1. Introduction
Subalpine fir,
Abies lasiocarpa (Hook.) Nutt. (Pinales: Pinaceae), mortality has been increasing in British Columbia (B.C.) and throughout the Pacific Northwest in recent decades [
1,
2,
3,
4,
5]. This continuous and increased mortality is sometimes referred to as subalpine fir decline [
3,
4,
5] and was defined by Sinclair and Hudler [
6] as a premature progressive loss of vigor and health due to both biotic and abiotic influences. The primary cause of subalpine fir decline is the western balsam bark beetle,
Dryocoetes confusus Swaine (Coleoptera: Curculionidae: Scolytinae), which is considered the most destructive insect pest of subalpine fir in western North America [
1,
4,
7,
8,
9,
10,
11]. Compared to other native tree-killing bark beetle species, such as the mountain pine beetle,
Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae: Scolytinae) (MPB),
D. confusus has not been exhaustively studied. This is largely because subalpine fir (often referred to as “balsam”) is not a species of high commercial importance in the United States [
12], while in B.C. it has typically comprised an average of 9% of the total volume harvested annually since 2000 (R. Midgley, pers. comm.). However, as the salvage of MPB-killed pine from the 2000–2015 MPB outbreak concludes, harvest activity in B.C. is increasingly targeting non-pine forests, and there is a renewed interest and focus on the health and sustainability of subalpine fir forests [
13]. During the height of MPB salvage harvesting in the B.C. southern interior, “balsam” made up only 6% of the total volume (m
3) harvested compared to over 13% of the total volume (m
3) harvested between 2015 and 2022 (R. Midgley, pers. comm.). By contrast, in northern B.C., the volume of “balsam” harvested between 2016 and 2022 ranged from 12% to 24% (avg. = 18.6%) of the total volume harvested (R. Midgley, pers. comm.). Until recently [
1,
2,
3,
4,
5], there have been few studies that have quantified complex disturbance dynamics and impacts, such as those caused by
D. confusus and other damage agents that are accelerating the decline of these sensitive and previously remote subalpine fir forests.
Subalpine fir ecosystems are extremely valuable due to their inherent hydrologic contribution [
14], carbon sequestration [
15,
16], recreation opportunities, and habitat attributes. This species grows well at high elevations, from 600 to 2250 meters, throughout most of the B.C. interior [
17,
18,
19]. In the mountains and plateaus of interior B.C., subalpine fir is often associated with interior spruce,
Picea engelmannii x
glauca Parry ex Engelm. (Pinales: Pinaceae) and is a significant component of the interior high-elevation forests from the Yukon Territory to Arizona. Cool summers, cold winters, and a deep snowpack are important in determining where subalpine fir will grow well [
19]. Both spruce and subalpine fir are shade tolerant and able to persist in the understory of mature stands, taking advantage when disturbances create small gaps in the canopy [
20]. This study was conducted in southern B.C., in the Engelmann spruce—subalpine fir biogeoclimatic zone (ESSF), where subalpine fir is most common, often in mixtures with spruce and smaller components of other tree species [
17,
21,
22].
Canada is experiencing climate change at nearly twice the rate of the world’s average (Climate change adaptation in Canada (
nrcan.gc.ca)) (accessed on 1 December 2022), with drought and heat-induced tree mortality accelerating in many forest ecosystems globally. It is well documented that temperatures at high elevations and in the northern hemisphere have risen faster than the global average [
23,
24,
25]. Subalpine fir has a low tolerance to change, or extremes, in the summer heat and drought [
26,
27] and has consequently suffered increased climate-induced stress over the past few decades, with severe implications for tree health, resilience, and insect outbreaks. The radial growth of subalpine fir is declining and increasingly influenced by summer conditions in the previous year before growth, indicating shifting adaptive responses to higher summer temperatures and moisture deficits in B.C. over the past eight decades [
27].
Dryocoetes confusus is likely adapting to this declining growth and increased susceptibility in trees [
10], coupled with more favorable summer temperatures for flight and reproduction. The result is that subalpine fir mortality in recent decades has far exceeded the expected rate of natural mortality [
2]. Increasing drought events are believed to be one of the underlying causes of forest mortality and decline in western North America, altering forest structure, composition, ecosystem function, and overall forest health [
15,
27,
28,
29].
Biotic and abiotic disturbances are the main drivers of natural forest succession, yet some, such as bark beetles [
16,
30], can cause excessive damage, even though they are integral to healthy and functioning ecosystems [
31]. Bark beetle outbreaks can have long-lasting impacts on forest ecosystems, ranging from decreased biodiversity to a reduction in carbon storage or the disruption of timber supply, which can have cascading effects on regional and global economies [
32]. British Columbia has experienced extensive, catastrophic escalations in tree mortality from bark beetles, most notably from the 2000–2015 MPB event that significantly altered forest landscapes [
33,
34,
35,
36,
37,
38]. Massive abiotic disturbances have also affected B.C., including drought and heat events in 1998 and 2017, which affected more than 10,035 and 119,000 hectares, respectively [
39,
40], and large wildfires, notably in 2017, 2018, and 2021 (Wildfire Averages—Province of British Columbia (
gov.bc.ca)) (accessed on 1 December 2022) which impacted over 1.3 million hectares of forest.
Dryocoetes confusus causes significant cumulative mortality [
2] over vast areas of high elevation and northern forests [
4,
5,
7,
8,
9,
11] with continuous pulses of low-level, spatially aggregated attack. This bark beetle becomes noticeable in stands aged 100 years or older [
2] and presumably attacks younger trees that are susceptible. As stands age and grow, the clumped pattern of attack creates small-scale gaps, lowers the overall stand density, and can allow the release of understory seedlings [
41]).
Dryocoetes confusus attacks the largest diameter standing live trees in a stand, although the mean diameter of the trees attacked can vary widely among stands [
10]. Recent windthrow can also be colonized, but this is a relatively rare resource for the beetle, and development seems to be more prolonged in downed trees compared to standing trees [
42,
43]. The relative scarcity of fresh windthrow as a host has most likely shaped the evolution of the beetles’ highly selective process of finding and colonizing live but slower growing [
10,
43] and often severely stressed host trees.
Dryocoetes confusus typically displays a two-year life cycle with new adults emerging from the trees in late June through July [
12,
42,
44,
45] and again later in the season from August to September, when a much smaller, secondary flight comprised these same parent beetles can occur [
12,
42,
46,
47,
48,
49].
Dryocoetes confusus is closely associated with the pathogenic fungus
Grosmannia dryocoetidis (Ophiostomatales: Ophiostomataceae) [
50], which is transmitted to the colonized trees, and this beetle–fungus complex can overcome the tree’s defenses. Adult beetles are active under the bark while snow remains on the ground, often weeks before subcortical temperatures trigger emergence and flight [
42]. This initial attack on the trees, and the early season extension of galleries by overwintered females under the bark, produces frass and fine sawdust on tree boles outside entrance holes. The presence of long resin streams on the bole, without the presence of frass, indicates an unsuccessful attack by the beetle. The brood overwinters, once as larvae and again as new adults, before emerging in the third summer to repeat the cycle. Females sometimes lay two broods within the same host over a period of two summers.
Dryocoetes confusus preferentially attack the slower-growing trees in a forest stand [
10], likely due to reduced primary defenses [
51] in these trees. As many subalpine fir forests experience further protracted periods of warm weather in spring and summer,
D. confusus could potentially shorten its life history to one year, similar to that observed with the spruce beetle,
Dendroctonus rufipennis Kirby (Coleoptera: Curculionidae: Scolytinae) [
52,
53,
54].
Dryocoetes confusus likely plays a critical role in forest succession. Infestations show a degree of aggregation [
5,
41,
55,
56], which could depend on the occurrence pattern of susceptible trees in stands [
10,
57]. The distribution of susceptible hosts may affect natural regeneration and potential stand management strategies, such as salvage harvesting. Disturbance factors create growing spaces that are gradually re-occupied by regeneration. Historically, this continual production and occupancy of small gaps have been a slow process, fundamental in self-perpetuating climax old-growth forests where the general overstory canopy remains. With increasing biotic and abiotic stress being placed on these stands, this slow successional process may be changing and accelerating the impacts on forest values, stand productivity, and in-stand mortality.
This long-term study aims to describe and quantify the factors that drive subalpine fir mortality and the role of D. confusus in natural succession and in shaping mature subalpine fir forests in southern B.C. We wanted to test whether our initial designation of stands into an “outbreak phase” of D. confusus was reasonable. Knowing the attributes reflected in an early-, mid-, or late-phase stand could enable managers to have greater insight into future stand conditions with potential management options. We established one-hectare permanent sample plots in ESSF stands to study the attack progress and patterns of D. confusus and its influence on the mortality of subalpine fir and stand succession by elucidating:
Stand and tree attributes of early-, mid-, and late-phase stands impacted by D. confusus;
The progression of D. confusus attack and subalpine fir mortality in stands;
The spatial-temporal distribution of live subalpine fir, other tree species, and D. confusus attacked subalpine fir.
This long-term, multi-faceted study will provide a better understanding of how mortality from D. confusus and other factors shape subalpine fir forests. This study is unique in that it follows individual trees from their death (attack) to down and monitors insect development within the trees. It also elucidates the spatial parameters that result from a long-term disturbance in subalpine fir forests. Results from this study may provide a pathway to address future the management of these valuable but delicate ecosystems.
4. Discussion and Conclusions
Dryocoetes confusus is integral to the successional processes in subalpine forests and has likely been the main disturbance agent for millennia. It is well adapted to its host and the harsh climatic conditions where both are found. Historically, succession in subalpine fir-dominated forests is a slow process not typically driven by stand replacement fires [
55] but rather due to the patchy, intermittent mortality caused by
D. confusus and other factors, which create small gaps and windthrow, thereby increasing the ground disturbance necessary for the promotion of new seedlings. This slow yet continuous turnover of mature subalpine fir maintains these forests in a gap-dynamic driven and functioning state. However, the accelerated rate of mortality observed in the past two decades is causing an imbalance in this delicate beetle-driven dynamic process. Our study represents just a small footprint within the vast area occupied by subalpine fir forests in B.C. However, we have shown significant mortality and disturbance occurring in these stands. If this is a correct reflection of even a portion of the landscape, then the successional progress, or decline, of these forests is becoming unbalanced.
The successional or outbreak phase of a mature subalpine fir stand can reasonably be estimated by considering stand density, tree mortality, recent
D. confusus attacks, and stand deterioration (dead from other causes and windthrow).
Dryocoetes confusus was the major disturbance in our study, causing over two-thirds of the total subalpine fir mortality observed. Other biotic and abiotic factors played a role in the general decline by killing subdominant trees in stands or possibly predisposing trees to attack by
D. confusus. We found stand density to be a strong predisposing factor in the decline of subalpine fir, with smaller diameter stems dying from a combination of abiotic stressors and secondary biotic factors, including drought,
Armillaria spp., and less aggressive bark beetles. Others [
4,
11] support this strong relationship between subalpine fir density, basal area (volume), and increasing
D. confusus attacks and suggest that it may take more than 100 years to return these heavily impacted stands to pre-outbreak conditions. With amplified heat and moisture stress affecting subalpine fir ecosystems, the recovery time may be much longer, and a return to pre-outbreak conditions may not be possible. A higher stem density creates more below- and above-ground competition for water, nutrients, and growing space. As the carrying capacity of a stand is reached, commonly described as the basal area or volume, growth is reduced, and trees die [
81]. This leads to senescence and a lowered capacity to defend from cortical invaders [
10,
16]. Our study confirms this successional strategy, with
D. confusus activity being very high in early outbreak phase plots and then declining as the cumulative stem mortality creates significantly lower live densities and volumes, moving stands into mid-to-late outbreak phases.
Dryocoetes confusus searches for and attacks the largest and presumably slowest-growing trees in a stand until the size and density of the remaining live hosts became sub-optimal. Even though there may be subalpine fir of an acceptable size (>20 cm) remaining in the stands when sph is reduced to below 400, the attacks decline. As the live component of subalpine fir declines, the remaining live trees may take advantage of this increased growing space by resuming radial growth and, in turn, become less susceptible to
D. confusus attack.
Dryocoetes confusus seems to have a wide and very finely tuned flight and search pattern for finding the most acceptable and vulnerable hosts. There was minimal change in the average size of live subalpine fir from the time of plot establishment to the final assessment due to selective thinning by
D. confusus and other damage agents.
A common host-selection characteristic of tree-killing bark beetles is to select large-diameter trees that provide a more suitable habitat for attacking beetles and developing offspring. Additionally, it is generally accepted that weaker or stressed trees are more susceptible to bark beetle attacks due to reduced primary defenses [
51,
82].
D. confusus displayed this same preference in our study by infesting all the largest trees, which were presumably slower growing [
10], until host size, density, and distribution were sub-optimal. Even though
D. confusus is most active in older stands [
2], there was no correlation between tree age and infestation phase, suggesting that
D. confusus is ubiquitous in mature stands and other tree and stand parameters are more important in the beetles’ host selection.
In the early phase of infestation progression in the stands, we found evidence that
D. confusus may be able to shorten its development time to one year. This may, in part, explain the rapid increase in attack rates observed in some plots. Bright [
52] and Negrón and Popp [
12] have also considered this possibility suggesting that extended warmer spring and summer temperatures may trigger earlier adult emergence or benefit accelerated development the year following oviposition. Our study revealed the presence of numerous nuptial chambers in attacked trees containing one male with multiple females but no brood galleries when sampling in late summer. This may be the jump-start needed to trigger a one-year life cycle.
Bark beetles and other pathogens create disturbances across diverse forest ecosystems resulting in a disruption to stand structure complexity, species composition, and even the function of these forested landscapes [
83,
84,
85]. Our study suggests that
D. confusus is altering subalpine fir landscapes in tandem with, and influenced by, the amplification of anthropogenic and climate disturbance. Horizontal and vertical structural heterogeneity influences various forest attributes (tree growth, plant species diversity, and insect and wildlife habitat), which, in turn, influences a forest’s resilience to disturbance [
86,
87,
88,
89]. The increasing mortality and windthrow have changed the area available (growing space) to trees over time, suggesting that “openings” were created in the overstory canopy, which, in turn, may contribute to gap structure and conditions that are suitable for regeneration [
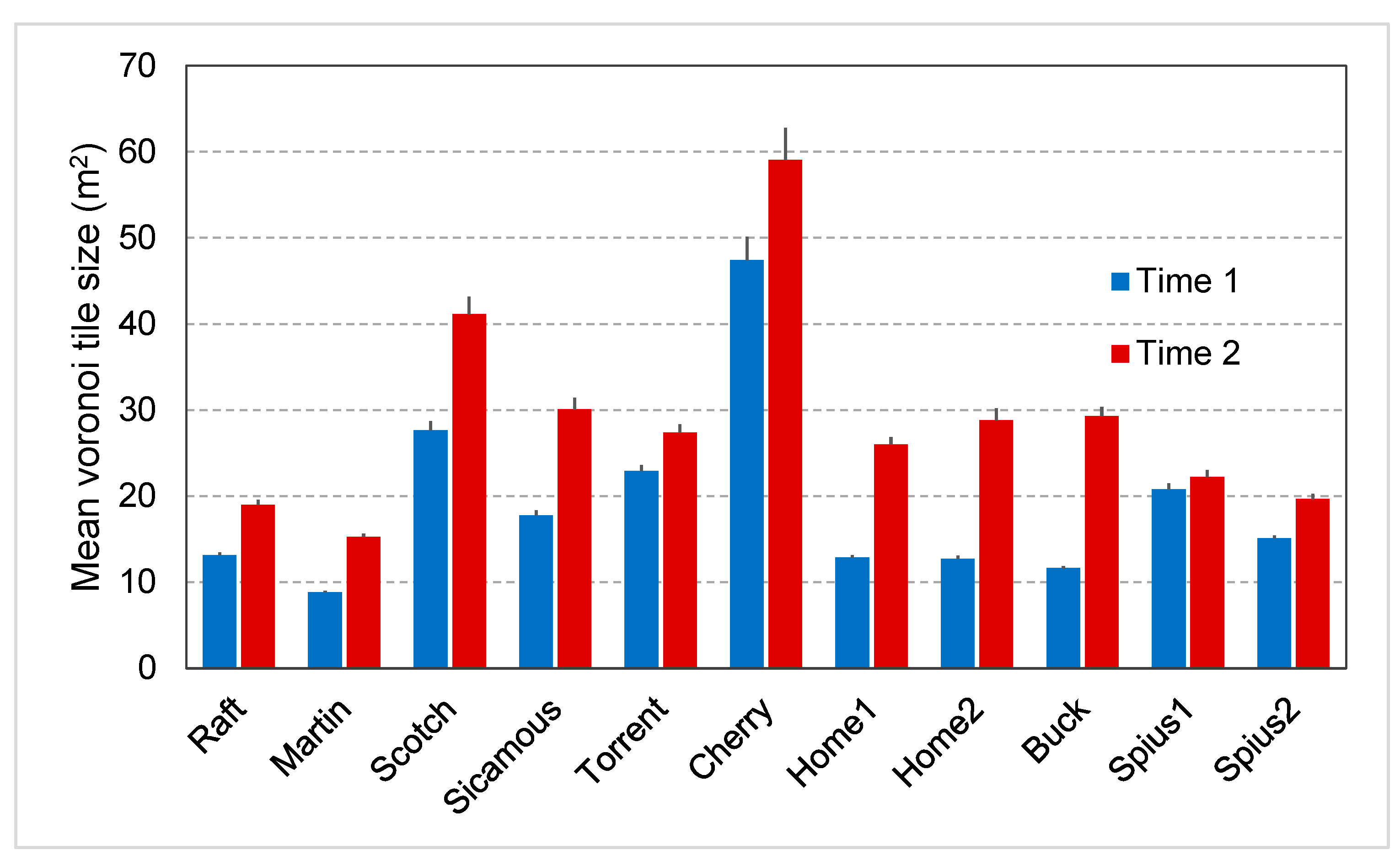
89]. Assuming that the crown diameter of a subalpine fir or spruce is about 4 m [
90,
91], the “occupied canopy area” would therefore be about 13 m
2. The average area potentially available to a tree changed from 11 m
2 at its establishment to 18 m
2 by the final assessment, suggesting that canopy openings were more than large enough to support the release of understory trees and perhaps trees located on the “edges of gaps” [
92]. Subalpine fir cannot compete successfully with commonly associated tree species (spruces, lodgepole pine, Douglas-fir,
Pseudotsuga menziesii var. glauca, (Mirb.) Franco (Pinales: Pinaceae)) if light intensity exceeds 50% of the full shade [
93].
In a cluster analysis of mixed conifer dry ponderosa pine forests, Larson and Churchill [
73] found that at least 6% of clusters were single trees at inter-tree distances of ≥7 m. In a similar approach, Sanchez Meador et al. [
94] found that trees were all members of the same patch when limiting inter-tree distance to an average of 12 m. Churchill et al. [
64] and Abella and Denton [
76] used an “average” crown diameter of 6 m (3 m radius) when assessing reference conditions for dry mixed conifer forests in the southwest USA. Mature subalpine fir and spruce both have long narrow, compact conical crowns and, in interior B.C., a crown radius of about 2 m [
26,
90,
91,
93]. In theory, an empty space distance with a radius of 3 m is big enough for a mature subalpine fir or spruce tree with a crown radius of 2 m but not big enough to accommodate two or more trees. Even so, in our study, there were few openings larger than a 5 m radius at either time and even though these gaps became sizable with increasing mortality, their shape could hinder future or rapid occupancy. Seedlings respond to the creation of canopy gaps, but recruitment into the overstory may not happen for a long time if they are too close to established overstory trees [
73].
The ecological integrity and economic value of subalpine fir ecosystems are changing. Stands are transitioning rapidly from early-phase to late-phase outbreak status, with a high proportion of the subalpine fir component standing dead or down. In less than a decade,
D. confusus-attacked trees have deteriorated in quality transitioning from live to various stages of decay, thereby reducing recoverable harvest volumes and leaving fewer options for rehabilitation. Dead-standing trees remain important for their hydrologic contribution [
14], carbon sequestration [
15], and numerous other ecological processes and use. Still, these too, may be compromised if the ratio of live-to-dead trees becomes unbalanced.
Although subalpine fir forests are susceptible to a range of disturbances, including fire [
55,
95,
96], historically, stand-replacement fire intervals have been very long, so small gap dynamics appear to have been the main successional and ecological driver in these ecosystems until very recently. The current accelerated decline of subalpine firs in conjunction with drier and hotter summer conditions may change future fire dynamics by providing more continuous and connected areas of dead and down fuel for fires should fire-starts occur. Therefore, forest managers must take a landscape-level view of these valuable but vulnerable and declining forests and determine where, when, and how much intervention is necessary to protect and conserve ecological functions. Our study has provided a tool for classifying stands into early-, mid-, and late-phase in the outbreak progression of
D. confusus. By applying this tool across susceptible landscapes, managers can better understand current losses and projected losses in the next decade. To maximize economic return, harvest and reforestation efforts should focus on early-phase stands. Conversely, the rehabilitation of late-phase stands, including harvest and prompt reforestation, may be of a higher priority. Further research is needed to determine whether
D. confusus can shift to a one-year life cycle and model how accelerated development rates may affect stand-level population expansions in early-to-mid-phase outbreak scenarios. A conscious application of proactive strategies on the landscape is necessary to prolong the viability of high-elevation and northern subalpine fir forests.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}