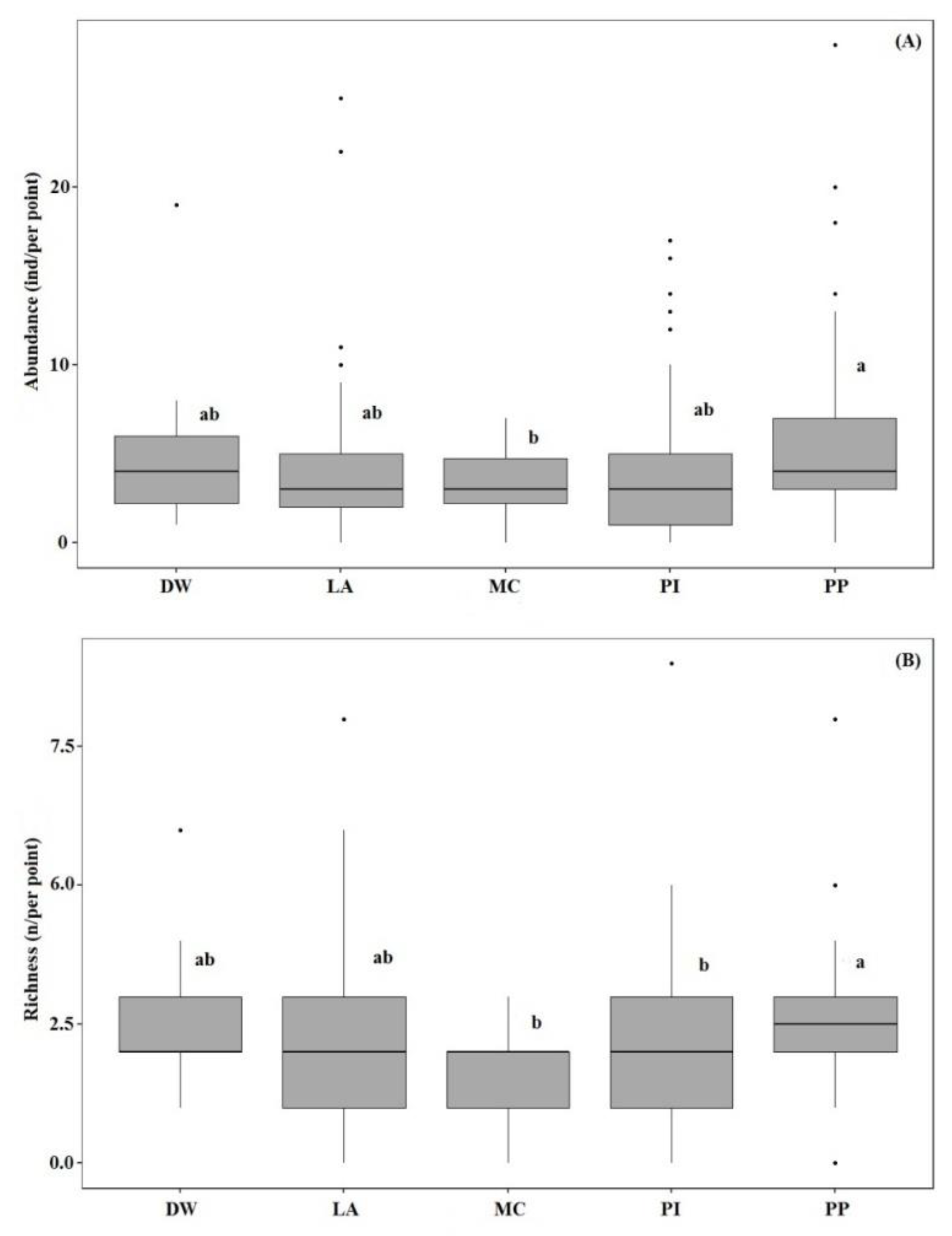
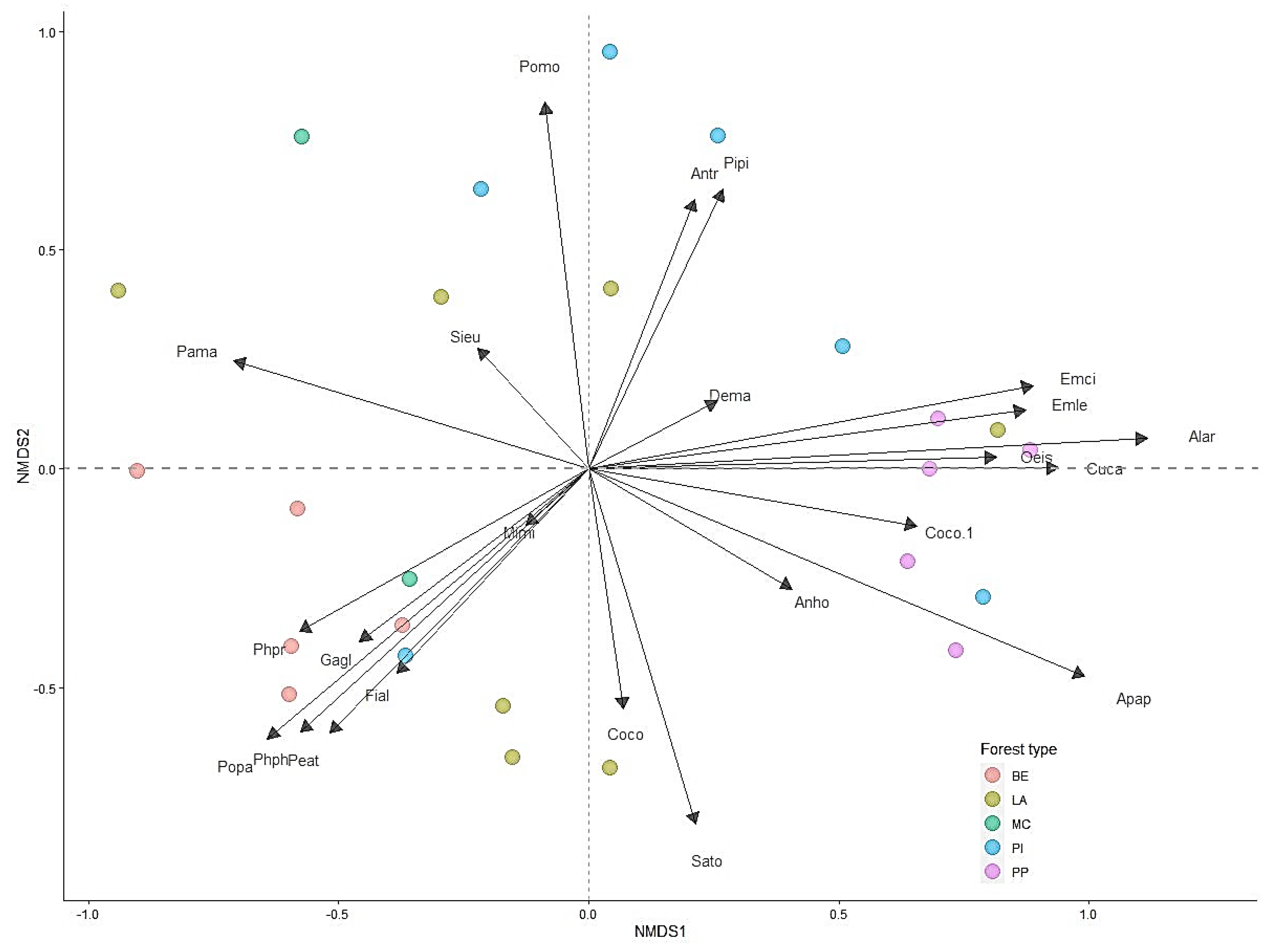
Results of one-way ANOVA showed that the bird abundance per point for pine plantations was significantly higher than that for mixed conifer forest habitats (F (4, 395) = 3.14, p < 0.05). Bird species richness per point in the pine plantation habitat was significantly higher than for other habitat types (F (4, 395) = 3.97, p < 0.05) (Figure 3A,B). Permutation analysis of variance and NMDS (stress = 0.22) revealed bird assemblages were significantly affected by habitat type (PERMANOVA, pseudo-F = 2.59, p < 0.001) (Figure 4). In addition, SIMPER analysis confirmed these dissimilarities and revealed that A. hodgsoni, P. major, P. palustris, E. leucocephalos, S. europaea, and P. montanus are major contributors to dissimilarities between habitat types (Table 4).
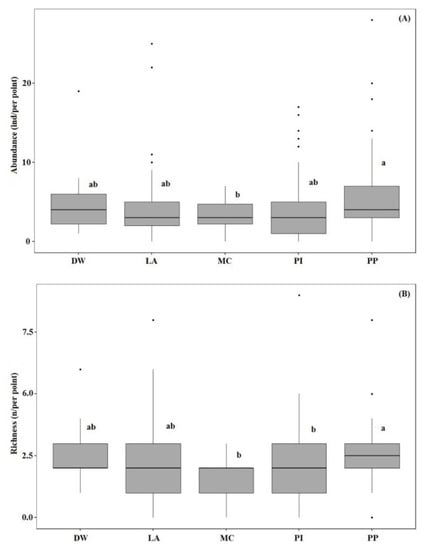
Figure 3.
Analysis of variance (ANOVA) for abundance (A) and species richness (B) of birds in the number of birds per point among selected habitats. Forest types are deciduous woodland (DW), larch (LA), pine (PI), pine plantation (PP), and mixed conifer (MC).
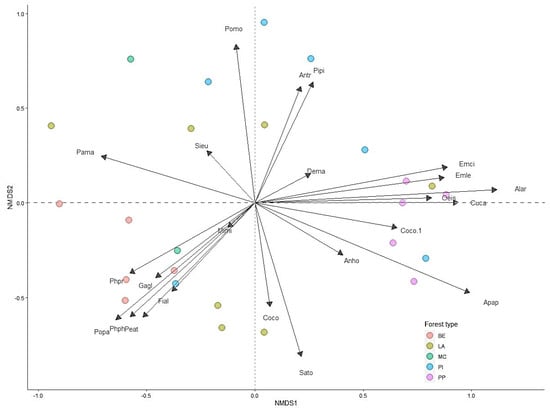
Figure 4.
Non-metric multidimensional scaling (NMDS) ordination of selected bird assemblages in different habitat types in the forest of the Khentii Mountains. Forest types are deciduous woodland (DW), larch (LA), pine (PI), pine plantation (PP), and mixed conifer (MC). Note: Species names are generic acronyms or the four letters of the scientific names in Table 1.
Table 4.
SIMPER analysis of dissimilarity among forest habitat types by most abundant species.
3.3. Relationships between Bird Assemblages and Environmental Variables
One-way ANOVA revealed that mixed conifer had the highest average altitude (1688.6 ± 204.6 m), followed by larch-dominant habitats (1374.7 ± 267.9 m), and the pine plantation habitat altitude was lower still (701.7 ± 18.3 m). The forest habitat types were significantly different at different altitudes (F4, 395 = 240.1, p < 0.01). The average numbers of snags of deciduous woodland and larch habitats were significantly higher than other habitat types (F4, 395 = 10.95, p < 0.05). The dbh sizes in deciduous woodland, larch, and mixed conifer habitats were significantly larger than in other habitat types (F4, 395 = 4.71, p < 0.05). The overstory, understory, and ground-level were significantly different in habitat structural variables (Table 5).
Table 5.
Results of one-way analysis of variance (ANOVA) with Tukey’s HSD test for environmental variables (mean ± SD) among different forest habitat types in the Khentii Mountains.
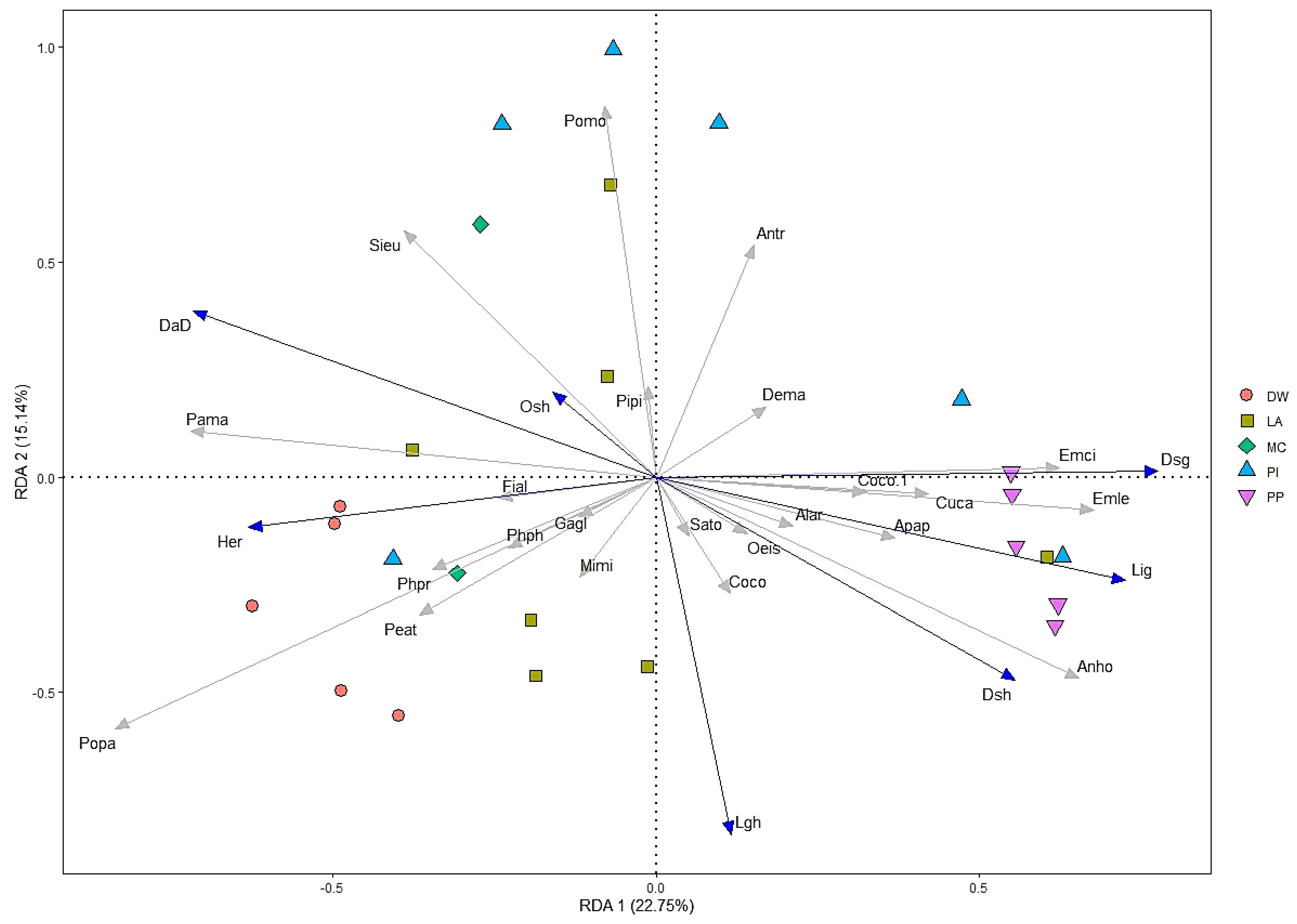
A total of 23 bird species were selected for redundancy analysis (RDA) with frequency of occurrence, and seven environmental variables were selected after a forward stepwise selection, including overstory, live grass, and dead grass height; and herbaceous, dead grass, live grass, and dead down wood cover. The first two axes (RDA1 and RDA2) accounted for 22.75% and 15.14% of the variation of 23 bird species, respectively (Figure 5). Different bird species preferred different environmental variables, which supports hypothesis ii. For instance, Sitta europaea, Parus major, and Pica pica were positively correlated with overstory height and dead wood, but negatively related to ground cover grass and height. Ficedula albicilla was positively related to herbaceous cover, but negatively related to dead standing grass cover. Moreover, Emberiza cioides, Emberiza leucocephalos, and Anthus hodgsoni also were positively correlated with grass cover dominant and pine plantation habitats, and negatively correlated with overstory height and dead down wood habitats.
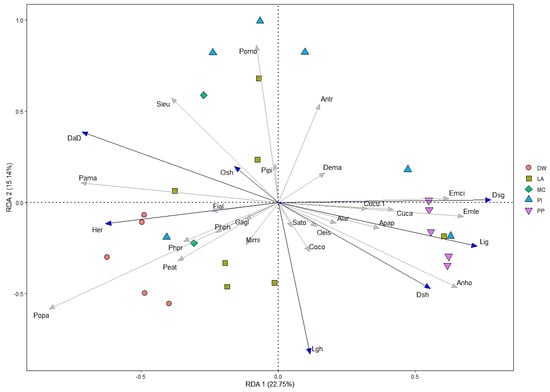
Figure 5.
Redundancy analysis (RDA) ordination among bird assemblage and habitat types and environmental variables in the forest of the Khentii Mountains. Forest types are deciduous woodland (DW), larch (LA), pine (PI), pine plantation (PP), and mixed conifer (MC). Note: Species and habitat structural variable names are generic acronyms or the four or three letters of the names in Table 1 and Table 2.
4. Discussion and Conclusions
Our study demonstrates the species richness and composition of bird assemblages, and the relations of forest habitat patterns of the Khentii Mountain boreal forest. Other studies have provided overviews of the taxonomic richness and composition of the avifauna across the Northern Mongolia [24,25,26,48]. Nevertheless, these only focused on avifauna, which resulted in forest regions remaining understudied. In total, 71 species were seen by the end of the survey, which were recorded in 400 point-counts, and on average, 3.14 species were seen per point. Birds in the study area account for 36.4% of the total species richness in the Khentii Mountains region and contribute to bird diversity [24], suggesting that forest habitat variables play an important role in bird diversity in forest areas [10,25]. Of this total, fourteen bird species occurred in all habitat types. Bird assemblages in the Khentii forested area were dominated mainly by Anthus hodgsoni, Parus major, Poecile palustris, Sitta europaea, Emberiza leucocephalos, and Poecile montanus. These species had relative abundance values of 5 to 12%, thereby contributing a lot to bird diversity in this area, accounting for 45% of total species richness. The bird species richness increased from the taiga zone to the forest-steppe zone. More specifically, our study found that bird richness was higher in larch dominated forests. Light mixed deciduous and larch forest communities have highly diverse bird assemblages, more so than taiga forest communities [24]. Larch forests, especially those burned by wildfires, change their landscapes and increase the forest layers of young birch, shrubs, and grasses [3]. The presence of deciduous trees in conifer-dominated forests generally allows for higher bird diversity compared to pure conifer stands [49,50].
In this study, forest habitat variables (altitude, overstory, understory, and ground composition) changed markedly by forest type. Different habitats are essential for bird assemblages in this forested area. The total species richness in the Khentii forested area was found to play an important role in the breeding and growth of the bird community, and a refuge for bird assemblages in the breeding season, for 57.3% of Mongolian bird species [25,26]. Among all types of habitats, the avifauna abundances were similar: the resident and migrant species accounted for 47.2% and 52.8%, respectively. The guild types also have a certain impact on the habitat selection of birds [25]; thus, insectivores (DW, LA, PI, PP, MC) were the dominant guild within each habitat type, and in our study area, there were 38 (53.5%) insectivores out of the 71 species sampled. The researchers have mentioned that insectivores are commonly observed to dominate forest habitats in terms of species richness and abundance; they are very important to forest regeneration [25,26,51].
Moreover, the present study showed bird assemblages in forest habitats of the Khentii Mountains were structured by a combination of overstory height and ground-layer features (e.g., dead down wood, herbaceous, and grass). Among these, ground layer-factors acted as the most important contributors to forest habitat types. The importance of forest habitat heterogeneity to the compositions of bird assemblages has been demonstrated by numerous studies [51,52,53,54,55]. The habitat variability and the diversity of the bird community are strongly influenced by the structure of the vegetation heterogeneity [56,57]. Our study found a significant difference in the tree and vegetation cover between forest type and canopy layer-based forested areas. Thus, in our study areas, ground-nesting bird species associated with grass (e.g., Anthus hodgsoni, Emberiza leucocephalos, Anthus trivialis, Alauda arvensis, and Emberiza cioides) tended to occupy areas with tall grass, such as pine plantations, and pines were often high in these habitats. The species that prefer an open forest canopy structure often tend to dominate the assemblages of dry Scots pine stands with sparse crown cover [20,58]. The high canopy-dominated conifer tree sites (LA, PI, MC) provide habitats for secondary cavity-nester bird species associated with low grasses and herbaceous cover (e.g., Sitta europaea, Parus major, and Poecile montanus). Those habitats are the most abundant in dead and down trees. Moreover, these species did not occur in pine plantation sites, and some secondary cavity-nester species were less abundant. Natural mature forests are highly suitable for cavity nesters or bark feeding species [59,60] and the abundance of the insectivores responds positively to an increase in basal area or dead wood volume [11]. On the other hand, the Phylloscopus proregulus, Poecile palustris, and Periparus ater have been recorded in high-herbaceous-cover areas dominated by deciduous woodland and some conifer habitats. As well, four species (Poecile palustris, Periparus ater, Phoenicurus phoenicurus, and Ficedula albicilla) are hole-nesters, and two species are low canopy nesters (Phylloscopus proregulus, and Garrulus glandarius), suggesting that the presence of deciduous trees is suitable for nesting for these species. Especially, species that generally require the presence of deciduous trees within stands are woodpeckers, tits, some nuthatches, and songbirds [54,61,62]. In deciduous woodland and deciduous mixed conifer forests, tits and warblers represented by far the most abundant portions of the community, and in pine and pine plantations, Emberiza species were more abundant than tits. However, woodpeckers were often the smallest fraction of the community. Our results showed that some species are more associated with sites with taller vegetation, whereas other species occupy sites with shorter vegetation and diverse herbaceous cover.
This study was the first to describe the forest-type variations of bird assemblages in the Khentii Mountains forest in Northern Mongolia. Identifying the assemblage pattern is useful for the conservation of not only birds but also biodiversity in general. In the course of the investigation, 71 bird species from 23 families and 11 orders were registered in the forest habitats of the Khentii partially of the forest area, and the Passeriformes order dominated; this result is the most up-to-date and systematically collected baseline data for future forest bird research. Information obtained from this study will enhance our understanding of the variation in bird assemblages, and then help to develop strategies for future forest bird conservation in such areas. Among these, insectivore birds dominate in all forest habitats. Many studies have also shown that insectivorous birds are more sensitive to habitat disturbance and loss than other feeding guilds [51,55,62,63,64]. From 2004 to 2014, the burned forest in the area increased by 1.4 million hectares, and the logging and insect-affected forest area increased by 340 thousand hectares [6]. Unfortunately, at the same time, the reforestation area increased by 2 thousand hectares [6], which shows a lack of reforestation. The bird community structure was affected by many factors, such as vegetation, the size and structure of the forest, and forest type.
In conclusion, according to our findings, Khentii forest areas are outstanding sites for migratory and resident birds. The results from this study show the importance of forest habitat structure for the abundance and diversity of birds in mixed tree and conifer forests. The bird species diversity and distribution of Khentii region can be currently regarded as moderately well studied. However, considering the bird assemblages of forested regions in Mongolia, further study is needed to fine-tune the species population estimates. In the future, we aim to study the co-effects of habitat disturbance and temporal variation on bird communities and bird density in different forest habitats. Thus, further studies on the relationship between temporal variation and bird density are important not only for ecological theory, but also for the scientific fundamentals of forest management and environmental protection in Mongolia.
Author Contributions
This paper received individual contributions from each author as specified: Conceptualization, Z.P. and J.W.L.; Methodology, Z.P.; Software, Z.P.; Validation, Z.P., J.W.L., W.K.P. and O.G.; Formal Analysis, Z.P.; Investigation, Z.P., J.W.L., M.M., O.G., U.G., E.P., B.N., T.A. and A.J.; Resources, Z.P. and J.W.L.; Data Curation, Z.P. and U.G.; Writing—Original Draft Preparation, Z.P.; Writing—Review and Editing, Z.P., O.G., M.M. and J.W.L.; Visualization, Z.P. and J.W.L.; Supervision, J.W.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the research fund of Chungnam National University: CNU;2020-0532-01.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data may be available upon request to the corresponding author, subject to the approval.
Acknowledgments
The authors appreciate the researchers and students of the Mongolian National University of Education, who participated in the field work. We would also like to thank our research colleagues from the Mongolian National University of Education (MNUE). J. Ariunbold partially supported.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Roberge, J.M.; Virkkala, R.; Monkkonen, M. Boreal 6 r Forest Bird Assemblages and Their Conservation. Ecol. Conserv. For. Birds 2018, 29, 183. [Google Scholar] [CrossRef]
- Dugarjav, C.H. Larch Forests of Mongolia; Bembi San: Ulan Bator, Mongolia, 2006; p. 317. (In Mongolian) [Google Scholar]
- FAO. Mongolia—Global Forest Resources Assessment 2015—Country Report; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy, 2014; p. 97. [Google Scholar]
- Batkhuu, N.O.; Lee, D.K.; Tsogtbaatar, J. Forest and forestry research and education in Mongolia. J. Sustain. For. 2011, 30, 600–617. [Google Scholar] [CrossRef]
- Government of Mongolia. Mongolia’s Forest Reference Level Submission to the United Nations Framework Convention on Climate Change; UN-REDD Mongolia National Programme, Ministry of Environment and Tourism: Ulaanbaatar, Mongolia, 2018; p. 62. [Google Scholar]
- Tsogtbaatar, J. Deforestation and reforestation of degraded forestland in Mongolia. In the Mongolian Ecosystem Network; Springer: Tokyo, Japan, 2013; pp. 83–98. [Google Scholar] [CrossRef]
- Sainnemekh, S.; Isabel, C.B.; Bulgamaa, D.; Brandon, B.; Ása, L.A. Rangeland degradation in Mongolia: A systematic review of the evidence. J. Arid. Environ. 2022, 196, 104654. [Google Scholar] [CrossRef]
- Erdenechuluun, T. Wood Supply in Mongolia: The Legal and Illegal Economies. In Mongolia Discussion Papers, East Asia and Pacific Environment and Social Development Department; World Bank: Washington, DC, USA, 2006. [Google Scholar]
- Wildlife Science and Conservation Center, Institute of Biology and BirdLife International. Directory of Important Bird Areas in Mongolia: Key Sites for Conservation; Nyambayar, B., Tseveenmyadag, N., Eds.; Wildlife Science and Conservation Center, Institute of Biology and BirdLife International: Ulaanbaatar, Mongolia, 2009. [Google Scholar]
- Collar, N.J.; Crosby, M.J.; Stattersfield, A.J. Birdlife International. Birds to Watch 2: The World List of Threatened Birds: The Official Source for Birds on the Iucn Red List; BirdLife International: Cambridge, UK, 1994. [Google Scholar]
- Balestrieri, R.; Basile, M.; Posillico, M.; Altea, T.; De Cinti, B.; Matteucci, G. A guild-based approach to assessing the influence of beech forest structure on bird communities. For. Ecol. Manag. 2015, 356, 216–223. [Google Scholar] [CrossRef]
- Kamp, J.; Oppel, S.; Heldbjerg, H.; Nyegaard, T.; Donald, P.F. Unstructured citizen science data fail to detect long-term population declines of common birds in Denmark. Divers. Distrib. 2016, 22, 1024–1035. [Google Scholar] [CrossRef]
- Donald, P.F.; Green, R.E.; Heath, M.F. Agricultiral intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. Lond. 2001, 268, 25–29. [Google Scholar] [CrossRef]
- Brinkert, A.; Hölzel, N.; Sidorova, T.V.; Kamp, J. Spontaneous steppe restoration on abandoned cropland in Kazakhstan: Grazing affects successional pathways. Biodivers. Conserv. 2016, 25, 2543–2561. [Google Scholar] [CrossRef]
- Kwok, H.K.; Corlett, R.T. The bird communities of a natural secondary forest and a Lophostemon confertus plantation in Hong Kong, South China. For. Ecol. Manag. 2000, 130, 227–234. [Google Scholar] [CrossRef]
- Lott, C.A.; Akresh, M.E.; Costanzo, B.E.; D’Amato, A.W.; Duan, S.; Fiss, C.J.; Fraser, J.S.; He, H.S.; King, D.I.; McNeil, D.J.; et al. Do Review Papers on Bird–Vegetation Relationships Provide Actionable Information to Forest Managers in the Eastern United States. Forests 2021, 12, 990. [Google Scholar] [CrossRef]
- Thompson, P.S.; Greenwood, J.D.; Greenaway, K. Birds in European gardens in the winter and spring of 1988–89. Bird Stud. 1993, 40, 120–134. [Google Scholar] [CrossRef]
- Tu, H.M.; Fan, M.W.; Ko, J.C. Different habitat types affect bird richness and evenness. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, W.; Seol, A.; Chung, O.-S.; Sagong, J.; Lee, J.K. Avian Reporting Rates in Chugcheongnam Province, South Korea Depend on Distance from Forest Edge, Size of Trees, and Size of Forest Fragments. Forests 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Gombobaatar, S.; Monks, E.M.; Seidler, R.; Sumiya, D.; Tseveenmyadag, N.; Bayarkhuu, S.; Baillie, J.E.; Boldbaatar, S.; Uuganbayar, C. Regional Red List Series Vol. 7. Birds; Zoological Society of London, National University of Mongolia, and Mongolian Ornithological Society: Ulaanbaatar, Mongolia, 2011; (In English and Mongolian). [Google Scholar]
- Bold, A. Mongolian Birds. In Mongolia Today Science, Culture, Environment and Development; Badarch, D., Zilinskas, R.A., Balint, P., Eds.; RoutledgeCurzon: London, UK, 2003; pp. 143–171. [Google Scholar]
- Bold, A. Mongolian Birds study of past 5 years. Sci. Proceed. Inst. Biol. Mong. Acad. Sci. 1973, 7, 143–171. (In Mongolian) [Google Scholar]
- Bold, A. Birds of Hentii mountain region. Sci. Proc. Inst. Biol. Mong. Acad. Sci. 1969, 3, 4–26. (In Mongolian) [Google Scholar]
- Bold, A. Birds of Hentii Mountain Region and Their Practical Importance. Ph.D. Dissertation, Department of Biology, National University of Mongolia, Ulaanbaatar, Mongolia, 1977. (In Russian). [Google Scholar]
- Bold, A. Census result of game birds in Hentii. Sci. Proc. Inst. Biol. Mong. Acad. Sci. 1970, 4, 19–27. (In Mongolian) [Google Scholar]
- Jambaajamts, A. Climate Brief Overview of the Republic of Mongolia; National Press Office: Ulaanbaatar, Mongolia, 1989. (In Mongolian) [Google Scholar]
- Tsegmid, S. Physical Geography of Mongolia; Mongolian Academy of Sciences, Institute of Geography and Permafrost, National publishing: Ulaanbaatar, Mongolia, 1969; p. 405. (In Mongolian) [Google Scholar]
- Sato, T.; Kimura, F.; Kitoh, A. Projection of global warming onto regional precipitation over Mongolia using a regional climate model. J. Hydrol. 2007, 333, 144–154. [Google Scholar] [CrossRef]
- Tsegmid, C. Some results of studies on microclimate and soil humidity of microassociations in mossy Larix forest of the eastern Khentey. Tesisi Docl Nauchnoi Konf. Posveshennie Vopr. Vozobnov. Resur. Lesa MNR 1989, 170–176. [Google Scholar]
- Ermakov, N.; Cherosov, M.; Gogoleva, P. Classification of ultracontinental boreal forests in central Yakutia. Folia Geobot. 2002, 37, 419–440. [Google Scholar] [CrossRef]
- Dulamsuren, C. Floristische Diversität, Vegetation und Standortbedingungen in der Gebirgstaiga des Westkhentej, Nordmongolei. Ber Forsch. Wald. A 2004, 191, 1–290. [Google Scholar]
- Mühlenberg, M.; Appelfelder, J.; Hoffmann, H.; Ayush, E.; Wilson, K.J. Structure of the montane taiga forests of West Khentii, Northern Mongolia. J. For. Sci. 2012, 58, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Hauck, M.; Mühlenberg, M. Vegetation at the taiga forest-steppe borderline in the western Khentej Mountains, northern Mongolia. Ann. Bot. Fenn. 2005, 42, 411–426. [Google Scholar]
- Bowman, J. Tujiin Nars: A Story of the Forest. Independent Study Project (ISP) Collection. 2012. Available online: https://digitalcollections.sit.edu/isp_collection/1453 (accessed on 20 June 2021).
- Bibby, C.J.; Burgess, N.D.; Hill, D.A. Bird Census Techniques; Academic Press: London, UK, 1992. [Google Scholar]
- Bibby, C.J.; Jones, M.; Marsden, S. Bird Surveys; Expedition Advisory Centre: London, UK, 1998. [Google Scholar]
- Hanni, D.J.; White, C.M.; van Lanen, N.J.; Birek, J.J.; Berven, J.M.; McLaren, M.F. Integrated Monitoring in Bird Conservation Regions (Imbcr): Field Protocol for Spatially-Balanced Sampling of Landbird Populations, Unpublished Report; Bird Conservancy of the Rockies: Brighton, CO, USA, 2016. [Google Scholar]
- Blakesley, J.A.; Hanni, D.J. Monitoring Colorado’s Birds, 2008. Technical Report M-MCB08-01; Rocky Mountain Bird Observatory: Brighton, CO, USA, 2009. [Google Scholar]
- QGIS. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: https://qgis.org/en/site/ (accessed on 20 June 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org/ (accessed on 15 September 2021).
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/package=vegan (accessed on 5 January 2022).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Stenzel, T.; Stubbe, M.; Samjaa, R.; Gombobaatar, S. Quantitative Investigations on Bird Communities in Different Habitats in the Orkhon-Selenge-Valley in Northern Mongolia; Erforschung biologischer Ressourcen der Mongolei/Exploration into the Biological Resources of Mongolia: Ulaanbaatar, Mongolia, 2005; p. 127. ISSN 0440-1298. [Google Scholar]
- Stokland, J.N. Representativeness and Efficiency of Bird and Insect Conservation in Norwegian Boreal Forest Reserves: Representatividad y Eficiencia en la Conservación de Aves e Insectos en las Reservas de Bosque Boreal de Noruega. Conserv. Biol. 1997, 11, 101–111. [Google Scholar] [CrossRef]
- Felton, A.; Andersson, E.; Ventorp, D.; Lindbladh, M. A comparison of avian diversity in spruce monocultures and spruce-birch polycultures in Southern Sweden. Silva Fenn. 2011, 45, 1143–1150. [Google Scholar] [CrossRef] [Green Version]
- Duco, R.A.; Fidelino, J.S.; Duya, M.V.; Ledesma, M.M.; Ong, P.S.; Duya, M.R. Bird Assemblage and Diversity along Different Habitat Types in a Karst Forest Area in Bulacan, Luzon Island, Philippines. Philipp. J. Sci. 2020, 15, 399–414. [Google Scholar]
- Berg, Å. Diversity and abundance of birds in relation to forest fragmentation, habitat quality and heterogeneity. Bird Study 1997, 44, 355–366. [Google Scholar] [CrossRef]
- Bersier, L.F.; Meyer, D.R. Bird assemblages in mosaic forests: The relative importance of vegetation structure and floristic composition along the successional gradient. Acta Oecologica 1994, 15, 561–576. [Google Scholar]
- Estades, C.F.; Temple, S.A. Deciduous-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecol. Appl. 1999, 9, 573–585. [Google Scholar] [CrossRef]
- Mansor, M.S.; Sah, S.A. The influence of habitat structure on bird species composition in lowland malaysian rain forests. Trop. Life Sci. Res. 2012, 23, 1–14. [Google Scholar] [PubMed]
- Deconchat, M.; Balent, G. Vegetation and bird community dynamics in fragmented coppice forests. Forestry 2001, 74, 105–118. [Google Scholar] [CrossRef]
- Haapanen, A. Bird fauna of the Finnish forests in relation to forest succession. In I. InAnnales Zoologici Fennici; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 1965; Volume 2, pp. 153–196. [Google Scholar]
- Bergner, A.; Avcı, M.; Eryiğit, H.; Jansson, N.; Niklasson, M.; Westerberg, L.; Milberg, P. Influences of forest type and habitat structure on bird assemblages of oak (Quercus spp.) and pine (Pinus spp.) stands in southwestern Turkey. For. Ecol. Manag. 2015, 336, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Nikolov, S.C. Effect of stand age on bird communities in late successional Macedonian pine forests in Bulgaria. For. Ecol. Manag. 2009, 257, 580–587. [Google Scholar] [CrossRef]
- Morozov, N.S. Breeding forest birds in the Valdai Uplands, north-west Russia: Assemblage composition, interspecific associations and habitat amplitudes. In InAnnales Zoologici Fennici; Finnish Zoological Publishing Board, formed by the Finnish Academy of Sciences, Societas Biologica Fennica Vanamo, Societas pro Fauna et Flora Fennica, and Societas Scientiarum Fennica: Helsinki, Finland, 1992; pp. 7–28. [Google Scholar]
- Enoksson, B.; Angelstam, P.; Larsson, K. Deciduous forest and resident birds: The problem of fragmentation within a coniferous forest landscape. Landsc. Ecol. 1995, 10, 267–275. [Google Scholar] [CrossRef]
- Laurance, S.G.; Stouffer, P.C.; Laurance, W.F. Effects of road clearings on movement patterns of understory rainforest birds in central Amazonia. Conserv. Biol. 2004, 18, 1099–1109. [Google Scholar] [CrossRef]
- Tvardíková, K. Bird abundances in primary and secondary growths in Papua New Guinea: A preliminary assessment. Trop. Conserv. Sci. 2010, 3, 373–388. [Google Scholar] [CrossRef]
- Şekercioḡlu, Ç.H.; Ehrlich, P.R.; Daily, G.C.; Aygen, D.; Goehring, D.; Sandí, R.F. Disappearance of insectivorous birds from tropical forest fragments. Proc. Natl. Acad. Sci. USA 2002, 99, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).