
Developing and Comparing Individual Tree Growth Models of Major Coniferous Species in South Korea Based on Stem Analysis Data



Abstract
:1. Introduction
2. Materials and Methods
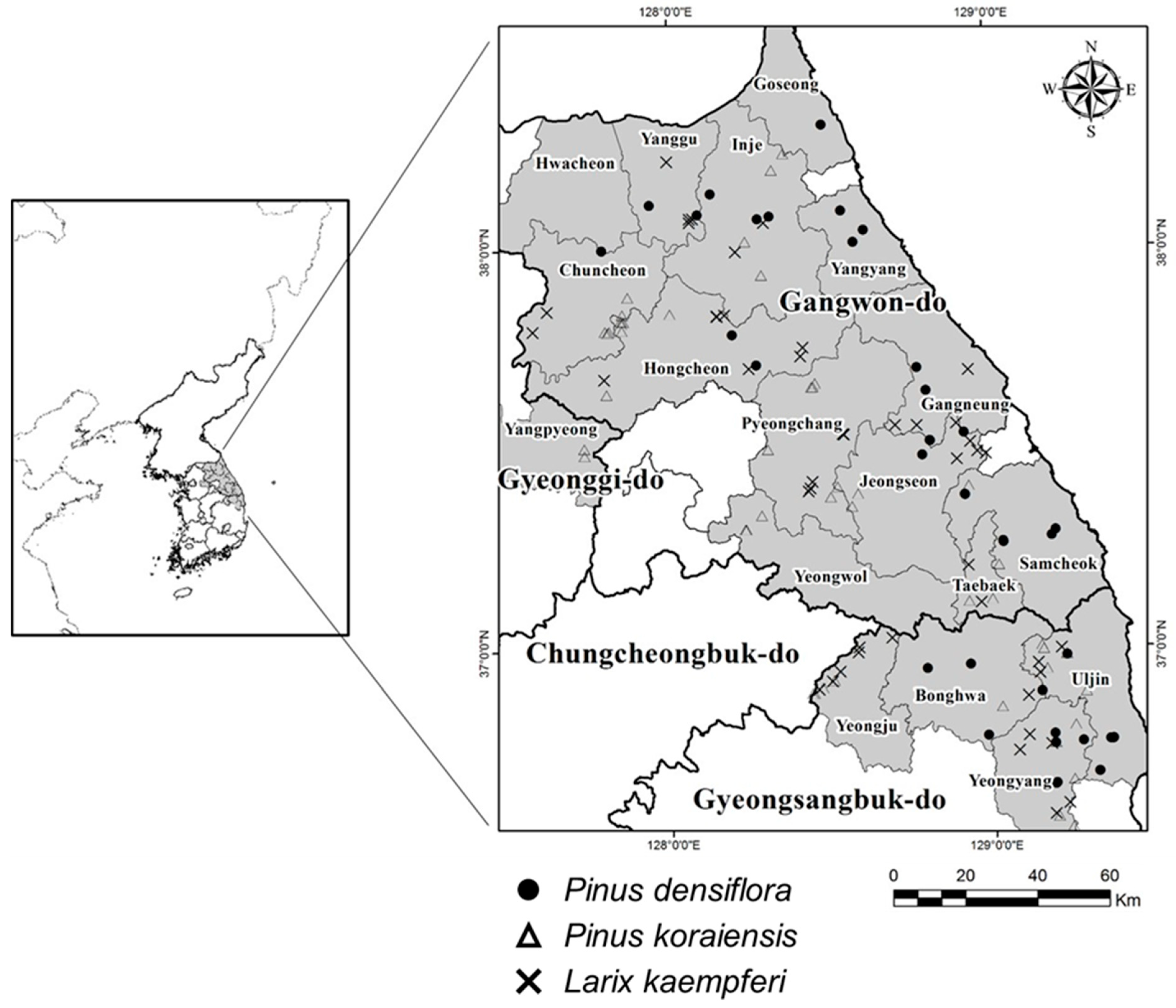
2.1. Study Area
2.1.1. Location and Site Characteristics
2.1.2. Plot Characteristic with Stand Density
2.2. Data Collection
2.2.1. Sample Tree Selection and Wood Disc Collection
2.2.2. Tree Ring Measurements and Stem Analysis
2.3. Modeling Approach
2.3.1. Selection of Candidate Growth Functions
2.3.2. Statistical Modeling and Validation
3. Results and Discussion
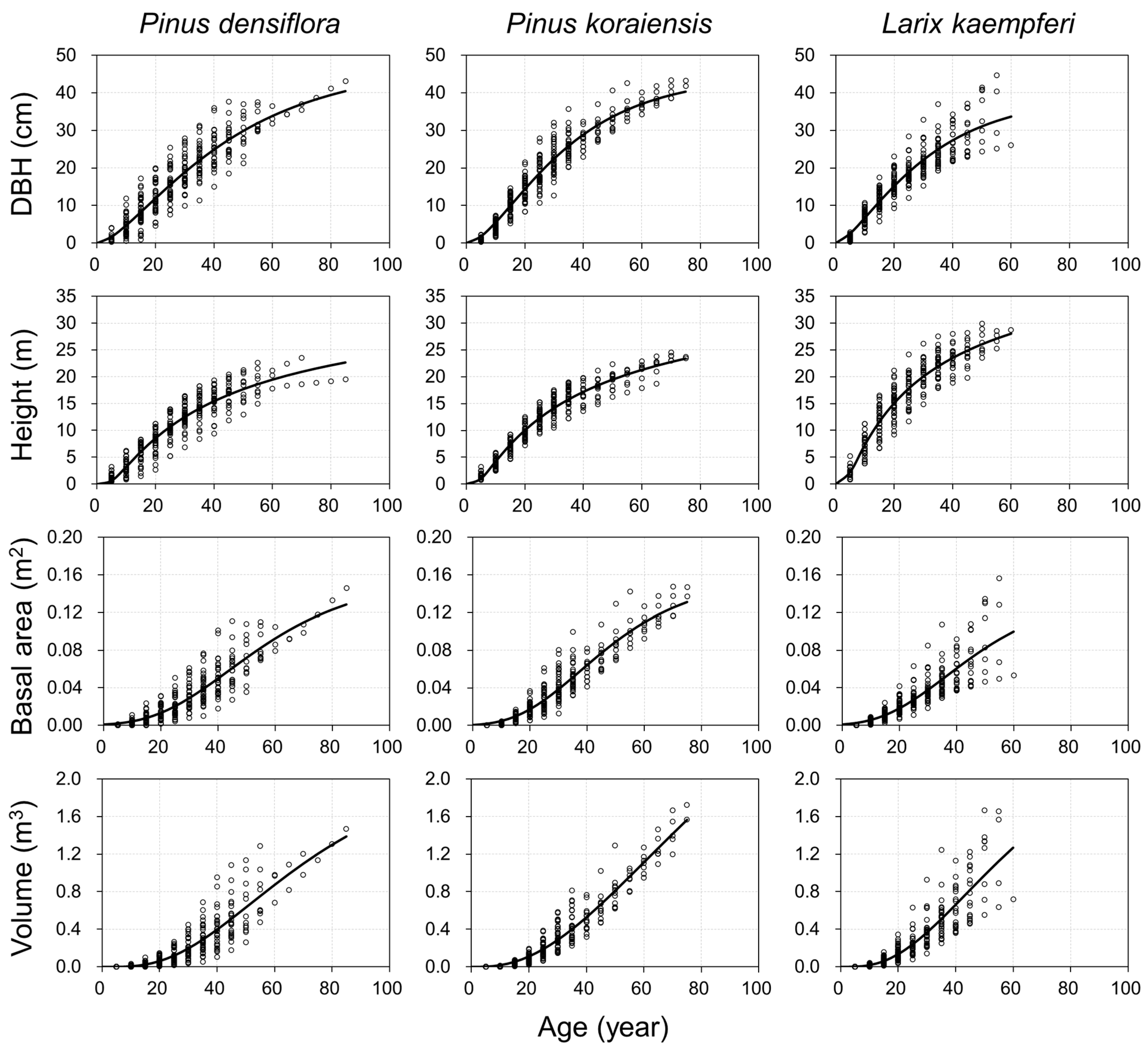
3.1. Parameters Estimates and Fit Statistics
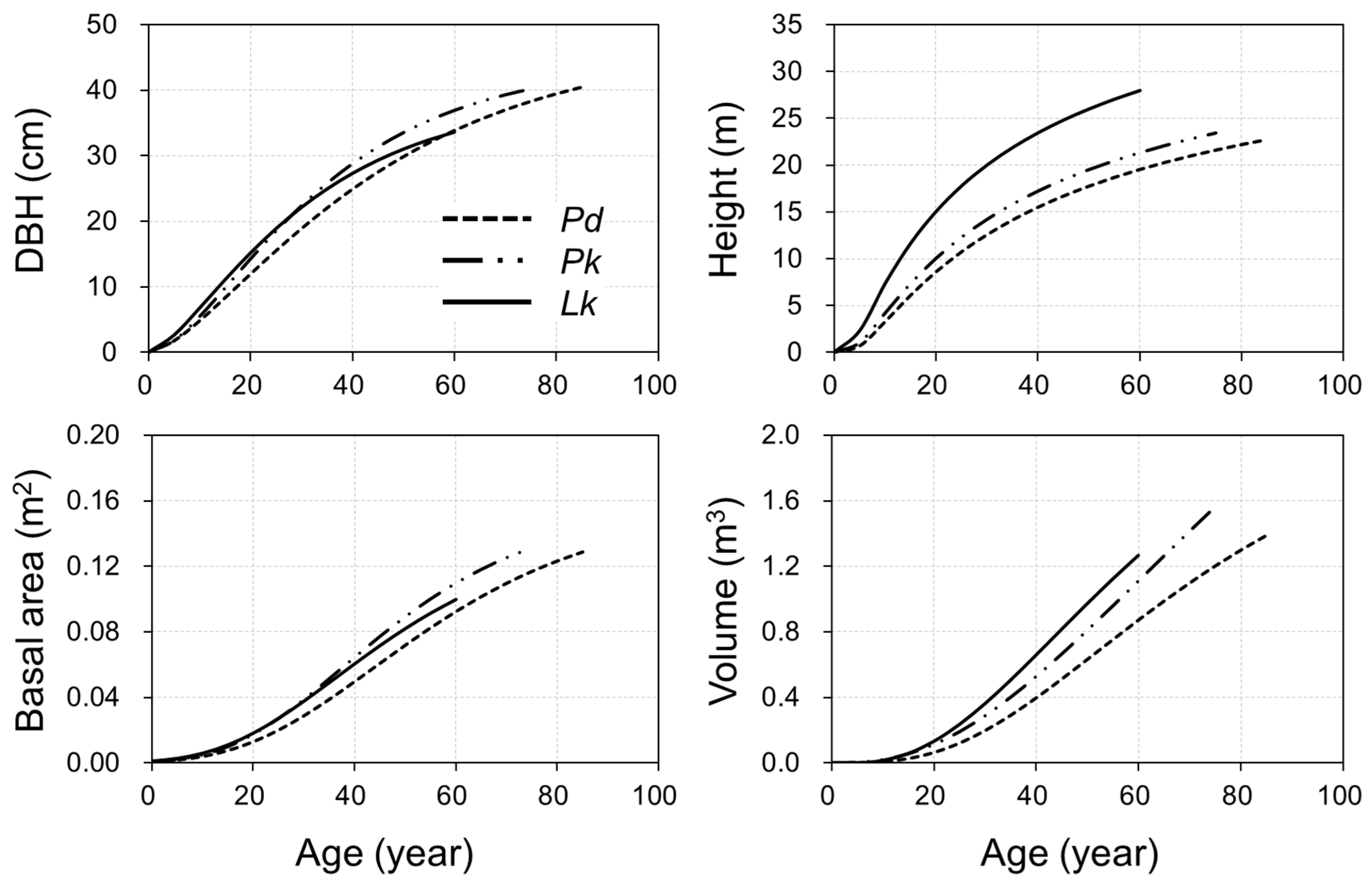
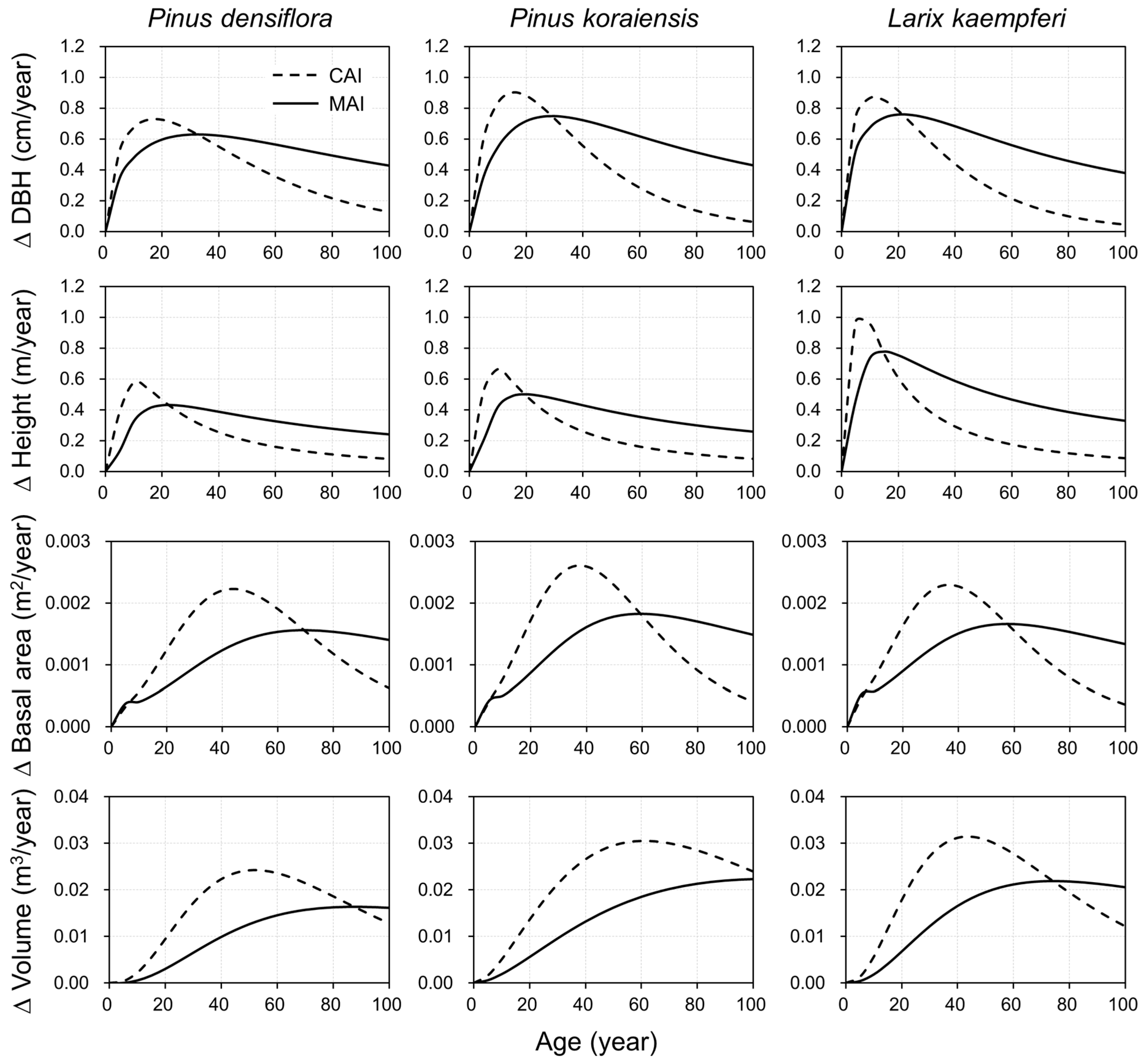
3.2. Growth Simulation and Characteristics
3.3. Model Evaluation and Applicability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korea Forest Research Institute. Commercial Tree Species 1 Korean Red Pine (Pinus densiflora Siebold & Zucc.); Korea Forest Research Institute: Seoul, Republic of Korea, 2012; ISBN 11-1400377-000569-01. [Google Scholar]
- Korea Forest Research Institute. Commercial Tree Species 3 Korean White Pine (Pinus koraiensis Siebold & Zucc.); Korea Forest Research Institute: Seoul, Republic of Korea, 2012; ISBN 11-1400377-000550-01. [Google Scholar]
- Korea Forest Research Institute. Commercial Tree Species 4 Japanese Larch (Larix kaempferi (Lamb.) Carriere); Korea Forest Research Institute: Seoul, Republic of Korea, 2012; ISBN 11-1400377-000551-01. [Google Scholar]
- Korea Forest Service. 2020 Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2021; ISBN 11-1400000-000001-10. [Google Scholar]
- Yang, R.C.; Kozak, A.; Smith, J.H.G. The potential of Weibull-type functions as flexible growth curves. Can. J. For. Res. 1978, 8, 424–431. [Google Scholar] [CrossRef]
- Tomé, J.; Tomé, M.; Barreiro, S.; Paulo, J.A. Age-independent difference equations for modelling tree and stand growth. Can. J. For. Res. 2006, 36, 1621–1630. [Google Scholar] [CrossRef]
- Parresol, B.R.; Devall, M.S. Patterns of Diametric Growth in Stem-Analyzed Laurel Trees (Cordia alliodora) in a Panamanian Forest. Southwest. Nat. 2013, 58, 170–178. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer Science & Business Media: New York, NY, USA, 2012; ISBN 978-94-007-1597-4. [Google Scholar]
- Monserud, R.A. Height growth and site index curves for inland Douglas-fir based on stem analysis data and forest habitat type. For. Sci. 1984, 30, 943–965. [Google Scholar]
- Zeide, B. Analysis of Growth Equations. For. Sci. 1993, 39, 594–616. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Avery, T.E.; Bullock, B.P. Forest Measurements, 6th ed.; Waveland Press Inc.: Long Grove, IL, USA, 2018; ISBN 978-1-4786-3618-2. [Google Scholar]
- García, O. Stand Growth models: Theory and practice. In Proceedings of the Advancement in Forest Inventory and Forest Management Sciences—Proceedings of the IUFRO Seoul Conference, Seoul, Republic of Korea, 20–25 September 1993; Forestry Research Institute of the Republic of Korea. pp. 22–45. [Google Scholar]
- Ito, T. A System of growth models for even-aged pure stands based on the Richards growth function (I) A growth model for average height. J. Jpn. For. Soc. 1987, 69, 221–227. [Google Scholar]
- Ito, T. A System of growth models for even-aged pure stands based on the Richards growth function (II) A growth model for stand volume. J. Jpn. For. Soc. 1987, 69, 391–397. [Google Scholar]
- Ito, T. A system of growth models for even-aged pure stands based on the Richards growth function (IV) Estimation of the parameters using data from a temporary sample plot and stem-analyzed sample trees. J. Jpn. For. Soc. 1988, 70, 95–103. [Google Scholar]
- Ito, T. A system of growth models for even-aged pure stands based on the Richards growth function (III) Estimation of the parameters using data from remeasured temporary sample plots. J. Jpn. For. Soc. 1988, 70, 57–63. [Google Scholar] [CrossRef]
- Li, F.; Zhao, B.; Su, G. A derivation of the generalized Korf growth equation and its application. J. For. Res. 2000, 11, 81–88. [Google Scholar] [CrossRef]
- Johansson, T. A site dependent top height growth model for hybrid aspen. J. For. Res. 2013, 24, 691–698. [Google Scholar] [CrossRef]
- Pommerening, A.; Muszta, A. Methods of modelling relative growth rate. For. Ecosyst. 2015, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.-W.; Balekoglu, S.; Choi, J.-K. Growth pattern analysis by stem analysis of Korean white pine (Pinus koraiensis) in the central northern region of Korea. For. Sci. Technol. 2014, 10, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Lee, D.; Choi, J. Growth Analysis of Red Pine (Pinus densiflora) by Stem Analysis in the Eastern Region of Korea. J. For. Environ. Sci. 2015, 31, 47–54. [Google Scholar] [CrossRef]
- Seo, Y.; Lee, D.; Choi, J. Growth pattern analysis of major coniferous tree species in South Korea. For. Sci. Technol. 2018, 14, 1–6. [Google Scholar] [CrossRef]
- Korea Meteorological Administration. Climate Characteristics by Region in Korea. Available online: https://www.weather.go.kr/w/obs-climate/climate/korea-climate/regional-char.do?area=2 (accessed on 15 September 2022).
- Lee, D.; Choi, J. Evaluating maximum stand density and size–density relationships based on the Competition Density Rule in Korean pines and Japanese larch. For. Ecol. Manag. 2019, 446, 204–213. [Google Scholar] [CrossRef]
- Lee, D.; Choi, J. Stocking Diagrams for Silvicultural Guideline in Korean Pines and Japanese Larch. Forests 2020, 11, 833. [Google Scholar] [CrossRef]
- Husch, B.; Beers, T.W.; Kershaw, J.A., Jr. Forest Mensuration, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2002; ISBN 978-0-471-01850-6. [Google Scholar]
- Kiviste, A. Mathematical Functions of Forest Growth; Estonian Agricultural Academy: Tartu, Estonia, 1988; p. 108, Supplement 171p. [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Pukkala, T.; Lähde, E.; Laiho, O. Species Interactions in the Dynamics of Even- and Uneven-Aged Boreal Forests. J. Sustain. For. 2013, 32, 371–403. [Google Scholar] [CrossRef]
- Liao, C.Y.; Podrázský, V.V.; Liu, G.B. Diameter and height growth analysis for individual White Pine trees in the area of Kostelec nad Černými lesy. J. For. Sci. 2003, 49, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Kwak, D.A.; Lee, W.K.; Kim, S.W.; Seo, J.H. Regional differences in height and tper form of Pinus densiflora in Korea. Korean J. For. Meas. 2004, 7, 49–60. [Google Scholar]
- Son, Y.M.; Lee, K.H.; Kweon, S.D. Estimation of stem taper equation and growth pattern in Pinus koraiensis by two districts. Korean J. For. Meas. 2004, 7, 9–14. [Google Scholar]
- Perez, D. Growth and volume equations developed from stem analysis for Tectona grandis in Costa Rica. J. Trop. For. Sci. 2008, 20, 66–75. [Google Scholar]
- Zuidema, P.A.; Vlam, M.; Chien, P.D. Ages and long-term growth patterns of four threatened Vietnamese tree species. Trees 2011, 25, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Chiba, Y. A Simple Mathematical Model of Growth Pattern in Tree Stems. Ann. Bot. 1994, 73, 91–98. [Google Scholar] [CrossRef]
- Brienen, R.J.; Zuidema, P.A. Lifetime growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. J. Ecol. 2006, 94, 481–493. [Google Scholar] [CrossRef]
- Jeon, B.-H.; Lee, S.-H.; Lee, Y.-J.; Kim, H.; Kang, H.-M. Estimation of site index and stem volume equations for Larix leptolepis stand in Jinan, Chonbuk. J. Korean For. Soc. 2007, 96, 40–47. [Google Scholar]
- Inoue, S.; Shirota, T.; Mitsuda, Y.; Ishii, H.; Gyokusen, K. Effects of individual size, local competition and canopy closure on the stem volume growth in a monoclonal Japanese cedar (Cryptomeria japonica D. Don) plantation. Ecol. Res. 2008, 23, 953–964. [Google Scholar] [CrossRef]
- Castagneri, D.; Storaunet, K.O.; Rolstad, J. Age and growth patterns of old Norway spruce trees in Trillemarka forest, Norway. Scand. J. For. Res. 2013, 28, 232–240. [Google Scholar] [CrossRef]
- Tian, N.; Fan, Z.; Matney, T.G.; Schultz, E.B. Growth and Stem Profiles of Invasive Triadica sebifera in the Mississippi Coast of the United States. For. Sci. 2017, 63, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-K.; von Gadow, K.; Chung, D.-J.; Lee, J.-L.; Shin, M.-Y. DBH growth model for Pinus densiflora and Quercus variabilis mixed forests in central Korea. Ecol. Model. 2004, 176, 187–200. [Google Scholar] [CrossRef]
- Sawata, S.; Nishizono, T.; Awaya, Y.; Nobori, Y. Analysis of Stem Growth Pattern in Japanese Cedar (Cryptomeria japonica) Trees in a Natural Forest in Akita, Northeastern Japan. J. Jpn. For. Soc. 2007, 89, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Choi, J. Development of Variable-Density Yield Models with Site Index Estimation for Korean Pines and Japanese Larch. Forests 2022, 13, 1150. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Statistics | Age (Year) | DBH (cm) | Height (m) | Volume (m3) | No. of Trees | No. of Sample Points at 5-Year Interval |
|---|---|---|---|---|---|---|---|
| Pinus densiflora | Mean | 47 | 30.5 | 17.0 | 0.6707 | 38 | 331 |
| SD | 16 | 8.4 | 4.0 | 0.3940 | |||
| Minimum | 9 | 5.2 | 3.6 | 0.0067 | |||
| Maximum | 99 | 47.3 | 23.8 | 1.6448 | |||
| Pinus koraiensis | Mean | 40 | 29.8 | 17.2 | 0.6935 | 46 | 351 |
| SD | 16 | 7.9 | 4.4 | 0.4743 | |||
| Minimum | 15 | 13.9 | 7.4 | 0.0527 | |||
| Maximum | 77 | 45.9 | 24.6 | 1.9642 | |||
| Larix kaempferi | Mean | 38 | 27.8 | 22.8 | 0.7480 | 45 | 321 |
| SD | 12 | 7.9 | 4.4 | 0.5160 | |||
| Minimum | 19 | 17.0 | 12.2 | 0.1614 | |||
| Maximum | 60 | 47.9 | 30.6 | 2.2803 |
| Function | Integral Form | Differential Form |
|---|---|---|
| Logistic | ||
| Korf | ||
| Gompertz | ||
| Chapman-Richards | ||
| Weibull |
| Species | Variables | Parm a | Parm b | Parm c | Fit Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | S.E. | Estimate | S.E. | Estimate | S.E. | R2 | RMSE | MAE | MAPE | ||
| Pinus densiflora (n = 303) | Logistic | 35.7659 (<0.0001) | 1.3117 | 0.0847 (<0.0001) | 0.0053 | 11.8485 (<0.0001) | 1.2272 | 0.8291 | 4.0989 | 3.2799 | 0.5316 |
| Korf | 134.3419 (0.0113) | 52.7221 | 10.8703 (<0.0001) | 1.6634 | 0.5042 (<0.0001) | 0.1035 | 0.8412 | 3.9507 | 3.1056 | 0.3499 | |
| Gompertz | 39.8321 (<0.0001) | 1.9710 | 3.3558 (<0.0001) | 0.1921 | 0.0499 (<0.0001) | 0.0040 | 0.8375 | 3.9962 | 3.1779 | 0.4498 | |
| Chapman-Richards | 47.4990 (<0.0001) | 4.7042 | 0.0278 (<0.0001) | 0.0053 | 1.6201 (<0.0001) | 0.1748 | 0.8410 | 3.9537 | 3.1232 | 0.3843 | |
| Weibull | 44.1380 (<0.0001) | 4.2293 | 0.0047 (<0.0001) | 0.0010 | 1.4011 (<0.0001) | 0.0886 | 0.8405 | 3.9590 | 3.1357 | 0.3984 | |
| Pinus koraiensis (n = 324) | Logistic | 37.5633 (<0.0001) | 0.6866 | 0.0980 (<0.0001) | 0.0040 | 12.7995 (<0.0001) | 1.0008 | 0.9125 | 3.2541 | 2.5803 | 0.4549 |
| Korf | 82.1652 (<0.0001) | 9.9198 | 14.0460 (<0.0001) | 1.8092 | 0.7005 (<0.0001) | 0.0657 | 0.9315 | 2.8781 | 2.1464 | 0.1926 | |
| Gompertz | 40.3021 (<0.0001) | 0.9038 | 3.5817 (<0.0001) | 0.1525 | 0.0606 (<0.0001) | 0.0028 | 0.9243 | 3.0261 | 2.3421 | 0.3319 | |
| Chapman-Richards | 44.5393 (<0.0001) | 1.6455 | 0.0393 (<0.0001) | 0.0035 | 1.8676 (<0.0001) | 0.1294 | 0.9294 | 2.9217 | 2.2029 | 0.2484 | |
| Weibull | 42.2732 (<0.0001) | 1.5261 | 0.0047 (<0.0001) | 0.0007 | 1.4882 (<0.0001) | 0.0567 | 0.9280 | 2.9509 | 2.2372 | 0.2784 | |
| Larix kaempferi (n = 314) | Logistic | 30.9468 (<0.0001) | 0.8779 | 0.1130 (<0.0001) | 0.0069 | 10.8427 (<0.0001) | 1.1272 | 0.8507 | 3.6823 | 2.8871 | 0.5809 |
| Korf | 86.7510 (0.0002) | 22.6673 | 9.5077 (<0.0001) | 1.2776 | 0.5696 (<0.0001) | 0.0951 | 0.8744 | 3.3769 | 2.5151 | 0.2813 | |
| Gompertz | 33.3797 (<0.0001) | 1.2273 | 3.2992 (<0.0001) | 0.1944 | 0.0708 (<0.0001) | 0.0051 | 0.8634 | 3.5213 | 2.7202 | 0.4476 | |
| Chapman-Richards | 39.1338 (<0.0001) | 3.0142 | 0.0398 (<0.0001) | 0.0068 | 1.5779 (<0.0001) | 0.1638 | 0.8712 | 3.4201 | 2.5994 | 0.3629 | |
| Weibull | 37.5837 (<0.0001) | 3.1412 | 0.0091 (<0.0001) | 0.0016 | 1.3459 (<0.0001) | 0.085 | 0.8701 | 3.4339 | 2.6253 | 0.3937 | |
| Species | Variables | Parm a | Parm b | Parm c | Fit Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | S.E. | Estimate | S.E. | Estimate | S.E. | R2 | RMSE | MAE | MAPE | ||
| Pinus densiflora (n = 331) | Logistic | 18.7523 (<0.0001) | 0.3992 | 0.1097 (<0.0001) | 0.0055 | 12.763 (<0.0001) | 1.2794 | 0.8825 | 2.0430 | 1.5882 | 0.3479 |
| Korf | 38.8374 (<0.0001) | 5.6742 | 12.832 (<0.0001) | 2.1414 | 0.7140 (<0.0001) | 0.0871 | 0.8848 | 2.0225 | 1.5436 | 0.2555 | |
| Gompertz | 20.1584 (<0.0001) | 0.5500 | 3.5208 (<0.0001) | 0.1950 | 0.0674 (<0.0001) | 0.0039 | 0.8883 | 1.9921 | 1.5135 | 0.2848 | |
| Chapman-Richards | 21.7626 (<0.0001) | 0.9170 | 0.0460 (<0.0001) | 0.0049 | 1.8879 (<0.0001) | 0.1715 | 0.8886 | 1.9890 | 1.5016 | 0.2519 | |
| Weibull | 20.5390 (<0.0001) | 0.7875 | 0.0055 (<0.0001) | 0.0011 | 1.5140 (<0.0001) | 0.0718 | 0.8890 | 1.9857 | 1.4970 | 0.2588 | |
| Pinus koraiensis (n = 351) | Logistic | 20.5229 (<0.0001) | 0.2727 | 0.1128 (<0.0001) | 0.0037 | 11.6502 (<0.0001) | 0.7722 | 0.9348 | 1.6225 | 1.3120 | 0.2559 |
| Korf | 41.1095 (<0.0001) | 3.2831 | 11.3164 (<0.0001) | 1.0129 | 0.6949 (<0.0001) | 0.0485 | 0.9510 | 1.4074 | 1.0828 | 0.1383 | |
| Gompertz | 21.7925 (<0.0001) | 0.3299 | 3.3721 (<0.0001) | 0.1156 | 0.0706 (<0.0001) | 0.0024 | 0.9461 | 1.4761 | 1.1706 | 0.1881 | |
| Chapman-Richards | 23.6574 (<0.0001) | 0.5455 | 0.0459 (<0.0001) | 0.0029 | 1.7276 (<0.0001) | 0.0914 | 0.9504 | 1.4154 | 1.0956 | 0.1468 | |
| Weibull | 22.8045 (<0.0001) | 0.5183 | 0.0082 (<0.0001) | 0.0009 | 1.4080 (<0.0001) | 0.0417 | 0.9492 | 1.4326 | 1.1195 | 0.1621 | |
| Larix kaempferi (n = 321) | Logistic | 24.6783 (<0.0001) | 0.4069 | 0.1341 (<0.0001) | 0.0063 | 9.8793 (<0.0001) | 0.8678 | 0.8995 | 2.4378 | 1.9596 | 0.2788 |
| Korf | 48.1994 (<0.0001) | 5.3799 | 9.2619 (<0.0001) | 1.0353 | 0.6928 (<0.0001) | 0.0697 | 0.9156 | 2.2340 | 1.7319 | 0.1806 | |
| Gompertz | 26.0162 (<0.0001) | 0.5230 | 3.1811 (<0.0001) | 0.1575 | 0.0873 (<0.0001) | 0.0044 | 0.9095 | 2.3140 | 1.8123 | 0.2247 | |
| Chapman-Richards | 28.1977 (<0.0001) | 0.9276 | 0.0570 (<0.0001) | 0.0055 | 1.6441 (<0.0001) | 0.1279 | 0.9142 | 2.2528 | 1.7540 | 0.2001 | |
| Weibull | 27.2797 (<0.0001) | 0.9147 | 0.0133 (<0.0001) | 0.0019 | 1.3617 (<0.0001) | 0.0599 | 0.9132 | 2.2658 | 1.7651 | 0.2093 | |
| Species | Variables | Parm a | Parm b | Parm c | Fit Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | S.E. | Estimate | S.E. | Estimate | S.E. | R2 | RMSE | MAE | MAPE | ||
| Pinus densiflora (n = 303) | Logistic | 0.1222 (<0.0001) | 0.0075 | 0.0824 (<0.0001) | 0.0054 | 40.6221 (<0.0001) | 6.0108 | 0.7974 | 0.0132 | 0.0098 | 9.3810 |
| Korf | 1.2308 (0.2652) | 1.1027 | 20.7981 (0.0001) | 5.3120 | 0.5068 (0.0005) | 0.1447 | 0.8092 | 0.0128 | 0.0090 | 1.5625 | |
| Gompertz | 0.1578 (<0.0001) | 0.0161 | 5.3828 (<0.0001) | 0.3859 | 0.0384 (<0.0001) | 0.0039 | 0.8067 | 0.0129 | 0.0093 | 4.8037 | |
| Chapman-Richards | 0.2152 (<0.0001) | 0.0486 | 0.0215 (0.0002) | 0.0056 | 2.6596 (<0.0001) | 0.3882 | 0.8088 | 0.0128 | 0.0091 | 2.3751 | |
| Weibull | 0.1632 (<0.0001) | 0.0281 | 0.0002 (0.0081) | 0.0001 | 2.0586 (<0.0001) | 0.1441 | 0.8079 | 0.0128 | 0.0092 | 3.0804 | |
| Pinus koraiensis (n = 324) | Logistic | 0.1300 (<0.0001) | 0.0043 | 0.0902 (<0.0001) | 0.0043 | 37.9451 (<0.0001) | 4.1735 | 0.9000 | 0.0112 | 0.0082 | 9.7820 |
| Korf | 0.7276 (0.0153) | 0.2985 | 22.6280 (<0.0001) | 4.5783 | 0.6046 (<0.0001) | 0.1010 | 0.9141 | 0.0104 | 0.0068 | 0.6608 | |
| Gompertz | 0.1581 (<0.0001) | 0.0085 | 5.3982 (<0.0001) | 0.2984 | 0.0449 (<0.0001) | 0.0030 | 0.9103 | 0.0106 | 0.0073 | 4.2698 | |
| Chapman-Richards | 0.1992 (<0.0001) | 0.0226 | 0.0272 (<0.0001) | 0.0042 | 2.7702 (<0.0001) | 0.2860 | 0.9131 | 0.0104 | 0.0070 | 1.6581 | |
| Weibull | 0.1597 (<0.0001) | 0.0145 | 0.0003 (0.0002) | 0.0001 | 2.0530 (<0.0001) | 0.1011 | 0.9118 | 0.0105 | 0.0072 | 2.5677 | |
| Larix kaempferi (n = 314) | Logistic | 0.1048 (<0.0001) | 0.0093 | 0.0976 (<0.0001) | 0.0086 | 35.2623 (<0.0001) | 5.8297 | 0.7502 | 0.0137 | 0.0089 | 10.6329 |
| Korf | 9.3665 (0.7741) | 32.6058 | 14.8994 (<0.0001) | 0.6978 | 0.2927 (0.1204) | 0.1880 | 0.7567 | 0.0135 | 0.0082 | 2.1481 | |
| Gompertz | 0.1420 (<0.0001) | 0.0238 | 4.9745 (<0.0001) | 0.3850 | 0.0440 (<0.0001) | 0.0066 | 0.7550 | 0.0136 | 0.0085 | 6.1127 | |
| Chapman-Richards | 0.2583 (0.1026) | 0.1578 | 0.0178 (0.1013) | 0.0108 | 2.1727 (<0.0001) | 0.4391 | 0.7567 | 0.0136 | 0.0083 | 2.9777 | |
| Weibull | 0.1616 (0.0201) | 0.0691 | 0.0004 (0.0039) | 0.0001 | 1.9114 (<0.0001) | 0.2065 | 0.7567 | 0.0135 | 0.0083 | 3.3699 | |
| Species | Variables | Parm a | Parm b | Parm c | Fit Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | S.E. | Estimate | S.E. | Estimate | S.E. | R2 | RMSE | MAE | MAPE | ||
| Pinus densiflora (n = 303) | Logistic | 1.2895 (<0.0001) | 0.085 | 0.0899 (<0.0001) | 0.0061 | 88.0312 (<0.0001) | 16.9867 | 0.7866 | 0.1326 | 0.0891 | 6.8688 |
| Korf | 8.2089 (0.1461) | 5.6334 | 43.3596 (0.0265) | 19.4449 | 0.7223 (0.0001) | 0.1831 | 0.7957 | 0.1298 | 0.0817 | 1.0726 | |
| Gompertz | 1.7361 (<0.0001) | 0.204 | 7.1821 (<0.0001) | 0.6973 | 0.0394 (<0.0001) | 0.0043 | 0.7946 | 0.1301 | 0.0832 | 2.6467 | |
| Chapman-Richards | 2.1471 (<0.0001) | 0.4443 | 0.0268 (<0.0001) | 0.0060 | 4.0338 (<0.0001) | 0.7078 | 0.7955 | 0.1298 | 0.0820 | 1.4566 | |
| Weibull | 1.5350 (<0.0001) | 0.2069 | 0.00002 (0.0767) | 0.00001 | 2.6495 (<0.0001) | 0.1816 | 0.7944 | 0.1302 | 0.0832 | 2.1675 | |
| Pinus koraiensis (n = 323) | Logistic | 1.6238 (<0.0001) | 0.0694 | 0.0844 (<0.0001) | 0.0037 | 64.6695 (<0.0001) | 6.4897 | 0.9200 | 0.1030 | 0.0756 | 12.2879 |
| Korf | 45.0916 (0.299) | 43.3502 | 22.8314 (<0.0001) | 3.4632 | 0.444 (<0.0001) | 0.0999 | 0.9337 | 0.0937 | 0.0591 | 0.8214 | |
| Gompertz | 2.3206 (<0.0001) | 0.1952 | 6.2032 (<0.0001) | 0.2834 | 0.0358 (<0.0001) | 0.0026 | 0.9299 | 0.0964 | 0.0649 | 4.8052 | |
| Chapman-Richards | 3.8923 (0.0001) | 0.9583 | 0.0175 (<0.0001) | 0.0040 | 2.9132 (<0.0001) | 0.2992 | 0.9328 | 0.0944 | 0.0613 | 1.7015 | |
| Weibull | 2.5051 (<0.0001) | 0.4821 | 0.00005 (<0.0001) | 0.00001 | 2.2834 (<0.0001) | 0.1106 | 0.9318 | 0.0951 | 0.0635 | 2.5439 | |
| Larix kaempferi (n = 313) | Logistic | 1.2817 (<0.0001) | 0.1067 | 0.1126 (<0.0001) | 0.0092 | 81.0024 (<0.0001) | 17.1789 | 0.7870 | 0.1495 | 0.0902 | 12.8173 |
| Korf | 21.2675 (0.5182) | 32.8801 | 25.1394 (0.0083) | 9.4589 | 0.5363 (0.0164) | 0.2222 | 0.7885 | 0.1489 | 0.0834 | 0.5679 | |
| Gompertz | 1.8400 (<0.0001) | 0.3157 | 6.7796 (<0.0001) | 0.7206 | 0.0475 (<0.0001) | 0.0068 | 0.7892 | 0.1487 | 0.0850 | 4.3490 | |
| Chapman-Richards | 2.5000 (0.0066) | 0.9134 | 0.0293 (0.0048) | 0.0103 | 3.5893 (<0.0001) | 0.7982 | 0.7891 | 0.1487 | 0.0835 | 1.2202 | |
| Weibull | 1.4885 (<0.0001) | 0.2979 | 0.00003 (0.1138) | 0.00002 | 2.6557 (<0.0001) | 0.2351 | 0.7899 | 0.1484 | 0.0837 | 1.9647 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, Y.; Lee, D.; Choi, J. Developing and Comparing Individual Tree Growth Models of Major Coniferous Species in South Korea Based on Stem Analysis Data. Forests 2023, 14, 115. https://doi.org/10.3390/f14010115
Seo Y, Lee D, Choi J. Developing and Comparing Individual Tree Growth Models of Major Coniferous Species in South Korea Based on Stem Analysis Data. Forests. 2023; 14(1):115. https://doi.org/10.3390/f14010115
Chicago/Turabian StyleSeo, Yeongwan, Daesung Lee, and Jungkee Choi. 2023. "Developing and Comparing Individual Tree Growth Models of Major Coniferous Species in South Korea Based on Stem Analysis Data" Forests 14, no. 1: 115. https://doi.org/10.3390/f14010115