
Genetic Structure of Pinus Populations in the Urals

 , , , and
, , , and 
Abstract
:1. Introduction
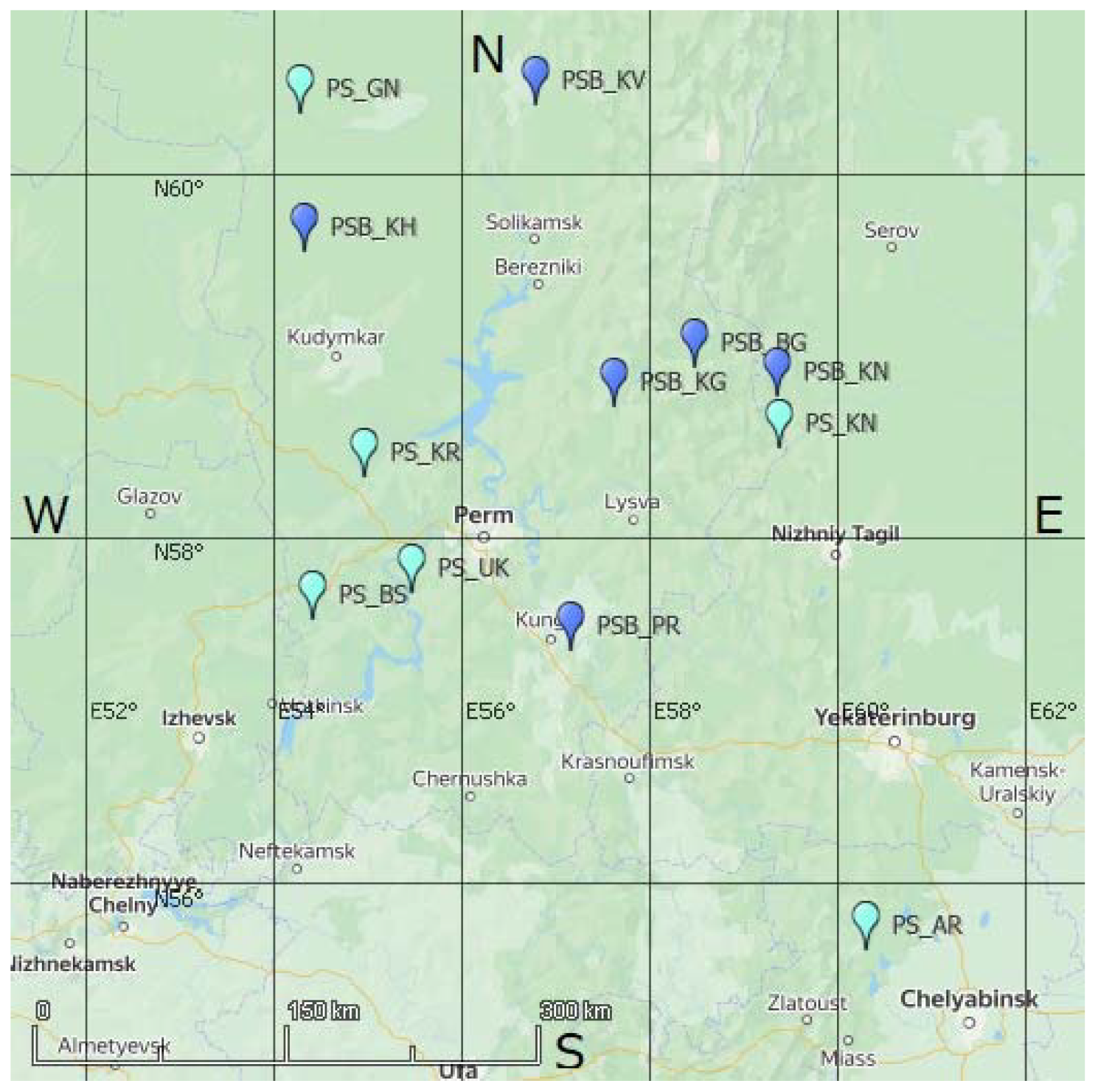
2. Materials and Methods
3. Results
3.1. Genetic Diversity of P. sylvestris
3.2. Genetic Diversity of P. sibirica
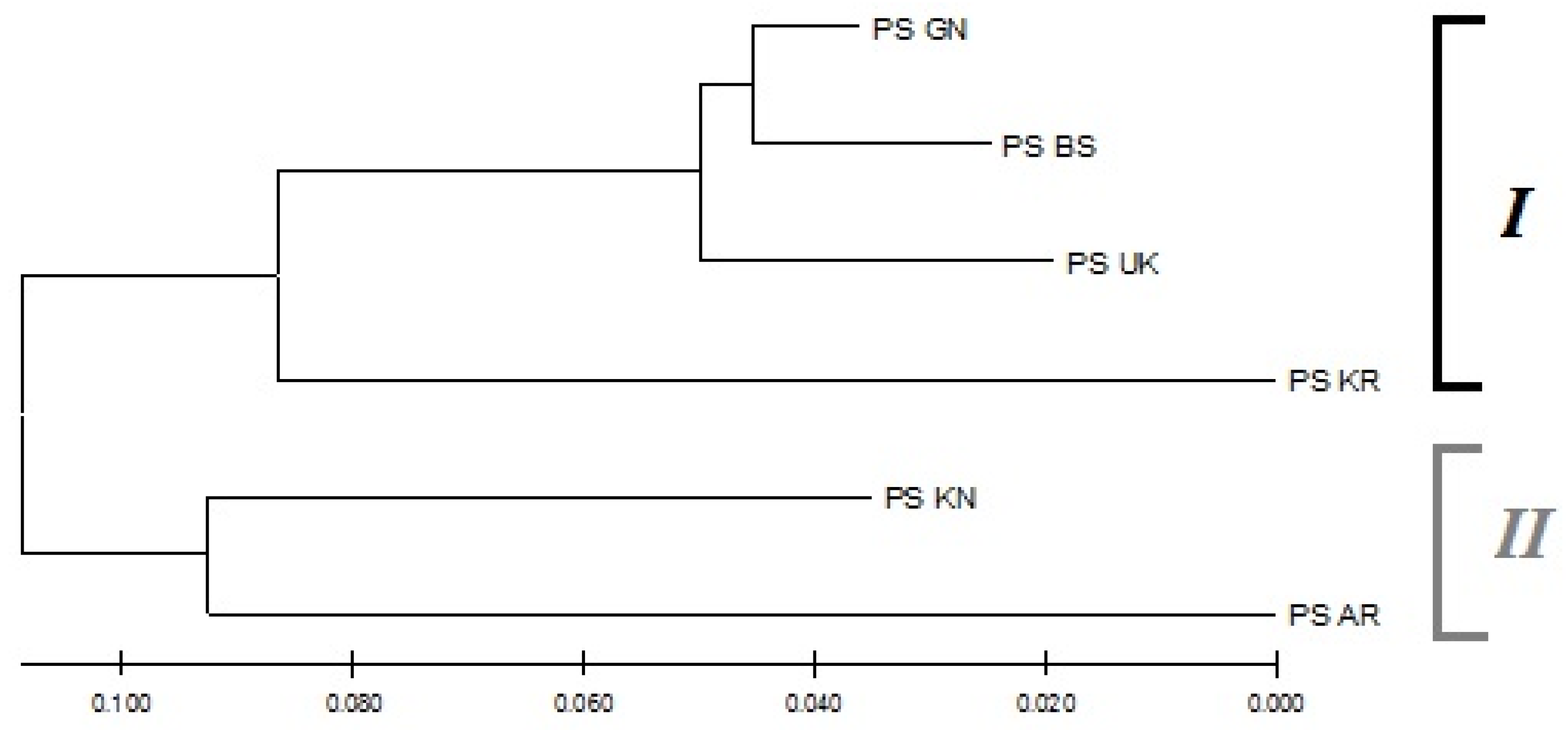
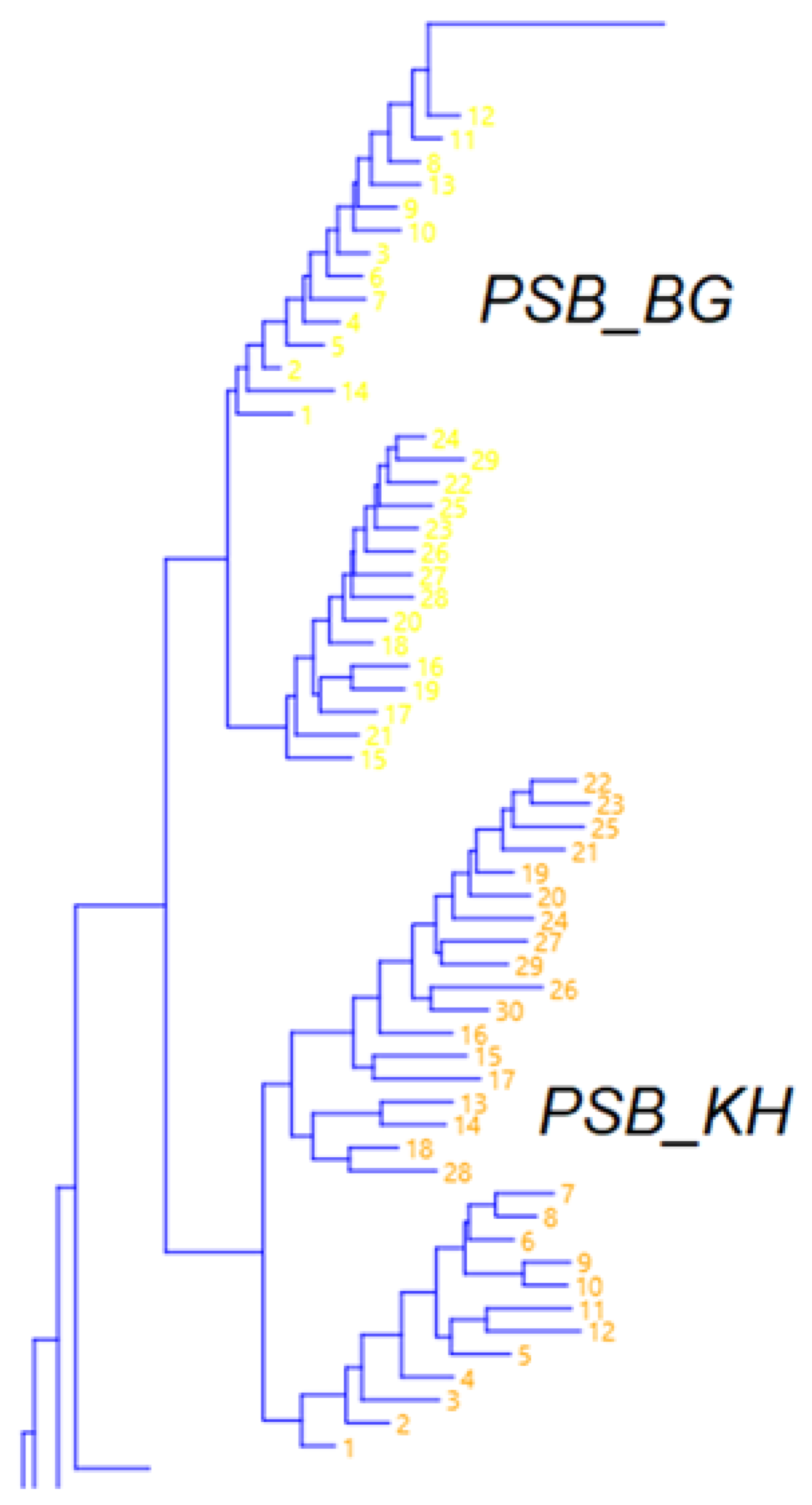
3.3. Population Genetic Structure of P. sylvestris
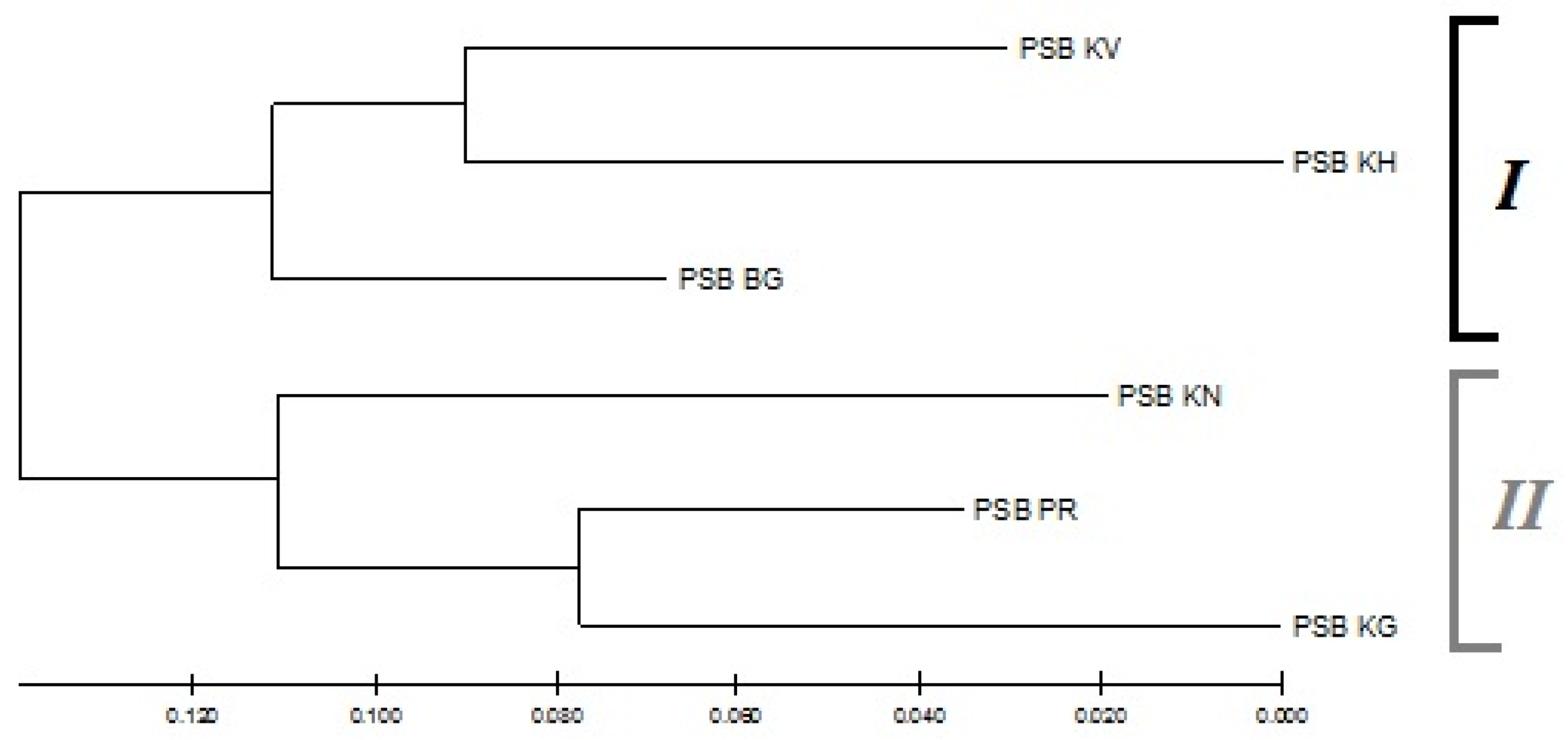
3.4. Population Genetic Structure of P. sibirica
4. Discussion
4.1. Genetic Diversity of P. sylvestris
4.2. Population Genetic Structure of P. sylvestris
4.3. Genetic Diversity of P. sibirica
4.4. Population Genetic Structure of P. sibirica
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. The Global Forest Resources Assessment 2020 (Fra 2020); FAO: Rome, Italy, 2020. [Google Scholar]
- Du, Z.; Yu, L.; Yang, J.; Xu, Y.; Chen, B.; Peng, S.; Zhang, T.; Fu, H.; Harris, N.; Gong, P. A global map of planting years of plantations. Sci. Data 2022, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Rocha, J.M.; Soares, L.; Andrade, E.R.; Gaiotto, F.A.; Cazetta, E. The impact of anthropogenic disturbances on the genetic diversity of terrestrial species: A global meta-analysis. Mol. Ecol. 2020, 29, 4812–4822. [Google Scholar] [CrossRef] [PubMed]
- Hogberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; Hogberg, M.N.; Nyberg, G.; Ottosson-Lofvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Lindén, A.; Heinonsalo, J.; Buchmann, N.; Oinonen, M.; Sonninen, E.; Hilasvuori, E.; Pumpanen, J. Contrasting effects of increased carbon input on boreal som decomposition with and without presence of living root system of Pinus sylvestris L. Plant Soil 2013, 377, 145–158. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Liu, X.; Chen, W.; Liu, Q.; Dai, J. Abietic acid suppresses non-small-cell lung cancer cell growth via blocking ikkbeta/nf-kappab signaling. OncoTargets Ther. 2019, 12, 4825–4837. [Google Scholar] [CrossRef]
- Dering, M.; Kosiński, P.; Wyka, T.P.; Pers-Kamczyc, E.; Boratyński, A.; Boratyńska, K.; Reich, P.B.; Romo, A.; Zadworny, M.; Żytkowiak, R.; et al. Tertiary remnants and holocene colonizers: Genetic structure and phylogeography of scots pine reveal higher genetic diversity in young boreal than in relict mediterranean populations and a dual colonization of fennoscandia. Divers. Distrib. 2017, 23, 540–555. [Google Scholar] [CrossRef]
- Ianbaev, R.; Boronnikova, S.; Yanbaev, Y.; Gainanov, S.; Kulagin, A. Population genetic diversity in quercus robur and ulmus laevis in southern urals (Russia): A comparatively study of adults and progeny in localities with contrast forest cover. Cent. Eur. For. J. 2022, 68, 101–108. [Google Scholar] [CrossRef]
- Makeeva, V.M.; Smurov, A.V.; Politov, D.V.; Belokon, M.M.; Belokon, Y.S.; Suslova, E.G. Comparative assessment of the gene pool and the viability of forest plantations from moscow and natural populations from the moscow region by example of norway spruce (Picea abies (L.) karst.). Russ. J. Genet. 2018, 54, 1015–1025. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Zanuso, E.; Genisheva, Z.; Rocha, C.M.R.; Teixeira, J.A. Green and sustainable valorization of bioactive phenolic compounds from pinus by-products. Molecules 2020, 25, 2931. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Petrova, I.V. Phylogenogeography and genotaxonomy of Pinus sylvestris L. Populations. Russ. J. Ecol. 2012, 43, 273–280. [Google Scholar] [CrossRef]
- Petrova, E.A.; Belokon, Y.S. Genetic polymorphism in siberian stone pine (Pinus sibirica Du Tour) populations from ural and altai-sayan mountains. Probl. Bot. South. Sib. Mong. 2020, 19, 081–086. [Google Scholar] [CrossRef]
- Milyutin, L.I. Study of siberian forest genetic resources. Sib. Lesn. Zurnal 2016, 3, 3–9. [Google Scholar] [CrossRef]
- Milyutin, L.I.; Muratova, E.N.; Larionova, A.Y. Development of forest genetics in russia. Sib. Lesn. Zurnal 2018, 1, 3–15. [Google Scholar] [CrossRef]
- Szmidt, A.E.; Wang, X.-R.; Lu, M.-Z. Empirical assessment of allozyme and rapd variation in Pinus sylvestris (L.) using haploid tissue analysis. Heredity 1996, 76, 412–420. [Google Scholar] [CrossRef]
- Shigapov, Z.K.; Putenikhin, K.V.; Shigapova, A.I.; Urazbakhtina, K.A.; Putenikhin, V.P. Genetic diversity of siberian stone pine under introduction in the south urals and bashkir cis-urals. Sib. Lesn. Zurnal 2016, 5, 137–146. [Google Scholar] [CrossRef]
- Tikhonova, I.V.; Ekart, A.K.; Zatsepina, K.G.; Kravchenko, A.N. Variability of allozyme locuss and inbreeding level in the age groups of southern-taiga and forest-steppe pine populations of the central siberia. Sib. Lesn. Zurnal 2019, 6, 70–80. [Google Scholar] [CrossRef]
- Semerikov, V.L.; Semerikova, S.A.; Dymshakova, O.S.; Zatsepina, K.G.; Tarakanov, V.V.; Tikhonova, I.V.; Ekart, A.K.; Vidyakin, A.I.; Jamiyansuren, S.; Rogovtsev, R.V.; et al. Microsatellite loci polymorphism of chloroplast DNA of scots pine (Pinus sylvestris L.) in asia and eastern europe. Russ. J. Genet. 2014, 50, 577–585. [Google Scholar] [CrossRef]
- Parfionava, N.S.; Paliavoi, S.A.; Spivak, A.A.; Hrebianchuk, A.E.; Hoh, A.N.; Kotava, S.A. Research of polymorphism of microsatellite markers of scots pine for the purposes of forensic DNA analysis. Exp. Biol. Biotechnol. 2022, 1, 90–98. [Google Scholar] [CrossRef]
- Tóth, E.G.; Vendramin, G.G.; Bagnoli, F.; Cseke, K.; Höhn, M. High genetic diversity and distinct origin of recently fragmented scots pine (Pinus sylvestris L.) populations along the carpathians and the pannonian basin. Tree Genet. Genomes 2017, 13, 47. [Google Scholar] [CrossRef]
- Tóth, E.G.; Köbölkuti, Z.A.; Pedryc, A.; Höhn, M. Evolutionary history and phylogeography of scots pine (Pinus sylvestris L.) in europe based on molecular markers. J. For. Res. 2017, 28, 637–651. [Google Scholar] [CrossRef]
- Kurt, W.; Hilde, N.; Markus, P.; Kirsten, W.; Günter, K. DNA Fingerprinting in Plants: Principles, Methods, and Applications, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2005; p. 472. [Google Scholar] [CrossRef]
- Oreshkova, N.V.; Sedel’nikova, T.S.; Pimenov, A.V.; Efremov, S.P. Analysis of genetic structure and differentiation of the bog and dry land populations of pinus sibirica du tour based on nuclear microsatellite loci. Russ. J. Genet. 2014, 50, 934–941. [Google Scholar] [CrossRef]
- Rubio-Moraga, A.; Candel-Perez, D.; Lucas-Borja, M.E.; Tiscar, P.A.; Vinegla, B.; Linares, J.C.; Gomez-Gomez, L.; Ahrazem, O. Genetic diversity of pinus nigra arn. Populations in southern spain and northern morocco revealed by inter-simple sequence repeat profiles. Int. J. Mol. Sci. 2012, 13, 5645–5658. [Google Scholar] [CrossRef]
- Labra, M.; Grassi, F.; Sgorbati, S.; Ferrari, C. Distribution of genetic variability in southern populations of scots pine (Pinus sylvestris L.) from the alps to the apennines. Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 468–476. [Google Scholar] [CrossRef]
- Vasilyeva, G.; Semerikov, V. Application of amplified fragment length polymorphisms markers to study the hybridization between pinus sibirica and p. Pumila. Ann. For. Res. 2014, 57, 175–180. [Google Scholar] [CrossRef]
- Vidyakin, A.I.; Boronnikova, S.V.; Nechayeva, Y.S.; Pryshnivskaya, Y.V.; Boboshina, I.V. Genetic variation, population structure, and differentiation in scots pine (Pinus sylvestris L.) from the northeast of the russian plain as inferred from the molecular genetic analysis data. Russ. J. Genet. 2015, 51, 1213–1220. [Google Scholar] [CrossRef]
- Chertov, N.; Vasilyeva, Y.; Zhulanov, A.; Nechaeva, Y.; Boronnikova, S.; Kalendar, R. Genetic structure and geographical differentiation of larix sibirica ledeb. In the urals. Forests 2021, 12, 1401. [Google Scholar] [CrossRef]
- Vasilyeva, Y.; Chertov, N.; Nechaeva, Y.; Sboeva, Y.; Pystogova, N.; Boronnikova, S.; Kalendar, R. Genetic structure, differentiation and originality of Pinus sylvestris L. Populations in the east of the east european plain. Forests 2021, 12, 999. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. Aflp: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (ssr)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Sivolap Iu, M.; Kalendar, R.N.; Chebotar, S.V. The genetic polymorphism of cereals demonstrated by pcr with random primers. TSitologiia I Genet. 1994, 28, 54–61. [Google Scholar]
- Voronova, A.; Rendon-Anaya, M.; Ingvarsson, P.; Kalendar, R.; Rungis, D. Comparative study of pine reference genomes reveals transposable element interconnected gene networks. Genes 2020, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Shuvaev, D.N.; Ibe, A.A. Genetic structure and postglacial recolonization of pinus sibirica du tour in the west siberian plain, inferred from nuclear microsatellite markers. Silvae Genet. 2021, 70, 99–107. [Google Scholar] [CrossRef]
- Kalendar, R.; Boronnikova, S.; Seppanen, M. Isolation and purification of DNA from complicated biological samples. Methods Mol. Biol. 2021, 2222, 57–67. [Google Scholar] [CrossRef]
- Kalendar, R.; Schulman, A.H. Irap and remap for retrotransposon-based genotyping and fingerprinting. Nat. Protoc. 2006, 1, 2478–2484. [Google Scholar] [CrossRef]
- Kalendar, R. A guide to using fastpcr software for pcr, in silico pcr, and oligonucleotide analysis. Methods Mol. Biol. 2022, 2392, 223–243. [Google Scholar] [CrossRef]
- Sboeva, Y.; Vasil’eva, Y.; Chertov, N.; Pystogova, N.; Boronnikova, S.; Kalendar, R.; Martynenko, N. Molecular genetic identification of scots pine and siberian larch populations in perm krai based on polymorphism of issr-pcr markers. Sib. Lesn. Zurnal 2020, 4, 35–44. [Google Scholar] [CrossRef]
- Nechaeva, Y.; Pystogova, N.; Chertov, N.; Boronnikova, S. Molecular genetic analysis of Pinus sylvestris L. And pinus sibirica du tour populations in perm krai based on polymorphism issr-pcr markers. Bull. Sci. Pract. 2021, 7, 12–21. [Google Scholar] [CrossRef]
- Kalendar, R.; Schulman, A.H. Transposon-based tagging: Irap, remap, and ipbs. Methods Mol. Biol. 2014, 1115, 233–255. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. Genalex 6: Genetic analysis in excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T.B.J.; Ye, Z.H.; Mao, J.X. Popgene, the User Friendly Shareware for Population Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1997; Volume 10. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- McInnes, L.; Healy, J.; Saul, N.; Großberger, L. Umap: Uniform manifold approximation and projection. J. Open Source Softw. 2018, 3, 861. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. Scipy 1.0: Fundamental algorithms for scientific computing in python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Khanova, E.; Konovalov, V.; Timeryanov, A.; Isyanyulova, R.; Rafikova, D. Genetic and selection assessment of the scots pine (Pinus sylvestris L.) in forest seed orchards. Wood Res. 2020, 65, 283–292. [Google Scholar] [CrossRef]
- Yanbaev, Y.; Sultanova, R.; Blonskaya, L.; Bakhtina, S.; Tagirova, A.; Tagirov, V.; Kulagin, A. Gene pool of scots pine (Pinus sylvestris L.) under reforestation in extreme environment. Wood Res. 2020, 65, 459–470. [Google Scholar] [CrossRef]
- Șofletea, N.; Mihai, G.; Ciocîrlan, E.; Curtu, A.L. Genetic diversity and spatial genetic structure in isolated scots pine (Pinus sylvestris L.) populations native to eastern and southern carpathians. Forests 2020, 11, 1047. [Google Scholar] [CrossRef]
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. New Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Petrova, I.V.; Egorov, E.V.; Sannikova, N.S. A system of pleistocene refugia for Pinus sylvestris L. In the southern marginal part of the species range. Russ. J. Ecol. 2014, 45, 167–173. [Google Scholar] [CrossRef]
- Petrova, Y.A.; Velisevich, S.N.; Belokon, M.M.; Belokon, Y.S.; Politov, D.V.; Goroshkevich, S.N. Genetic diversity and differentiation of siberian stone pine populations at the southern edge in lowland part of west siberia. Ecol. Genet. 2014, 12, 48–61. [Google Scholar] [CrossRef]
- Yang, C.; Wei, L.; Jiang, J.; Liu, G.; Zhao, G. Analysis of genetic diversity for nineteen populations of pinus sibirica du tour with technique of issr. J. Northeast. For. Univ. 2005, 33, 1–3. [Google Scholar]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors Influencing Levels of Genetic Diversity in Woody Plant Species; Springer: Dordrecht, The Netherlands, 1992; pp. 95–124. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Sequence (5′–3′) | Tm (°C) | Ta (°C) * | Total Bands | PIC * |
|---|---|---|---|---|---|
| ISSR-1(AC)8T | ACACACACACACACACT | 59.0 | 56 | 15 | 0.315 |
| CR-212(CT)8TG | CTCTCTCTCTCTCTCTTG | 55.1 | 56 | 14 | 0.316 |
| CR-215(CA)6GT | CACACACACACAGT | 52.0 | 56 | 24 | 0.331 |
| M27(GA)8C | GAGAGAGAGAGAGAGAC | 54.3 | 52 | 17 | 0.251 |
| X10(AGC)6C | AGCAGCAGCAGCAGCAGCC | 72.5 | 64 | 19 | 0.300 |
| X11(AGC)6G | AGCAGCAGCAGCAGCAGCG | 72.5 | 64 | 30 | 0.309 |
| CR-217(GT)6GG | GTGTGTGTGTGTGG | 53.8 | 52 | 21 | 0.331 |
| ISSR-9(ACG)7G | ACGACGACGACGACGACGACGG | 73.7 | 64 | 29 | 0.305 |
| M1(AC)8CG | ACACACACACACACACCG | 63.6 | 60 | 22 | 0.369 |
| Populations | He | ne | I | R |
|---|---|---|---|---|
| PS_GN | 0.185 | 1.312 | 0.280 | 0 |
| (0.021) | (0.040) | (0.030) | ||
| PS_KN | 0.149 | 1.244 | 0.229 | 3 |
| (0.020) | (0.036) | (0.029) | ||
| PS_KG | 0.163 | 1.269 | 0.248 | 4 |
| (0.020) | (0.037) | (0.030) | ||
| PS_BS | 0.152 | 1.244 | 0.235 | 1 |
| (0.019) | (0.034) | (0.028) | ||
| PS_UK | 0.150 | 1.246 | 0.231 | 0 |
| (0.020) | (0.035) | (0.029) | ||
| PS_AR | 0.180 | 1.305 | 0.272 | 1 |
| (0.021) | (0.039) | (0.031) | ||
| Total | 0.163 | 1.270 | 0.249 | 9 |
| (0.008) | (0.015) | (0.012) |
| Populations | He | ne | I | R |
|---|---|---|---|---|
| PSB_KV | 0.115 | 1.195 | 0.174 | 1 |
| (0.016) | (0.029) | (0.023) | ||
| PSB_KH | 0.200 | 1.346 | 0.298 | 4 |
| (0.018) | (0.034) | (0.026) | ||
| PSB_BG | 0.159 | 1.273 | 0.238 | 1 |
| (0.018) | (0.032) | (0.025) | ||
| PSB_PR | 0.127 | 1.214 | 0.192 | 0 |
| (0.016) | (0.030) | (0.024) | ||
| PSB_KG | 0.125 | 1.221 | 0.186 | 1 |
| (0.017) | (0.032) | (0.025) | ||
| PSB_KN | 0.160 | 1.238 | 0.262 | 8 |
| (0.014) | (0.025) | (0.021) | ||
| Total | 0.148 | 1.248 | 0.225 | 15 |
| (0.007) | (0.013) | (0.010) |
| PS_GN | PS_KN | PS_KR | PS_BS | PS_UK | |
|---|---|---|---|---|---|
| 0.402 | - | 0.001 | 0.001 | 0.001 | PS_KN |
| 0.405 | 0.502 | - | 0.001 | 0.001 | PS_KR |
| 0.137 | 0.433 | 0.425 | - | 0.001 | PS_BS |
| 0.233 | 0.449 | 0.477 | 0.242 | - | PS_UK |
| 0.428 | 0.370 | 0.462 | 0.464 | 0.461 | PS_AR |
| Subdivision Index | df | SS | MS | Variance | % | p |
|---|---|---|---|---|---|---|
| Between populations | 5 | 873,646 | 174,729 | 5710 | 41% | <0.001 |
| Within populations | 169 | 1,410,822 | 8348 | 8348 | 59% | <0.001 |
| PS_GN | PS_KN | PS_KR | PS_BS | PS_UK | |
|---|---|---|---|---|---|
| 0.147 | PS_KN | ||||
| 0.136 | 0.201 | PS_KR | |||
| 0.035 | 0.148 | 0.136 | PS_BS | ||
| 0.040 | 0.151 | 0.161 | 0.060 | PS_UK | |
| 0.194 | 0.147 | 0.205 | 0.195 | 0.199 | PS_AR |
| PSB_KV | PSB_KH | PSB_BG | PSB_PR | PSB_KG | |
|---|---|---|---|---|---|
| 0.424 | - | 0.001 | 0.001 | 0.001 | PSB_KH |
| 0.424 | 0.437 | - | 0.001 | 0.001 | PSB_BG |
| 0.566 | 0.517 | 0.515 | - | 0.001 | PSB_PR |
| 0.629 | 0.565 | 0.542 | 0.424 | - | PSB_KG |
| 0.493 | 0.486 | 0.477 | 0.397 | 0.446 | PSB_KN |
| Subdivision Index | df | SS | MS | Variance | % | p |
|---|---|---|---|---|---|---|
| Between populations | 5 | 1,389,452 | 277,890 | 11,078 | 49% | <0.001 |
| Within populations | 140 | 1,598,370 | 11,417 | 11,417 | 51% | <0.001 |
| PSB_KV | PSB_KH | PSB_BG | PSB_PR | PSB_KG | |
|---|---|---|---|---|---|
| 0.145 | PSB_KH | ||||
| 0.121 | 0.160 | PSB_BG | |||
| 0.214 | 0.220 | 0.198 | PSB_PR | ||
| 0.271 | 0.285 | 0.192 | 0.125 | PSB_KG | |
| 0.227 | 0.256 | 0.192 | 0.167 | 0.195 | PSB_KN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chertov, N.; Nechaeva, Y.; Zhulanov, A.; Pystogova, N.; Danilova, M.; Boronnikova, S.; Kalendar, R. Genetic Structure of Pinus Populations in the Urals. Forests 2022, 13, 1278. https://doi.org/10.3390/f13081278
Chertov N, Nechaeva Y, Zhulanov A, Pystogova N, Danilova M, Boronnikova S, Kalendar R. Genetic Structure of Pinus Populations in the Urals. Forests. 2022; 13(8):1278. https://doi.org/10.3390/f13081278
Chicago/Turabian StyleChertov, Nikita, Yulia Nechaeva, Andrei Zhulanov, Nina Pystogova, Maria Danilova, Svetlana Boronnikova, and Ruslan Kalendar. 2022. "Genetic Structure of Pinus Populations in the Urals" Forests 13, no. 8: 1278. https://doi.org/10.3390/f13081278
APA StyleChertov, N., Nechaeva, Y., Zhulanov, A., Pystogova, N., Danilova, M., Boronnikova, S., & Kalendar, R. (2022). Genetic Structure of Pinus Populations in the Urals. Forests, 13(8), 1278. https://doi.org/10.3390/f13081278