
Aboveground Biomass Prediction of Plots in the Natural Forests of Arid Mountains Based on Large Trees

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Measurements
2.3. AGB Estimation from Field Plot Measurements
2.4. Analysis of the Relationship between AGB and Diameter at Breast Height
2.5. Construction of the Plot AGB Prediction Model Based on Large Trees
2.6. Optimal Number of Large Trees for Plot AGB Prediction
3. Results
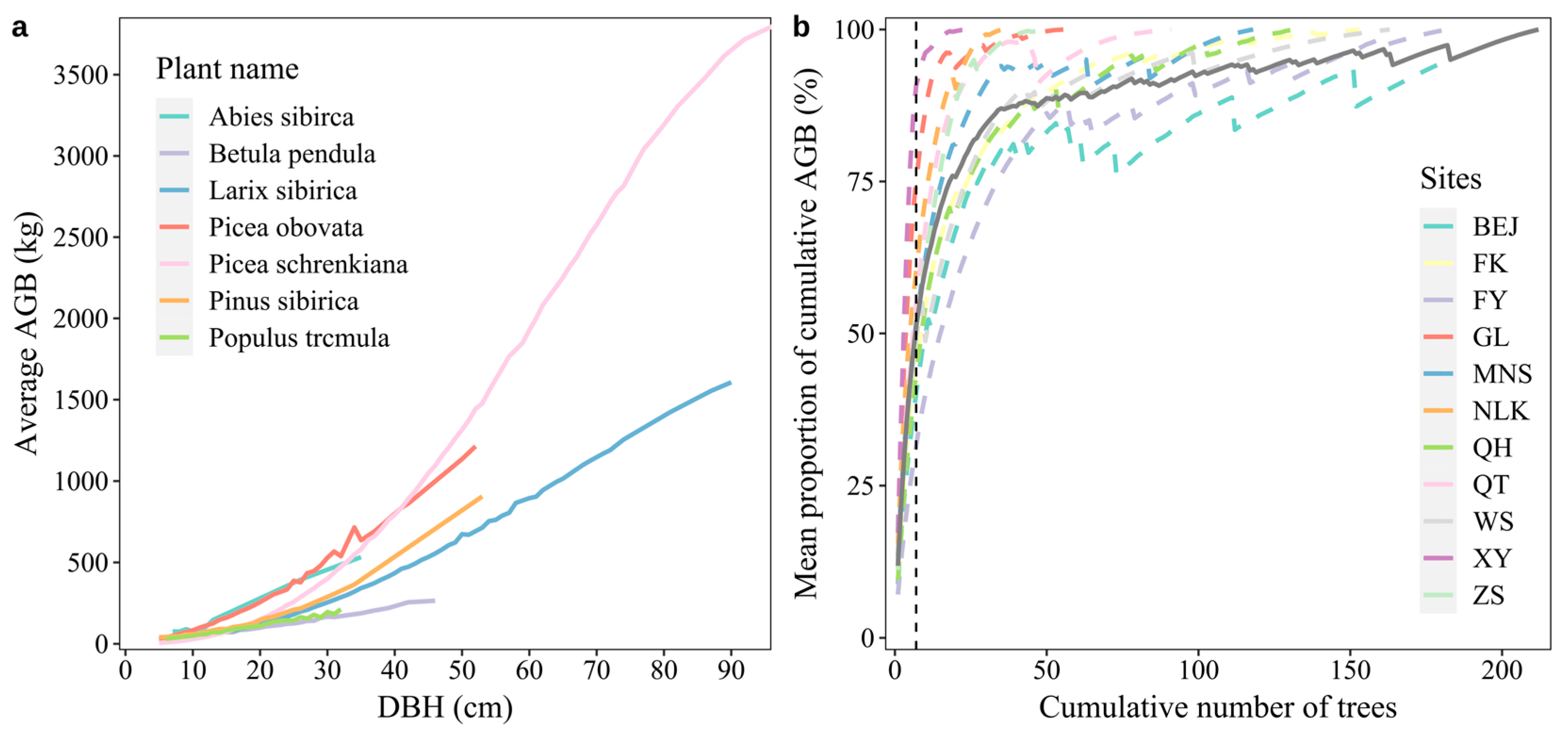
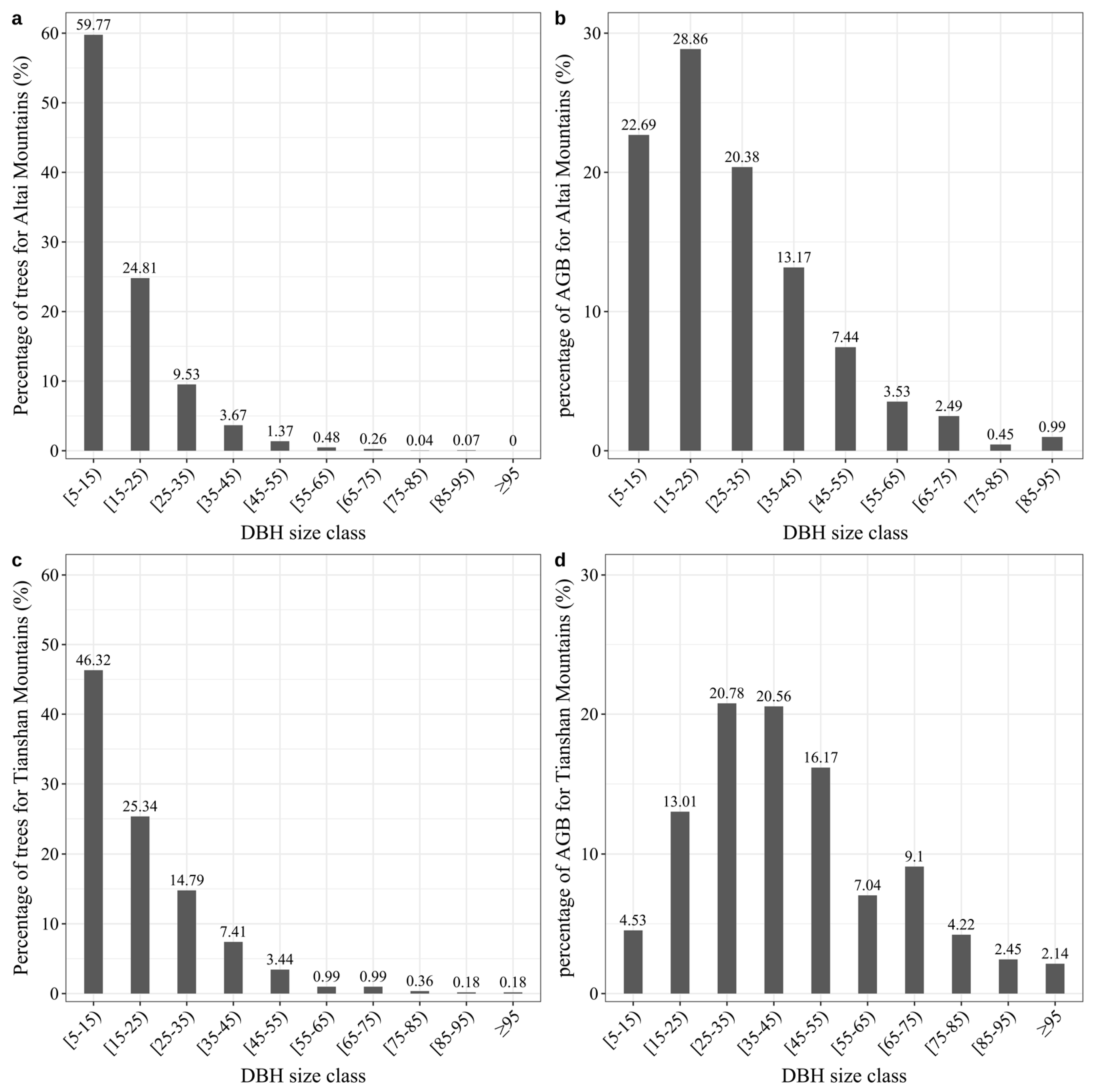
3.1. Contribution of Large Trees to AGB
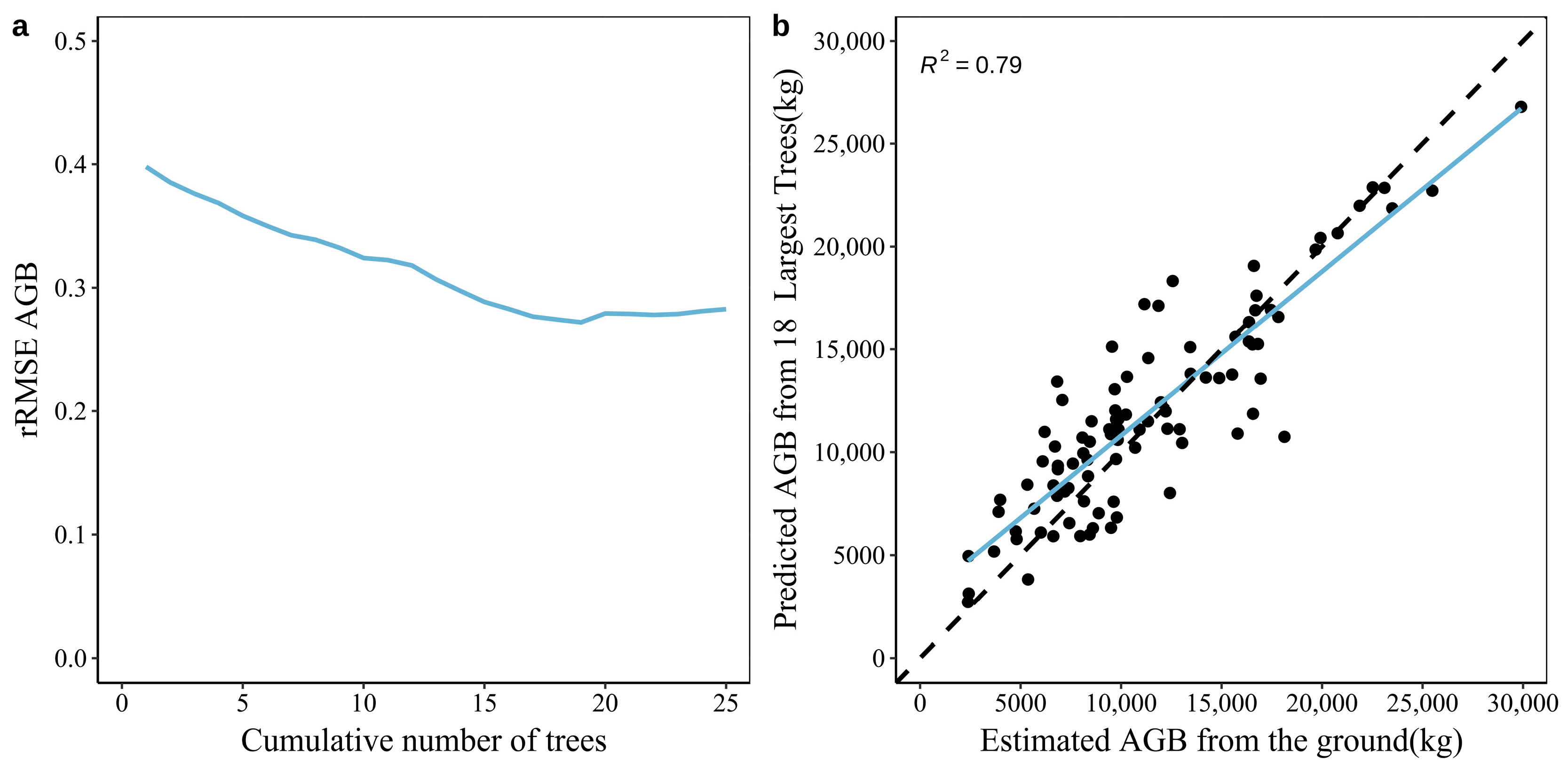
3.2. Construction of the AGB Prediction Model for The Plots of The Natural Forests of Arid Mountains Based on Large Trees
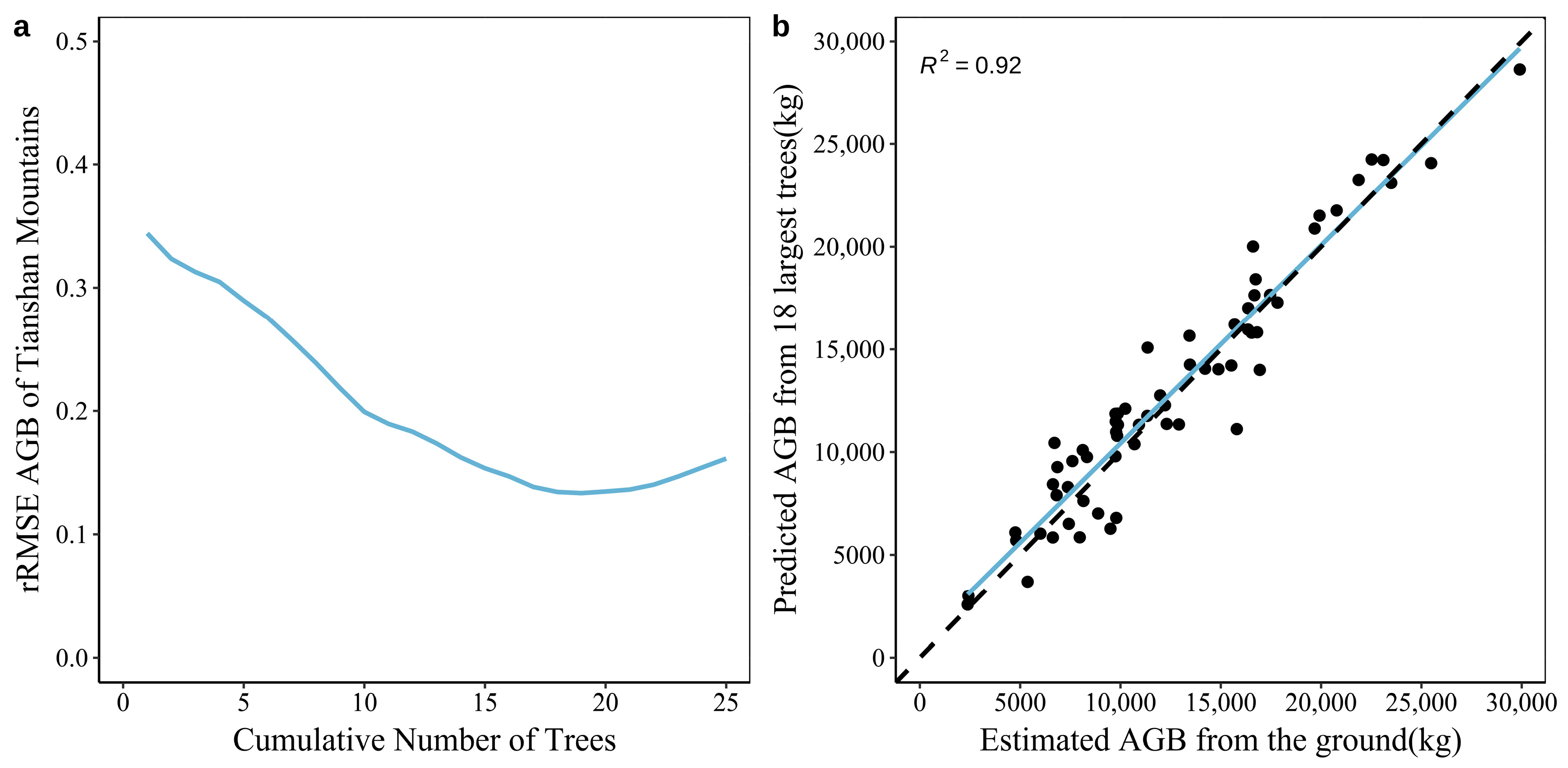
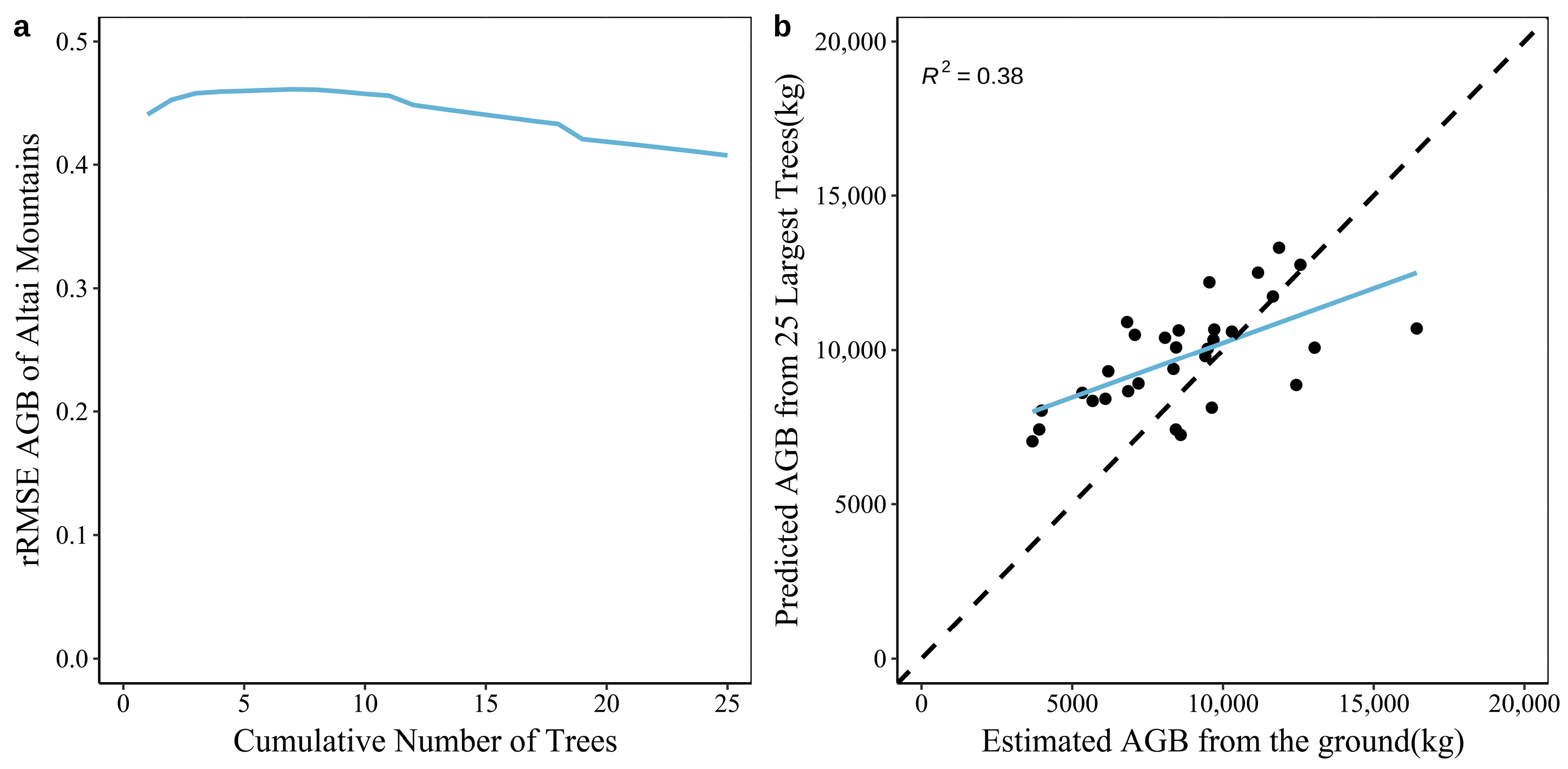
3.3. Construction of the AGB Prediction Model for Natural Forest Plots in the Tianshan and Altai Mountains Based on Large Trees
4. Discussion
4.1. Comparing Tianshan and Altai Mountains’ Natural Forests
4.2. Contrasting Arid Mountains and Tropical Forests
4.3. Significance of Large Trees in Arid Mountains’ Forests
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, S.; Lu, N.; Fu, B.; Wang, S.; Deng, L.; Wang, L. Current and Future Carbon Stocks of Natural Forests in China. For. Ecol. Manag. 2022, 511, 120137. [Google Scholar] [CrossRef]
- Tu, H.; Zhou, H.; Ma, G.; Zhang, R.; Yang, S. Characteristics of Forest Carbon Storage in Yunnan Based on the Ninth Forest Inventory Data. J. Northwest For. Univ. 2023, 38, 185–193. [Google Scholar]
- Guo, Z.; Huang, J.; Lu, X.; Wang, J.; Zang, R. Nature tree ecosystem services evaluation in Xinjiang based on the seventh national forest assessment data. Ecol. Sci. 2015, 34, 118–124. [Google Scholar] [CrossRef]
- Wang, X.; Wang, T.; Xu, J.; Shen, Z.; Yang, Y.; Chen, A.; Wang, S.; Liang, E.; Piao, S. Enhanced Habitat Loss of the Himalayan Endemic Flora Driven by Warming-Forced Upslope Tree Expansion. Nat. Ecol. Evol. 2022, 6, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Jian, N. Forest Productivity of the Altay and Tianshan Mountains in the Dryland, Northwestern China. For. Ecol. Manag. 2004, 202, 13–22. [Google Scholar] [CrossRef]
- Chen, Z.; Zan, M.; Yang, X.; Dong, Y. Prediction of forest vegetation carbon storage in Xinjiang. Ecol. Environ. Sci. 2023, 32, 226–234. [Google Scholar] [CrossRef]
- Alaniz, A.J.; Carvajal, M.A.; Marquet, P.A.; Vergara, P.M.; Meneses, L.; Moreira-Arce, D. Analyzing the Spatiotemporal Patterns of Forests Carbon Sink and Sources between 2000 and 2019. Earth’s Future 2022, 10, e2021EF002560. [Google Scholar] [CrossRef]
- Dai, L.; Zhang, Y.; Wang, L.; Zheng, S.; Xu, W. Assessment of Carbon Density in Natural Mountain Forest Ecosystems at Northwest China. Int. J. Environ. Res. Public Health 2021, 18, 2098. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, H.; Liu, L.; Huang, W.; Huete, A.R.; Zhang, X.; Wang, F.; Yu, L.; Xie, Q.; Wang, C.; et al. Estimating the Aboveground Biomass for Planted Forests Based on Stand Age and Environmental Variables. Remote Sens. 2019, 11, 2270. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rueger, N.; et al. Rate of Tree Carbon Accumulation Increases Continuously with Tree Size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef]
- Zhao, M.; Sun, M.; Xiong, T.; Tian, S.; Liu, S. On the Link between Tree Size and Ecosystem Carbon Sequestration Capacity across Continental Forests. Ecosphere 2022, 13, e4079. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Paoli, G.; McGuire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large Trees Drive Forest Aboveground Biomass Variation in Moist Lowland Forests across the Tropics. Glob. Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.-L.; He, J.-K.; Kong, F.-M.; Yu, J.-H.; Jiang, H.-S. Big-Sized Trees Overrule Remaining Trees’ Attributes and Species Richness as Determinants of Aboveground Biomass in Tropical Forests. Glob. Change Biol. 2019, 25, 2810–2824. [Google Scholar] [CrossRef] [PubMed]
- Mildrexler, D.J.; Berner, L.T.; Law, B.E.; Birdsey, R.A.; Moomaw, W.R. Large Trees Dominate Carbon Storage in Forests East of the Cascade Crest in the United States Pacific Northwest. Front. For. Glob. Change 2020, 3, 594274. [Google Scholar] [CrossRef]
- Yuan, Z.; Ali, A.; Sanaei, A.; Ruiz-Benito, P.; Jucker, T.; Fang, L.; Bai, E.; Ye, J.; Lin, F.; Fang, S.; et al. Few Large Trees, Rather than Plant Diversity and Composition, Drive the above-Ground Biomass Stock and Dynamics of Temperate Forests in Northeast China. For. Ecol. Manag. 2021, 481, 118698. [Google Scholar] [CrossRef]
- Lutz, J.A.; Furniss, T.J.; Johnson, D.J.; Davies, S.J.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.J.; Andrade, A.; Baltzer, J.; Becker, K.M.L.; et al. Global Importance of Large-Diameter Trees. Glob. Ecol. Biogeogr. 2018, 27, 849–864. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Barbier, N.; Réjou-Méchain, M.; Fayolle, A.; Gourlet-Fleury, S.; Maniatis, D.; de Haulleville, T.; Baya, F.; Beeckman, H.; Beina, D.; et al. Seeing Central African Forests through Their Largest Trees. Sci. Rep. 2015, 5, 13156. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Rutishauser, E.; Kellner, J.R.; Saatchi, S.; Pelissier, R.; Herault, B.; Slik, F.; Bogaert, J.; De Canniere, C.; Marshall, A.R.; et al. Pan-Tropical Prediction of Forest Structure from the Largest Trees. Glob. Ecol. Biogeogr. 2018, 27, 1366–1383. [Google Scholar] [CrossRef]
- Ali, A.; Wang, L.-Q. Big-Sized Trees and Forest Functioning: Current Knowledge and Future Perspectives. Ecol. Indic. 2021, 127, 107760. [Google Scholar] [CrossRef]
- Cheng, Y.; Lan, S.; Fan, X.; Tjahjadi, T.; Jin, S.; Cao, L. A Dual-Branch Weakly Supervised Learning Based Network for Accurate Mapping of Woody Vegetation from Remote Sensing Images. Int. J. Appl. Earth Obs. Geoinf. 2023, 124, 103499. [Google Scholar] [CrossRef]
- Lu, D.; Chen, Q.; Wang, G.; Liu, L.; Li, G.; Moran, E. A Survey of Remote Sensing-Based Aboveground Biomass Estimation Methods in Forest Ecosystems. Int. J. Digit. Earth 2016, 9, 63–105. [Google Scholar] [CrossRef]
- Gonzalez de Tanago, J.; Lau, A.; Bartholomeusm, H.; Herold, M.; Avitabile, V.; Raumonen, P.; Martius, C.; Goodman, R.; Disney, M.; Manuri, S.; et al. Estimation of Above-Ground Biomass of Large Tropical Trees with Terrestrial LiDAR. Methods Ecol. Evol. 2018, 9, 223–234. [Google Scholar] [CrossRef]
- Emin, M.; Anwar, E.; Liu, S.; Emin, B.; Mamut, M.; Abdukeram, A.; Liu, T. Target Detection-Based Tree Recognition in a Spruce Forest Area with a High Tree Density-Implications for Estimating Tree Numbers. Sustainability 2021, 13, 3279. [Google Scholar] [CrossRef]
- Jiang, J.; Zhu, W.; Qiao, K.; Jiang, Y. An Identification method for mountains coniferous in Tianshan with Sentinel-2 data. Remote Sens. Technol. Appl. 2021, 36, 847–856. [Google Scholar]
- Yang, B.-Y.; Ali, A.; Xu, M.-S.; Guan, M.-S.; Li, Y.; Zhang, X.-N.; He, X.-M.; Yang, X.-D. Large Plants Enhance Aboveground Biomass in Arid Natural Forest and Plantation along Differential Abiotic and Biotic Conditions. Front. Plant Sci. 2022, 13, 999793. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.; Murphy, H.T. The Importance of Large-Diameter Trees in the Wet Tropical Rainforests of Australia. PLoS ONE 2019, 14, e0208377. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Y.; Tang, Z.; Guo, Z.; Fang, J. Community structure and Its affecting factors of mountains coniferous forests in XinJinag China. Arid Zone Res. 2011, 28, 31–39. [Google Scholar] [CrossRef]
- Jiao, L.; Chen, K.; Liu, X.; Qi, C.; Xue, R. Comparison of the Response Stability of Siberian Larch to Climate Change in the Altai and Tianshan. Ecol. Indic. 2021, 128, 107823. [Google Scholar] [CrossRef]
- Wang, T.; Bao, A.; Xu, W.; Zheng, G.; Nzabarinda, V.; Yu, T.; Huang, X.; Long, G.; Naibi, S. Dynamics of Forest Net Primary Productivity Based on Tree Ring Reconstruction in the Tianshan Mountains. Ecol. Indic. 2023, 146, 109–713. [Google Scholar] [CrossRef]
- Xu, W.; Yang, L.; Chen, X.; Gao, Y.; Wang, L. Carbon storage, spatial distribution and the influence factors in Tianshan Mountains. Chin. J. Plant Ecol. 2016, 40, 364–373. [Google Scholar] [CrossRef]
- Zheng, S.; Xu, W.; Yang, L.; Gao, Y.; Li, J.; Wang, L. Carbon density and storage of forest ecosystem in Altay Mountains, Xinjiang. J. Nat. Resour. 2016, 31, 1553–1563. [Google Scholar]
- Bedane, G.A.; Feyisa, G.L.; Senbeta, F. Spatial Distribution of above Ground Carbon Density in Harana Forest, Ethiopia. Ecol. Process. 2022, 11, 4. [Google Scholar] [CrossRef]
- Li, W.; Wang, C.; Liu, H.; Wang, W.; Sun, R.; Li, M.; Shi, Y.; Zhu, D.; Du, W.; Ma, L.; et al. Fine Root Biomass and Morphology in a Temperate Forest Are Influenced More by Canopy Water Addition than by Canopy Nitrogen Addition. Front. Ecol. Evol. 2023, 11, 1132248. [Google Scholar] [CrossRef]
- Ali, F.; Khan, N.; Abd_Allah, E.F.; Ahmad, A. Species Diversity, Growing Stock Variables and Carbon Mitigation Potential in the Phytocoenosis of Monotheca Buxifolia Forests along Altitudinal Gradient across Pakistan. Appl. Sci. 2022, 12, 1292. [Google Scholar] [CrossRef]
- Dyola, N.; Sigdel, S.R.; Liang, E.; Babst, F.; Camarero, J.J.; Aryal, S.; Chettri, N.; Gao, S.; Lu, X.; Sun, J.; et al. Species Richness Is a Strong Driver of Forest Biomass along Broad Bioclimatic Gradients in the Himalayas. Ecosphere 2022, 13, e4107. [Google Scholar] [CrossRef]
- Tian, H.; Pei, J.; Huang, J.; Li, X.; Wang, J.; Zhou, B.; Qin, Y.; Wang, L. Garlic and Winter Wheat Identification Based on Active and Passive Satellite Imagery and the Google Earth Engine in Northern China. Remote Sens. 2020, 12, 3539. [Google Scholar] [CrossRef]
- Luo, Q.; Xu, Z.; Xu, Z.; Li, L.; Chang, Y.; Xu, X.; Song, X. Individual Biomass Allocation and Its Variation of Picea Schrenkiana Forests. Arid. Land Geogr. 2019, 42, 1378–1386. [Google Scholar]
- Hauck, M.; Csapek, G.; Dulamsuren, C. The Significance of Large Old Trees and Tree Cavities for Forest Carbon Estimates. For. Ecol. Manag. 2023, 546, 121319. [Google Scholar] [CrossRef]
- Piponiot, C.; Anderson-Teixeira, K.J.; Davies, S.J.; Allen, D.; Bourg, N.A.; Burslem, D.F.R.P.; Cárdenas, D.; Chang-Yang, C.; Chuyong, G.; Cordell, S.; et al. Distribution of Biomass Dynamics in Relation to Tree Size in Forests across the World. New Phytol. 2022, 234, 1664–1677. [Google Scholar] [CrossRef]
- Mensah, S.; Noulekoun, F.; Ago, E.E. Aboveground Tree Carbon Stocks in West African Semi-Arid Ecosystems: Dominance Patterns, Size Class Allocation and Structural Drivers. Glob. Ecol. Conserv. 2020, 24, e01331. [Google Scholar] [CrossRef]
- Kaushal, S.; Baishya, R. Stand Structure and Species Diversity Regulate Biomass Carbon Stock under Major Central Himalayan Forest Types of India. Ecol. Process 2021, 10, 14. [Google Scholar] [CrossRef]
- Birdsey, R.A.; DellaSala, D.A.; Walker, W.S.; Gorelik, S.R.; Rose, G.; Ramírez, C.E. Assessing Carbon Stocks and Accumulation Potential of Mature Forests and Larger Trees in U.S. Federal Lands. Front. For. Glob. Change 2023, 5, 1074508. [Google Scholar] [CrossRef]
- Teich, M.; Becker, K.M.L.; Raleigh, M.S.; Lutz, J.A. Large-Diameter Trees Affect Snow Duration in Post-Fire Old-Growth Forests. Ecohydrology 2022, 15, e2414. [Google Scholar] [CrossRef]
- Zuleta, D.; Arellano, G.; McMahon, S.M.; Aguilar, S.; Bunyavejchewin, S.; Castaño, N.; Chang-Yang, C.; Duque, A.; Mitre, D.; Nasardin, M.; et al. Damage to Living Trees Contributes to Almost Half of the Biomass Losses in Tropical Forests. Glob. Change Biol. 2023, 29, 3409–3420. [Google Scholar] [CrossRef]
- Bennett, A.C.; McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.J. Larger Trees Suffer Most during Drought in Forests Worldwide. Nat. Plants 2015, 1, 15139. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Number of Plots | Number of Trees | DBH (cm) | H (m) | |||
|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | ||||
| Altai Mountains | BEJ | 13 | 902 | 5.00–81.90 | 16.20 | 3.84–29.92 | 13.42 |
| FY | 12 | 1111 | 5.10–62.60 | 15.00 | 5.72–24.73 | 11.68 | |
| QH | 10 | 684 | 5.00–90.10 | 16.40 | 5.60–25.12 | 12.15 | |
| Tianshan Mountains | FK | 12 | 1015 | 5.00–65.50 | 15.30 | 5.60–29.92 | 10.91 |
| GL | 8 | 213 | 5.00–114.90 | 30.30 | 5.60–23.69 | 15.19 | |
| MNS | 16 | 779 | 5.00–78.30 | 20.20 | 5.60–23.67 | 12.90 | |
| NLK | 6 | 168 | 6.00–78.80 | 30.70 | 6.20–23.68 | 16.61 | |
| QT | 13 | 543 | 5.00–66.80 | 20.90 | 5.60–24.48 | 13.67 | |
| WS | 6 | 422 | 5.00–57.80 | 15.10 | 5.60–23.42 | 10.76 | |
| XY | 6 | 86 | 5.20–123.50 | 45.00 | 4.20–23.69 | 17.19 | |
| ZS | 3 | 121 | 6.90–66.40 | 26.40 | 6.73–23.68 | 15.72 | |
| Total | 11 | 105 | 6044 | 5.00–123.50 | 22.86 | 3.84–29.92 | 13.65 |
| Forest Type | Species of Trees | Allometric Growth Equation | R2 | Reference |
|---|---|---|---|---|
| Coniferous forest | Picea schrenkiana | WS = 0.0375(DBH2H)0.928 | 0.998 | [30] |
| WB = 0.0014(DBH2H)1.0972 | 0.998 | |||
| WL = 0.0117(DBH2H)0.8304 | 0.998 | |||
| Larix sibirica | WS = 0.099496(DBH2H)0.786530 | 0.990 | [8] | |
| WB = 0.098620(DBH2H)0.598367 | 0.990 | |||
| WL = 0.294136(DBH2H)0.357506 | 0.990 | |||
| Pinus sibirica | WS = 0.01278(DBH2H)0.99599 | 0.991 | [8] | |
| WB = 7.86722(DBH2H)0.18862 | 0.604 | |||
| WL = 3.32370(DBH2H)0.24569 | 0.658 | |||
| Picea obovata Abies sibirca | WS = 0.1283(DBH2H)0.7534 | 0.913 | [8] | |
| WB = 0.093(DBH2H)0.6732 | 0.913 | |||
| WL = 0.7753(DBH2H)0.5903 | 0.913 | |||
| Broad-leaved forest | Betula pendula Populus trcmula | WS = 0.6039(DBH2H)0.5325 | 0.959 | [8] |
| WB = 1.016(DBH2H)0.3922 | 0.957 | |||
| WL = 0.6989(DBH2H)0.2475 | 0.960 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, S.; Yi, L.; Bao, A.; Wang, Z.; Tao, Z.; Xu, W. Aboveground Biomass Prediction of Plots in the Natural Forests of Arid Mountains Based on Large Trees. Forests 2023, 14, 2426. https://doi.org/10.3390/f14122426
Xiong S, Yi L, Bao A, Wang Z, Tao Z, Xu W. Aboveground Biomass Prediction of Plots in the Natural Forests of Arid Mountains Based on Large Trees. Forests. 2023; 14(12):2426. https://doi.org/10.3390/f14122426
Chicago/Turabian StyleXiong, Shimei, Lubei Yi, Anming Bao, Zhengyu Wang, Zefu Tao, and Wenqiang Xu. 2023. "Aboveground Biomass Prediction of Plots in the Natural Forests of Arid Mountains Based on Large Trees" Forests 14, no. 12: 2426. https://doi.org/10.3390/f14122426
APA StyleXiong, S., Yi, L., Bao, A., Wang, Z., Tao, Z., & Xu, W. (2023). Aboveground Biomass Prediction of Plots in the Natural Forests of Arid Mountains Based on Large Trees. Forests, 14(12), 2426. https://doi.org/10.3390/f14122426