
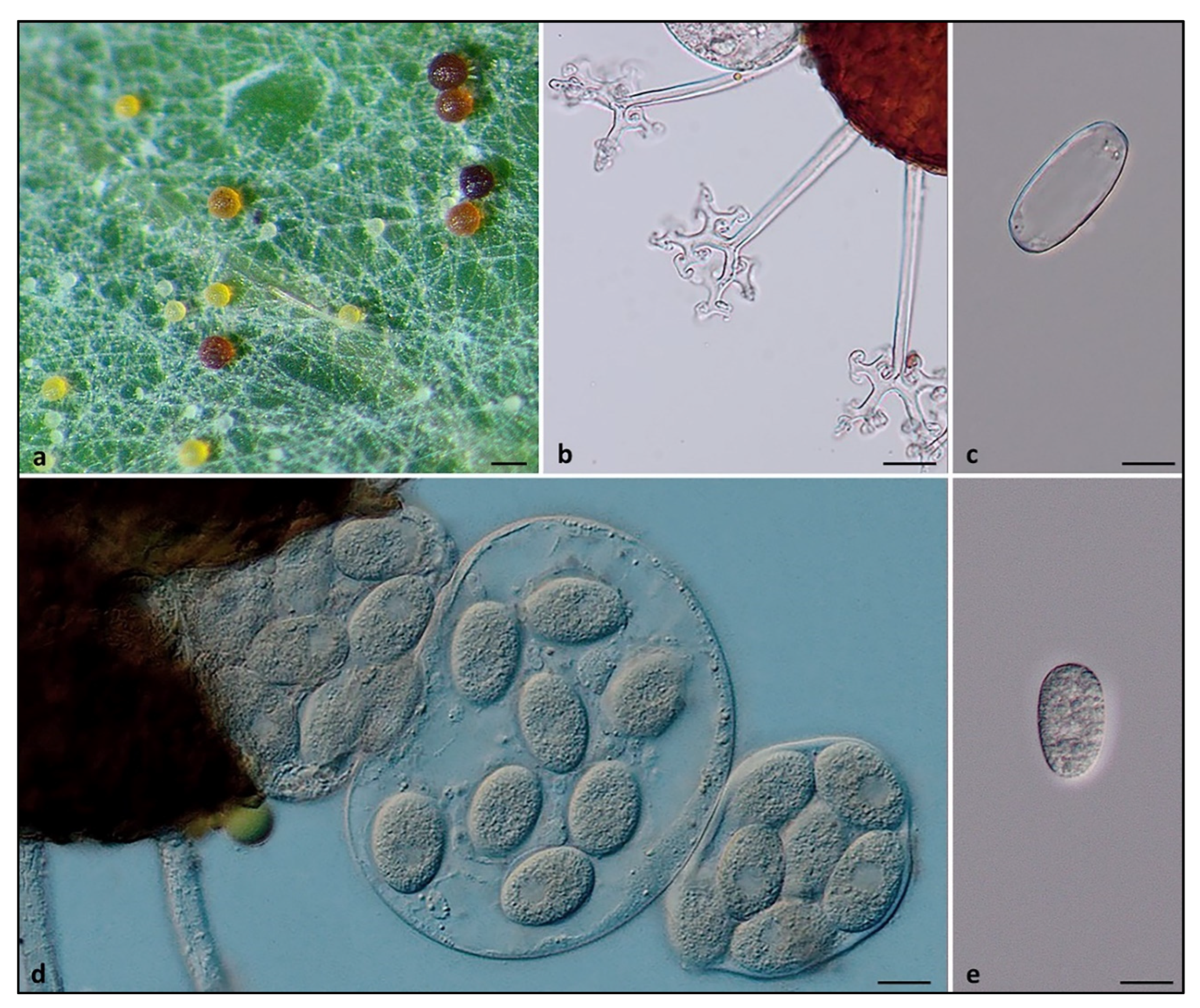
Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L.
 ,
,  , , ,
, , , 
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Those at the physiological level (net rate of photosynthesis, stomatal conductance, and transpiration);
- The differential pathogen or ectomycorrhiza-specific accumulation of osmotically active substances such as glycine-betaine and proline;
- The differential regulation of polyamine metabolism resulting in different foliar polyamine profiles;
- The differential activation of the total antioxidant (ABTS) and reducing (FRAP) capacity and accumulation of various antioxidants (i.e., non-protein thiols, phenolics, flavonoids, and tannins).
2. Materials and Methods
2.1. Experimental Design
- ECM-PM—Oak plants infected with powdery mildew, whose roots were previously primed with ectomycorrhiza;
- NM-PM—Oak plants infected with powdery mildew with no mycorrhiza on their root tips;
- ECM-C—Oak plants that were not infected by powdery mildew but were primed and inoculated with ectomycorrhizal inoculum;
- NM-C—Healthy plants that were neither infected with powdery mildew nor inoculated by ectomycorrhizal inoculum.
2.2. Physiological Measurement
2.3. Measurements of Osmolytes’ Accumulation
- Polyamine profile and content of three main polyamines in plants—putrescine (PUT), spermidine (SPD), and spermine (SPM)—were determined in oak leaves via HPLC method after derivatization with dansyl-chloride as a pre-treatment [54]. Plant tissues (approx. 20 mg DW of freeze-dried material) were extracted with 10 volumes of 4% perchloric acid (PCA); the homogenate was kept on ice for 1 h and then centrifuged at 15,000× g for 30 min. Aliquots of the supernatants and standard solutions of dansylated derivatives were extracted with toluene, dried, and resuspended in acetonitrile prior to HPLC analysis. PAs were separated and quantified by HPLC (Şchimadzu, Kyoto, Japan) using a reverse phase C18 column (Spherisorb ODS2, 5-μm particle diameter, 4.6 × 250 mm, Waters, Wexford, Ireland) and a programmed acetonitrile-water step gradient, as previously described [54].
- Proline (PRO) concentration was estimated following the well-established ninhydrin method [55]. Data were calculated on a DW basis.
- Glycine-betaine (GB), as predominant quaternary ammonium compound (QAC), was quantified using the precipitation method of QAC-periodide complexes in acid medium [56]. Data were calculated on a DW basis.
2.4. Assays of Antioxidant Defense Systems
- Lipid peroxidation was quantified by using a thiobarbituric assay where the level of lipid peroxidation corresponds to the amount of accumulated end-product malondialdehyde (MDA). Results are expressed as nmol MDA equivalents on a DW basis.
- Total non-protein thiol compounds were measured according to a modified colorimetric assay based on measuring the absorbance of yellow Ellman’s reagent (5,5′-dithiobis-(2-nitrobenyoic acid; DTNB) reduced by sulfhydryl compounds at 413 nm. After construction of the calibration curve, where we used reduced glutathione (GSH) as standard, total non-protein thiol compounds were expressed as GSH equivalents on a DW basis [57].
- Trolox® Equivalent Antioxidant Capacity (TEAC) was estimated with the 2,2′-azinobis-(3-ethylbenzothiozoline-6-sulfonic acid) (ABTS) assay based on the capability of the ethanolic extract to scavenge ABTS radicals [58]. Data were calculated on a DW basis.
- The Ferric Reducing Antioxidant Power (FRAP) assay provided additional measurement of the antioxidant activity level of ethanolic extracts. In this assay, the ability of the plant extract to reduce the ferric 2, 4, 6-tripyridyl-S-triazine complex [Fe3+—(TPTZ)2]3− to the intensely blue-colored ferrous complex [Fe3+—(TPTZ)2]2− in acidic medium was estimated [59]. Data are expressed as TEAC on a DW basis.
- Total phenolic content (TPC) was determined according to reactivity with Folin–Ciocalteu reagent [60]. Data are expressed as mg of Gallic Acid Equivalents (GAE) on a DW basis.
- Total flavonoid content (TFC) was measured by the aluminum chloride colorimetric method [61]. Data are expressed as mg of Quercetin Equivalent (QE) on a DW basis.
- Condensed tannins (CT) content was determined from methanolic extracts using butanol-HCl-Fe (III) method [62]. Data were expressed as leucocyanidin equivalents (LE) on a DW basis.
2.5. Elemental Analysis of Nitrogen and Carbon Content
2.6. Statistical Analysis
3. Results
Principal Component Analysis (PCA) and Correlation Matrix
4. Discussion
4.1. Effects of Ectomycorrhiza and/or Powdery Mildew on Oaks’ Nitrogen, Carbon, and Leaf Water Content
4.2. Perturbations in the Gas Exchange Parameters Caused by Powdery Mildew Infection and Inoculation with ECM Fungi
4.3. Erysiphe Alphioides and Ectomycorrhizal Inoculum Alter Amounts of Compatible Solutes
4.4. Oaks’ Foliar Phenolics Content Is Modulated by Ectomycorrhiza and/or Powdery Mildew
4.5. Reinforcement of Polyamine Metabolism Induced by Ectomycorrhizal Fungi and E. alphitodes
4.6. Limitation of the Study and Future Perspective
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Minx, J.C.; Farahani, E.; Kadner, S.; Seyboth, K.; Adler, A.; Baum, I.; Brunner, S.; et al. (Eds.) Climate Change 2014 Mitigation of Climate Change Working Group III Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Monahan, W.B.; Fisichelli, N.A. Climate Exposure of US National Parks in a New Era of Change. PLoS ONE 2014, 9, e101302. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, D.; Matović, B.; Orlović, S.; Kržič, A.; Trudić, B.; Galić, Z.; Stojnić, S.; Pekeč, S. Future of the Main Important Forest Tree Species in Serbia from the Climate Change Perspective. South-East Eur. For. 2014, 5, 117–124. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Pap, P.; Ranković, B.; Maširević, S. Significance and Need of Powdery Mildew Control (Microsphaera alphitoides Griff. et Maubl.) in the Process of Regeneration of the Pedunculate Oak (Quercus robur L.) Stands in the Ravni Srem Area. Period Biol. 2012, 114, 91–102. [Google Scholar]
- Marçais, B.; Desprez-Loustau, M.L. European Oak Powdery Mildew: Impact on Trees, Effects of Environmental Factors, and Potential Effects of Climate Change. Ann. For. Sci. 2014, 71, 633–642. [Google Scholar] [CrossRef]
- Keča, N.; Koufakis, I.; Dietershagen, J.; Nowakowska, J.A.; Oszako, T. European Oak Decline Phenomenon in Relationto Climatic Changes. Folia For. Pol. Ser. A 2016, 58, 170–177. [Google Scholar] [CrossRef]
- Bojović, M.; Nikolić, N.; Borišev, M.; Pajević, S.; Horák, R.; Pavlović, L.; Vaštag, E. The Effect of Drought Stress and Recovery on Pedunculate Oak Populations Grown in Semi-Controlled Conditions. Topola 2017, 199–200, 193–207. [Google Scholar]
- Desprez-Loustau, M.-L.; Massot, M.; Toïgo, M.; Fort, T.; Gülden, A.; Kaya, A.; Boberg, J.; Braun, U.; Capdevielle, X.; Cech, T.; et al. From Leaf to Continent: The Multi-Scale Distribution of an Invasive Cryptic Pathogen Complex on Oak. Fungal Ecol. 2018, 36, 39–50. [Google Scholar] [CrossRef]
- Essling, M.; McKay, S.; Petrie, P.R. Fungicide Programs Used to Manage Powdery Mildew (Erysiphe necator) in Australian Vineyards. Crop Prot. 2021, 139, 105369. [Google Scholar] [CrossRef]
- Berrie, A.; Xu, X. Developing Biopesticide-Based Programmes for Managing Powdery Mildew in Protected Strawberries in the UK. Crop Prot. 2021, 149, 105766. [Google Scholar] [CrossRef]
- Oszako, T.; Voitka, D.; Stocki, M.; Stocka, N.; Nowakowska, J.A.; Linkiewicz, A.; Hsiang, T.; Belbahri, L.; Berezovska, D.; Malewski, T. Trichoderma asperellum Efficiently Protects Quercus Robur Leaves against Erysiphe alphitoides. Eur. J. Plant Pathol. 2021, 159, 295–308. [Google Scholar] [CrossRef]
- Milović, M.; Kebert, M.; Orlović, S. How mycorrhizas can help forests to cope with ongoing climate change? Šumarski List 2021, 145, 279–286. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.M.; Ton, J. Mycorrhiza-Induced Resistance: More than the Sum of Its Parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Rabara, R.C.; Negi, S. AMF: The future prospect for sustainable agriculture. Physiol. Mol. Plant Pathol. 2018, 102, 36–45. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, A.; Shukla, A.; Kumar, S.; Uthappa, A.R.; Chaturvedi, O.P. Effect of arbuscular mycorrhiza fungi (AMF) on early seedling growth of some multipurpose tree species. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6–7, 3885–3892. [Google Scholar] [CrossRef]
- Simard, S.; Austin, M. The role of mycorrhizas in forest soil stability with climate change. In Climate Change and Variability; IntechOpen: London, UK, 2010. [Google Scholar]
- Keymer, A.; Gutjahr, C. Cross-Kingdom Lipid Transfer in Arbuscular Mycorrhiza Symbiosis and Beyond. Curr. Opin. Plant Biol. 2018, 44, 137–144. [Google Scholar] [CrossRef]
- Johny, L.; Cahill, D.M.; Adholeya, A. AMF Enhance Secondary Metabolite Production in Ashwagandha, Licorice, and Marigold in a Fungi-Host Specific Manner. Rhizosphere 2021, 17, 100314. [Google Scholar] [CrossRef]
- Ghanbarzadeh, Z.; Zamani, H.; Mohsenzadeh, S.; Marczak, Ł.; Stobiecki, M.; Zarei, M. Rhizosphere Symbionts Improve Water Stress Tolerance in Moldavian Balm through Modulation of Osmolytes. Rhizosphere 2021, 19, 100367. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, B. Arbuscular Mycorrhiza Induced Putrescine Degradation into γ-Aminobutyric Acid, Malic Acid Accumulation, and Improvement of Nitrogen Assimilation in Roots of Water-Stressed Maize Plants. Mycorrhiza 2020, 30, 329–339. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef]
- Romero, F.M.; Maiale, S.J.; Rossi, F.R.; Marina, M.; Ruíz, O.A.; Gárriz, A. Polyamine Metabolism Responses to Biotic and Abiotic Stress. Methods Mol. Biol. 2018, 1694, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Groppa, M.D.; Benavides, M.P. Polyamines and Abiotic Stress: Recent Advances Review Article. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, Z. Effects of Endogenous Abscisic Acid, Jasmonic Acid, Polyamines, and Polyamine Oxidase Activity in Tomato Seedlings under Drought Stress. Sci. Hortic. 2013, 159, 172–177. [Google Scholar] [CrossRef]
- Kebert, M.; Rapparini, F.; Neri, L.; Bertazza, B.; Orlović, S.; Biondi, S. Copper-Induced Responses in Poplar Clones Are Associated with Genotype- and Organ-Specific Changes in Peroxidase Activity and Proline, Polyamine, ABA, and IAA Levels. J. Plant Growth Regul. 2017, 36, 131–147. [Google Scholar] [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines Function in Stress Tolerance: From Synthesis to Regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef]
- Gerlin, L.; Baroukh, C.; Genin, S. Polyamines: Double Agents in Disease and Plant Immunity. Trends Plant Sci. 2021, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Gautam, A.; Dubey, A.K. Polyamines Metabolism and NO Signaling in Plants. In Nitric Oxide in Plant Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 345–372. [Google Scholar] [CrossRef]
- Bellin, D.; Asai, S.; Delledonne, M.; Yoshioka, H. Nitric Oxide as a Mediator for Defense Responses. Mol. Plant-Microbe Interact. 2013, 26, 271–277. Mol. Plant-Microbe Interact. 2013, 26, 271–277. [Google Scholar] [CrossRef]
- Jiménez-Bremont, J.F.; Marina, M.; de la Luz Guerrero-González, M.; Rossi, F.R.; Sánchez-Rangel, D.; Rodríguez-Kessler, M.; Ruiz, O.A.; Gárriz, A. Physiological and Molecular Implications of Plant Polyamine Metabolism during Biotic Interactions. Front. Plant Sci. 2014, 5, 95. [Google Scholar] [CrossRef]
- Sannazzaro, A.I.; Álvarez, C.L.; Menéndez, A.B.; Pieckenstain, F.L.; Albertó, E.O.; Ruiz, O.A. Ornithine and Arginine Decarboxylase Activities and Effect of Some Polyamine Biosynthesis Inhibitors on Gigaspora rosea Germinating Spores. FEMS Microbiol. Lett. 2004, 230, 115–121. [Google Scholar] [CrossRef]
- Wu, Q.S.; He, X.H.; Zou, Y.N.; Liu, C.Y.; Xiao, J.; Li, Y. Arbuscular Mycorrhizas Alter Root System Architecture of Citrus Tangerine through Regulating Metabolism of Endogenous Polyamines. Plant Growth Regul. 2012, 68, 27–35. [Google Scholar] [CrossRef]
- Shamshiri, M.H.; Fattahi, M. Evaluation of two biochemical markers for salt stress in three pistachio rootstocks inoculated with arbuscular mycorrhiza (Glomus mosseae). J. Stress Physiol. Biochem. 2014, 10, 335–346. [Google Scholar]
- Chen, T.H.H.; Murata, N. Glycinebetaine Protects Plants against Abiotic Stress: Mechanisms and Biotechnological Applications. Plant Cell Environ. 2011, 34, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Becker, D.F. Connecting Proline Metabolism and Signaling Pathways in Plant Senescence. Front. Plant Sci. 2015, 6, 552. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Tausz, M.; Šircelj, H.; Grill, D. The Glutathione System as a Stress Marker in Plant Ecophysiology: Is a Stress-Response Concept Valid? J. Exp. Bot. 2004, 55, 1955–1962. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Islam Anee, T.; Fujita, M. Glutathione in Plants: Biosynthesis and Physiological Role in Environmental Stress Tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Popović, B.M.; Štajner, D.; Ždero, R.; Orlović, S.; Galić, Z. Antioxidant Characterization of Oak Extracts Combining Spectrophotometric Assays and Chemometrics. Sci. World J. 2013, 2013, 134656. [Google Scholar] [CrossRef]
- Anttila, A.K.; Pirttilä, A.M.; Häggman, H.; Harju, A.; Venäläinen, M.; Haapala, A.; Holmbom, B.; Julkunen-Tiitto, R. Condensed Conifer Tannins as Antifungal Agents in Liquid Culture. Holzforschung 2013, 67, 825–832. [Google Scholar] [CrossRef]
- Sharma, K.P. Tannin Degradation by Phytopathogen’s Tannase: A Plant’s Defense Perspective. Biocatal. Agric. Biotechnol. 2019, 21, 101342. [Google Scholar] [CrossRef]
- Repáč, I.; Balanda, M.; Vencurik, J.; Kmeť, J.; Krajmerová, D.; Paule, L. Effects of Substrate and Ectomycorrhizal Inoculation on the Development of Two-Years-Old Container-Grown Norway Spruce (Picea abies Karst.) Seedlings. Iforest-Biogeosci. For. 2014, 8, 487. [Google Scholar] [CrossRef]
- Kowalski, S.; Chomicz, E.; Czeremuga, M.; Piecha, M. Selected Hebeloma species in ectomycorrhizal synthesis with Scots pine [Pinus sylvestris L.] and pedunculate oak [Quercus robur L.] in container nursery. Acta Agrar. Et Silvestria. Ser. Silvestris 2007, 45, 3–25. [Google Scholar]
- Olchowik, J.; Mariusz Bzdyk, R.; Studnicki, M.; Bederska-Błaszczyk, M.; Urban, A.; Aleksandrowicz-Trzcí Nska, M. The Effect of Silver and Copper Nanoparticles on the Condition of English Oak (Quercus robur L.) Seedlings in a Container Nursery Experiment. Forests 2017, 8, 310. [Google Scholar] [CrossRef]
- Milović, M.; Kovačević, B.; Pekeč, S.; Pilipović, A.; Kesić, L.; Orlović, S.; Gavranović Markić, A. Diversity of Ectomycorrhizal Fungi in Young Pedunculate Oak Stand from Morović, Serbia. South-East Eur. For. 2022, 13, 19–25. [Google Scholar] [CrossRef]
- Agerer, R. Exploration Types of Ectomycorrhizae. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Montecchio, L.; Mutto Accordi, S.; Rossi, S.; Causin, R. Changes in Ectomycorrhizal Diversity in a Declining ”Quercus ilex” Coastal Forest. In Phytopathologia Mediterranea; Firenze University Press: Florence, Italy, 2004; pp. 1000–1009. [Google Scholar]
- Karličić, V.; Jovičić-Petrović, J.; Marojević, V.; Zlatković, M.; Orlović, S.; Raičević, V. Potential of Trichoderma spp. and Pinus sylvestris Bark Extracts as Biocontrol Agents against Fungal Pathogens Residing in the Botryosphaeriales. Environ. Sci. Proc. 2020, 2021, 99. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Marković, M.; Orlović, S. Bacterial Canker Disease on Populus × Euramericana Caused by Lonsdalea Populi in Serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- McKinney, H.H. Investigations of the rosette disease of of Wheat and Its Control. J. Agric. Res. 1923, 23, 771. [Google Scholar]
- Bombelli, A.; Gratani, L. Interspecific differences of leaf gas exchange and water relations of three evergreen Mediterranean shrub species. Photosynthetica 2003, 41, 619–625. [Google Scholar] [CrossRef]
- Scaramagli, S.; Biondi, S.; Capitani, F.; Gerola, P.; Altamura, M.M.; Torrigiani, P. Polyamine Conjugate Levels and Ethylene Biosynthesis: Inverse Relationship with Vegetative Bud Formation in Tobacco Thin Layers. Physiol. Plant 1999, 105, 366–375. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid Assay for Determination of Water Soluble Quaternary Ammonium Compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- De Kok, L.J.; Buwalda, F.; Bosma, W. Determination of Cysteine and Its Accumulation in Spinach Leaf Tissue upon Exposure to Excess Sulfur. J. Plant Physiol. 1988, 133, 502–505. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors Influencing the Antioxidant Activity Determined by the ABTS + Radical Cation Assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant Capacity of Phenolic Phytochemicals from Various Cultivars of Plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colometric Methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The Conversion of Procyanidins and Prodelphinidins to Cyanidin and Delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef]
- Kassambara, A. Pipe-Friendly Framework for Basic Statistical Tests [R Package Rstatix Version 0.7.0]; Free Software Foundation Inc.: Boston, MA, USA, 2021. [Google Scholar]
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Agerer, R. Colour Atlas of Ectomycorrhizae 1987–2002; Einhorn-Verlag: Schwäbisch Gmünd, Germany, 1987. [Google Scholar]
- Agerer, R.; Rambold, G. DEEMY-An Information System for Characterization and Determination of Ectomycorrhizae 2004–2015; Ludwig-Maximilians-Universität: München, Germany, 2015. [Google Scholar]
- Hawley, G.L.; Taylor, A.F.S.; Dames, J.F. Ectomycorrhizas in Association with Pinus Patula in Sabie, South Africa. S. Afr. J. Sci. 2008, 104, 273–283. [Google Scholar] [CrossRef]
- Gil-Pelegrín, E.; Peguero-Pina, J.J.; Sancho-Knapik, D. (Eds.) Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Tree Physiology; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Allison, S.D.; Treseder, K.K. Warming and Drying Suppress Microbial Activity and Carbon Cycling in Boreal Forest Soils. Glob. Chang. Biol. 2008, 14, 2898–2909. [Google Scholar] [CrossRef]
- Beniwal, R.S.; Langenfeld-Heyser, R.; Polle, A. Ectomycorrhiza and Hydrogel Protect Hybrid Poplar from Water Deficit and Unravel Plastic Responses of Xylem Anatomy. Environ. Exp. Bot. 2010, 69, 189–197. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, V.; Tuteja, N. Mobilization of Micronutrients by Mycorrhizal Fungi. In Mycorrhiza—Function, Diversity, State of the Art; Springer: Berlin/Heidelberg, Germany, 2017; pp. 9–26. [Google Scholar] [CrossRef]
- Talbot, J.M.; Allison, S.D.; Treseder, K.K. Decomposers in Disguise: Mycorrhizal Fungi as Regulators of Soil C Dynamics in Ecosystems under Global Change. Funct. Ecol. 2008, 22, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J. Carbon Storage in Trees: Pathogens Have Their Say. Tree Physiol. 2014, 34, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Woodruff, D.R.; Meinzer, F.C. Carbon Dynamics in Trees: Feast or Famine? Tree Physiol. 2012, 32, 764–775. [Google Scholar] [CrossRef]
- Gojon, A.; Krouk, G.; Perrine-Walker, F.; Laugier, E. Nitrate Transceptor(s) in Plants. J. Exp. Bot. 2011, 62, 2299–2308. [Google Scholar] [CrossRef]
- Snoeijers, S.S.; Pérez-García, A.; Joosten, M.H.A.J.; de Wit, P.J.G.M. The Effect of Nitrogen on Disease Development and Gene Expression in Bacterial and Fungal Plant Pathogens. Eur. J. Plant Pathol. 2000, 106, 493–506. [Google Scholar] [CrossRef]
- Sanchez-Bel, P.; Troncho, P.; Gamir, J.; Pozo, M.J.; Camañes, G.; Cerezo, M.; Flors, V. The Nitrogen Availability Interferes with Mycorrhiza-Induced Resistance against Botrytis Cinerea in Tomato. Front. Microbiol. 2016, 7, 1598. [Google Scholar] [CrossRef]
- Pap, P.; Stojnić, S.; Nikolić, N.; Orlović, S.; Marković, M.; Vasić, V.; Stevanov, M. Impact of Erysiphe alphitoides (Griffon & Maubl.) U. Braun & S. Takam. on Leaf Physiological Parameters in Pedunculate Oak (Quercus robur L.) Saplings. Balt. For. 2014, 20, 2–9. [Google Scholar]
- Copolovici, L.; Väärtnõu, F.; Estrada, M.P.; Niinemets, Ü. Oak Powdery Mildew (Erysiphe alphitoides) Induced Volatile Emissions Scale with the Degree of Infection in Quercus robur. Tree Physiol. 2014, 34, 1399. [Google Scholar] [CrossRef]
- Hajji, M.; Dreyer, E.; Marçais, B. Impact of Erysiphe alphitoides on Transpiration and Photosynthesis in Quercus robur Leaves. Eur. J. Plant Pathol. 2009, 125, 63–72. [Google Scholar] [CrossRef]
- Liović, B. The Influence of Powdery Mildew (Microsphaera alphitoides Griff. et Maubl.) on Growth and Survival Rate of Oak Seedlings. Šumarski List 2011, 135, 122–129. Šumarski List 2011, 135, 122–129. [Google Scholar]
- Vaštag, E.E.; Kastori, R.R.; Orlović, S.S.; Bojović, M.M.; Kesić, L.A.; Pap, P.L.; Stojnić, S.M. Effects of Oak Powdery Mildew (Erysiphe alphitoides [Griffon and Maubl.] U. Braun and S. Takam.) on Photosynthesis of Pedunculate Oak (Quercus robur L.). Zb. Matice Srp. Prir. Nauk. 2019, 136, 43–56. [Google Scholar] [CrossRef]
- Luo, Z.-B.; Li, K.; Jiang, X.; Polle, A. Ectomycorrhizal Fungus (Paxillus involutus) and Hydrogels Affect Performance of Populus Euphratica Exposed to Drought Stress. Ann. For. Sci. 2009, 66, 106. [Google Scholar] [CrossRef]
- Kipfer, T.; Wohlgemuth, T.; van der Heijden, M.G.A.; Ghazoul, J.; Egli, S. Growth Response of Drought-Stressed Pinus sylvestris Seedlings to Single- and Multi-Species Inoculation with Ectomycorrhizal Fungi. PLoS ONE 2012, 7, e35275. [Google Scholar] [CrossRef]
- Lakso, A.N.; Pratt, C.; Pearson, R.C.; Pool, R.M.; Seem, R.C.; Welser, M.J. Photosynthesis, transpiration, and water use efficiency of mature grape leaves infected with Uncinula necator (powdery mildew). Phytopathology 1982, 72, 232–236. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Emery, S.M.; Rudgers, J.A. Fungal symbionts alter plant responses to global change. Am. J. Bot. 2013, 100, 1445–1457. [Google Scholar] [CrossRef]
- Wang, H.; Hao, Z.; Zhang, X.; Xie, W.; Chen, B. Arbuscular Mycorrhizal Fungi Induced Plant Resistance against Fusarium Wilt in Jasmonate Biosynthesis Defective Mutant and Wild Type of Tomato. J. Fungi 2022, 8, 422. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling Mycorrhiza-Induced Resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Schweiger, R.; Baier, M.C.; Persicke, M.; Müller, C. High Specificity in Plant Leaf Metabolic Responses to Arbuscular Mycorrhiza. Nat. Commun. 2014, 5, 3886. [Google Scholar] [CrossRef] [PubMed]
- Van Kal, J.A.L. Licensed to Kill: The Lifestyle of a Necrotrophic Plant Pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Sharma, S.; Villamor, J.G.; Verslues, P.E. Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol. 2011, 157, 292–304. [Google Scholar] [CrossRef]
- Signorelli, S.; Arellano, J.B.; Melø, T.B.; Borsani, O.; Monza, J. Proline does not quench singlet oxygen: Evidence to reconsider its protective role in plants. Plant Physiol. Biochem. 2013, 64, 80–83. [Google Scholar] [CrossRef]
- Fabro, G.; Kovács, I.; Pavet, V.; Szabados, L.; Alvarez, M.E. Proline Accumulation and AtP5CS2 Gene Activation Are Induced by Plant-Pathogen Incompatible Interactions in Arabidopsis. Mol. Plant-Microbe Interact. 2004, 17, 343–350. [Google Scholar] [CrossRef]
- Uchida, A.; Jagendorf, A.T.; Hibino, T.; Takabe, T.; Takabe, T. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci. 2002, 163, 515–523. [Google Scholar] [CrossRef]
- Lei, Y.; Yin, C.; Ren, J.; Li, C. Effect of Osmotic Stress and Sodium Nitroprusside Pretreatment on Proline Metabolism of Wheat Seedlings. Biol. Plant. 2007, 51, 386–390. [Google Scholar] [CrossRef]
- Tan, J.; Zhao, H.; Hong, J.; Han, Y.; Li, H.; Zhao, W. Effects of Exogenous Nitric Oxide on Photosynthesis, Antioxidant Capacity and Proline Accumulation in Wheat Seedlings Subjected to Osmotic Stress. World J. Agric. Sci. 2008, 4, 307–313. [Google Scholar]
- Chen, C.; Dickman, M.B. Proline Suppresses Apoptosis in the Fungal Pathogen Colletotrichum Trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed]
- Molinari, H.B.C.; Marur, C.J.; Daros, E.; Freitas de Campos, M.K.; Portela de Carvalho, J.F.R.; Bespalhok Filho, J.C.; Protasio Pereira, L.F.; Esteves Vieira, L.G. Evaluation of the Stress-inducible Production of Proline in Transgenic Sugarcane (Saccharum spp.): Osmotic Adjustment, Chlorophyll Fluorescence and Oxidative Stress. Physiol. Plant. 2007, 130, 218–229. [Google Scholar] [CrossRef]
- Alvarez-G. Omez, T.B.; Ramírez-Trujillo, J.A.; Ramírez-Yáñez, M.; Suárez-Rodríguez, R. Overexpression of SlERF3b and SlERF5 in Transgenic Tomato Alters Fruit Size, Number of Seeds and Promotes Early Flowering, Tolerance to Abiotic Stress and Resistance. Ann. Appl. Biol. 2021, 179, 382–394. [Google Scholar] [CrossRef]
- Kumar, P. Stress Amelioration Response of Glycine Betaine and Arbuscular Mycorrhizal Fungi in Sorghum under Cr Toxicity. PLoS ONE 2021, 16, e0253878. [Google Scholar] [CrossRef] [PubMed]
- Aslanpour, M.; Baneh, H.D.; Tehranifar, A.; Shoor, M. The effect of mycorrhizal fungi on the amount of glycine betaine, soluble sugar, proline, leaf water content and leaf chlorophyll of the white seedless grape under drought stress conditions. Int. J. Adv. Biotechnol. Res. 2016, 7, 1119–1133. [Google Scholar]
- Constabel, C.P.; Yoshida, K.; Walker, V. Diverse ecological roles of plant tannins: Plant defense and beyond. Recent Adv. Polyphen. Res. 2014, 4, 115–142. [Google Scholar]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. 17-Role of phenols and polyphenols in plant defense response to biotic and abiotic stresses. In Biocontrol Agents and Secondary Metabolites; Applications and Immunization for Plant Growth and Protection; Woodhead Publishing: Cambridge, UK, 2021; pp. 419–441. [Google Scholar]
- Daayf, F.; Ongena, M.; Boulanger, R.; el Hadrami, I.; Bélanger, R.R. Induction of Phenolic Compounds in Two Cultivars of Cucumber by Treatment of Healthy and Powdery Mildew-Infected Plants with Extracts of Reynoutria sachalinensis. J. Chem. Ecol. 2000, 26, 1579–1593. [Google Scholar] [CrossRef]
- Ashry, N.A.; Mohamed, H.I. Impact of Secondary Metabolites and Related Enzymes in Flax Resistance and/or Susceptibility to Powdery Mildew. World J. Agric. Sci. 2011, 7, 78–85. [Google Scholar] [CrossRef]
- Singh, D.P.; Srivastava, J.S.; Bahadur, A.; Singh, U.P.; Singh, S.K. Arbuscular mycorrhizal fungi induced biochemical changes in pea (Pisum sativum) and their effect on powdery mildew (Erysiphe pisi)/Arbusculäre Mykorrhizapilze induzieren biochemische Veränderungen in Erbsen (Pisum sativum) und ihre Wirkung auf Echten Mehltau (Erysiphe pisi). J. Plant Dis. Prot. 2004, 111, 266–272. [Google Scholar]
- Jugran, A.K.; Bahukhandi, A.; Dhyani, P.; Bhatt, I.D.; Rawal, R.S.; Nandi, S.K.; Palni, L.M.S. The effect of inoculation with mycorrhiza: AM on growth, phenolics, tannins, phenolic composition and antioxidant activity in Valeriana jatamansi Jones. J. Soil Sci. Plant Nutr. 2015, 15, 1036–1049. [Google Scholar] [CrossRef]
- Jing, X.; Wang, H.; Gong, B.; Liu, S.; Wei, M.; Ai, X.; Li, Y.; Shi, Q. Secondary and sucrose metabolism regulated by different light quality combinations involved in melon tolerance to powdery mildew. Plant Physiol. Biochem. 2018, 124, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Zhi-lin, Y.; Chuan-chao, D.; Lian-qing, C. Regulation and Accumulation of Secondary Metabolites in Plant-Fungus Symbiotic System. Afr. J. Biotechnol. 2007, 6, 1266–1271. [Google Scholar]
- Wu, Q.S.; Zou, Y.N.; Liu, C.Y.; Ting, L.U. Interacted effect of arbuscular mycorrhizal fungi and polyamines on root system architecture of citrus seedlings. J. Integr. Agric. 2012, 11, 1675–1681. [Google Scholar] [CrossRef]
- Zou, Y.N.; Zhang, F.; Srivastava, A.K.; Wu, Q.S.; Kuča, K. Arbuscular Mycorrhizal Fungi Regulate Polyamine Homeostasis in Roots of Trifoliate Orange for Improved Adaptation to Soil Moisture Deficit Stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Sannazzaro, A.I.; Echeverría, M.; Albertó, E.O.; Ruiz, O.A.; Menéndez, A.B. Modulation of polyamine balance in Lotus glaber by salinity and arbuscular mycorrhiza. Plant Physiol. Biochem. 2007, 45, 39–46. [Google Scholar] [CrossRef]
- Zhang, F.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizas modulate root polyamine metabolism to enhance drought tolerance of trifoliate orange. Environ. Exp. Bot. 2020, 171, 103926. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, Z.; Sun, H.; Yang, W.; Xu, H. The Response Patterns of Arbuscular Mycorrhizal and Ectomycorrhizal Symbionts Under Elevated CO2: A Meta-Analysis. Front. Microbiol. 2018, 11, 1248. [Google Scholar] [CrossRef]
- Shahzad, T.; Chenu, C.; Genet, P.; Barot, S.; Perveen, N.; Mougin, C.; Fontaine, S. Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species. Soil Biol. Biochem. 2015, 80, 146–155. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Balmer, A.; Pastor, V.; Gamir, J.; Flors, V.; Mauch-Mani, B. The ‘prime-ome’: Towards a holistic approach to priming. Trends Plant Sci. 2015, 20, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Cicatelli, A.; Lingua, G.; Todeschini, V.; Biondi, S.; Torrigiani, P.; Castiglione, S. Arbuscular mycorrhizal fungi restore normal growth in a white poplar clone grown on heavy metal-contaminated soil, and this is associated with upregulation of foliar metallothionein and polyamine biosynthetic gene expression. Ann. Bot. 2010, 106, 791–802. [Google Scholar] [CrossRef]
- Salloum, M.S.; Menduni, M.F.; Benavides, M.P.; Larrauri, M.; Luna, C.M.; Silvente, S. Polyamines and flavonoids: Key compounds in mycorrhizal colonization of improved and unimproved soybean genotypes. Symbiosis 2018, 76, 265–275. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.M.; Ibrahim, A.H.; Al-Amri, S.M.; Shoker, A.E. Synergistic effect of arbuscular mycorrhizal fungi and spermine on amelioration of salinity stress of wheat (Triticum aestivum L. cv. gimiza 9). Aust. J. Crop Sci. 2013, 7, 1525–1532. [Google Scholar]
- Talaat, N.B.; Shawky, B.T. Modulation of nutrient acquisition and polyamine pool in salt-stressed wheat (Triticum aestivum L.) plants inoculated with arbuscular mycorrhizal fungi. Acta Physiol. Plant. 2013, 35, 2601–2610. [Google Scholar] [CrossRef]
- Sharma, K.; Gupta, S.; Thokchom, S.D.; Jangir, P.; Kapoor, R. Arbuscular Mycorrhiza-Mediated Regulation of Polyamines and Aquaporins During Abiotic Stress: Deep Insights on the Recondite Players. Front. Plant Sci. 2021, 12, 642101. [Google Scholar] [CrossRef]
- Garg, N.; Sharma, A. Role of Putrescine (Put) in Imparting Salt Tolerance through Modulation of Put Metabolism, Mycorrhizal and Rhizobial Symbioses in Cajanus cajan (L.) Millsp. Symbiosis 2019, 79, 59–74. [Google Scholar] [CrossRef]
- Shaul-Keinan, O.; Gadkar, V.; Ginzberg, I.; Grünzweig, J.M.; Chet, I.; Elad, Y.; Wininger, S.; Belausov, E.; Eshed, Y.; Ben-Tal, Y.; et al. Hormone Concentrations in Tobacco Roots Change during Arbuscular Mycorrhizal Colonization with Glomus intraradices. New Phytol. 2002, 154, 501–507. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The interaction between foliar GA3 application and arbuscular mycorrhizal fungi inoculation improves growth in salinized tomato (Solanum lycopersicum L.) plants by modifying the hormonal balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Evelin, H.; Giri, B.; Kapoor, R. Ultrastructural Evidence for AMF Mediated Salt Stress Mitigation in Trigonella foenum-graecum. Mycorrhiza 2013, 23, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Saroy, K. Interactive Effects of Polyamines and Arbuscular Mycorrhiza in Modulating Plant Biomass, N2 Fixation, Ureide, and Trehalose Metabolism in Cajanus cajan (L.) Millsp. Genotypes under Nickel Stress. Environ. Sci. Pollut. Res. 2020, 27, 3043–3064. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Santiago, L.; Cervantes-Chávez, J.A.; Léon-Ramírez, C.G.; Ruiz-Herrera, J. Polyamine Metabolism in Fungi with Emphasis on Phytopathogenic Species. J. Amino Acids 2012, 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Plett, J.M.; Plett, K.L.; Wong-Bajracharya, J.; De Freitas Pereira, M.; Dutra Costa, M.; Kohler, A.; Martin, F.; Anderson, I.C.; Plett, J. Mycorrhizal Effector PaMiSSP10b Alters Polyamine Biosynthesis in Eucalyptus Root Cells and Promotes Root Colonization. New Phytol. 2020, 228, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Sebastiana, M.; Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Monteiro, F.; Figueiredo, A.; Maia, M.; Nascimento, R.; Silva, M.S.; Ferreira, A.N.; et al. Metabolomics and Transcriptomics to Decipher Molecular Mechanisms Underlying Ectomycorrhizal Root Colonization of an Oak Tree. Sci. Rep. 2021, 11, 8576. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.R. Polyamines and Plant Disease. Phytochemistry 2003, 64, 97–107. [Google Scholar] [CrossRef]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef]
- Majumdar, R.; Minocha, R.; Lebar, M.D.; Rajasekaran, K.; Long, S.; Carter-Wientjes, C.; Minocha, S.; Cary, J.W. Contribution of Maize Polyamine and Amino Acid Metabolism toward Resistance against Aspergillus Flavus Infection and Aflatoxin Production. Front. Plant Sci. 2019, 10, 692. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Kazan, K.; Praud, S.; Torney, F.J.; Rusu, A.; Manners, J.M. Early Activation of Wheat Polyamine Biosynthesis during Fusarium Head Blight Implicates Putrescine as an Inducer of Trichothecene Mycotoxin Production. BMC Plant Biol. 2010, 10, 289. [Google Scholar] [CrossRef]
- Rocha, R.O.; Elowsky, C.; Pham, N.T.; Wilson, R.A. Spermine-mediated tight sealing of the Magnaporthe oryzae appressorial pore–rice leaf surface interface. Nat. Microbiol. 2020, 5, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Popovic, R.B.; Kyle, D.J.; Cohen, A.S.; Zalik, S. Stabilization of thylakoid membranes by spermine during stress-induced senescence of barley leaf discs. Plant Physiol. 1979, 64, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Dobrovinskaya, O.R.; Muñiz, J.; Pottosin, I.I. Inhibition of Vacuolar Ion Channels by Polyamines. J. Membr. Biol. 1999, 167, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Montilla-Bascón, G.; Rubiales, D.; Prats, E. Changes in polyamine profile in host and non-host oat–powdery mildew interactions. Phytochem. Lett. 2014, 8, 207–212. [Google Scholar] [CrossRef]
- Montilla-Bascón, G.; Rubiales, D.; Altabella, T.; Prats, E. Free polyamine and polyamine regulation during pre-penetration and penetration resistance events in oat against crown rust (Puccinia coronata f. sp. avenae). Plant Pathol. 2016, 65, 392–401. [Google Scholar] [CrossRef]
- Greenland, A.J.; Lewis, D.H. Amines in barley laves infected by brown rust and their possible relevance to formation of ‘green islands’. New Phytol. 1984, 96, 283–291. [Google Scholar] [CrossRef]
- Lowe-Power, T.M.; Hendrich, C.G.; von Roepenack-Lahaye, E.; Li, B.; Wu, D.; Mitra, R.; Dalsing, B.L.; Ricca, P.; Naidoo, J.; Cook, D.; et al. Metabolomics of Tomato Xylem Sap during Bacterial Wilt Reveals Ralstonia solanacearum Produces Abundant Putrescine, a Metabolite That Accelerates Wilt Disease. Environ. Microbiol. 2018, 20, 1330–1349. [Google Scholar] [CrossRef]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with Regulatory Functions in Plant Abiotic Stress Tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Anwar, R.; Mattoo, A.K.; Handa, A.K. Polyamine interactions with plant hormones: Crosstalk at several levels. In Polyames; Springer: Tokyo, Japan, 2015; pp. 267–302. [Google Scholar] [CrossRef]
- Yamasaki, H.; Cohen, M.F. NO signal at the crossroads: Polyamine-induced nitric oxide synthesis in plants? Trends Plant Sci. 2006, 11, 522–524. [Google Scholar] [CrossRef]
- Tailor, A.; Tandon, R.; Bhatla, S.C. Nitric Oxide Modulates Polyamine Homeostasis in Sunflower Seedling Cotyledons under Salt Stress. Plant Signal. Behav. 2019, 14, 1667730. [Google Scholar] [CrossRef]
- Sharma, S.; Pareek, S.; Sagar, N.A.; Valero, D.; Serrano, M. Modulatory effects of exogenously applied polyamines on postharvest physiology, antioxidant system and shelf life of fruits: A review. Int. J. Mol. Sci. 2017, 18, 1789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, L.; Li, F.; Meng, D.; Sheng, J. Methyl salicylate-induced arginine catabolism is associated with up-regulation of polyamine and nitric oxide levels and improves chilling tolerance in cherry tomato fruit. J. Agric. Food Chem. 2011, 59, 9351–9357. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Tiburcio, A.F. Plant Polyamines in Stress and Development: An Emerging Area of Research in Plant Sciences. Front. Plant Sci. 2014, 5, 319. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Basu, S.; Kumar, G. Polyamines metabolism: A way ahead for abiotic stress tolerance in crop plants. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Academic Press: Cambridge, MA, USA, 2018; pp. 39–55. [Google Scholar]
- Wang, D.; Li, L.; Xu, Y.; Limwachiranon, J.; Li, D.; Ban, Z.; Luo, Z. Effect of Exogenous Nitro Oxide on Chilling Tolerance, Polyamine, Proline, and γ-Aminobutyric Acid in Bamboo Shoots (Phyllostachys praecox f. Prevernalis). ACS Publ. 2017, 65, 5607–5613. [Google Scholar] [CrossRef] [PubMed]
- Priya, M.; Sharma, L.; Kaur, R.; Bindumadhava, H.; Nair, R.M.; Siddique, K.H.M.; Nayyar, H. GABA (γ-aminobutyric acid), as a thermo-protectant, to improve the reproductive function of heat-stressed mungbean plants. Sci. Rep. 2019, 9, 7788. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kebert, M.; Kostić, S.; Zlatković, M.; Stojnic, S.; Čapelja, E.; Zorić, M.; Kiprovski, B.; Budakov, D.; Orlović, S. Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L. Forests 2022, 13, 1491. https://doi.org/10.3390/f13091491
Kebert M, Kostić S, Zlatković M, Stojnic S, Čapelja E, Zorić M, Kiprovski B, Budakov D, Orlović S. Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L. Forests. 2022; 13(9):1491. https://doi.org/10.3390/f13091491
Chicago/Turabian StyleKebert, Marko, Saša Kostić, Milica Zlatković, Srdjan Stojnic, Eleonora Čapelja, Martina Zorić, Biljana Kiprovski, Dragana Budakov, and Saša Orlović. 2022. "Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L." Forests 13, no. 9: 1491. https://doi.org/10.3390/f13091491