Abstract
As an attempt to examine the causes of forest disturbance and degradation of the orchid community, a comparative study on diversity and ecology in eight undisturbed and ten disturbed forests in Peninsular Malaysia was conducted that varied in areas, elevations, vegetation types, and disturbance regimes. Density and individual-based rarefaction curves were used to describe the abundance. Univariate and multivariate analyses were also performed to explore the associations of species abundance with biotic and abiotic factors. The study reported 239 orchid species belonging to 65 genera. Species richness, abundance, density, and diversity of orchids varied by locality. Higher density of orchids (2.433 plants/km2) occurred in the undisturbed forests than in the disturbed forests (0.228 plants/km2). As with the character of undisturbed forests, the temperature was between 27.8 ± 0.3 °C and 31.2 ± 0.2 °C, humid (77.1 ± 1.2%–89.6 ± 0.9%), and with low light intensity (23.8 ± 3.3 μmol m−2s−1–171.7 ± 18.8 μmol m−2s−1), thus supporting the high density of the plants. Disturbed forests had higher diversity (H = 4.934 and 1-D = 0.990) and abundance (183 species of 57 genera) but were determined to be highly influenced by the higher abundance of epiphytic orchids on the fallen trees and ease of accessibility in the logged forests. Terrestrial and mycoheterotroph orchids were much lower in density and abundance in the disturbed habitat indicating a gradual reduction in their niche availability following the disturbance. Additionally, the ecology data show that the microclimate conditions of the canopy-covered forest was influenced by proximity to the logged area which had eventually reduced the orchids’ habitat quality. Furthermore, the results show that the abundance of epiphytic orchid communities was associated with the host plant characteristics. Host types and bark texture preference were apparent for the epiphytic orchid species, with certain types and textures hosting more orchid species than others. Overall results show that extreme temperature, humidity, and light intensity caused by the canopy opening inflicted damages to the habitat conditions and bark textures of the host plants and limits recolonisation of the orchids in the disturbed forests. The species diversity and density patterns of orchids in undisturbed and disturbed forests revealed in this study provide a baseline for conservationists, policy makers, and forest authorities in expanding the understanding of the forest ecology and vegetation along the disturbance gradient, forest regeneration, and criteria for plant selection for forest restoration in Peninsular Malaysia.
1. Introduction
Orchids are diverse and a very good group for studying diversity, ecology, distribution, and host-species associations in undisturbed and disturbed habitats [1]. The diversity study focused on the orchids occurring in Peninsular Malaysia, where previously a total of 982 orchid species were recorded and most of them were epiphytes (personal observation). Apart from focusing on their diversity, ecology, distribution patterns, and conservation, this paper explores the associations between diversity and distribution of epiphytic orchids and host characteristics in different vegetation types of Peninsular Malaysia. Although the association between epiphyte diversity and different host characteristics are well studied for plants, very little is known about variation in the abundance of epiphytic orchids and their host associations in tropical rainforests. Host plant characteristics play an important role in colonisation of host-dependent species [2,3,4]. In studies of relationship between tree size and epiphyte species richness, the host’s size matters due to greater bark surface available for colonisation on large trees and the creation of additional microhabitats [5,6]. Host position and type, and growth of epiphytic orchid species are influenced by microclimate conditions, including annual precipitation, light intensity, and humidity [3,7]. Conservation-wise, the degree of orchids’ host specificity is important to look at, because being host specialists, wild orchids are threatened by co-extinction with their hosts due to habitat alterations and climate change [8,9,10].
Lack of monitoring programs assessing the before or aftereffects of disturbances in Malaysian rainforests is one of the major limitations to quantifying the commonalities and differences of disturbance effects on forest ecosystem and its inhabitants. Epiphytes are a diverse group with very interesting niche ecology [11,12,13]. Unlike terrestrial species, epiphytes are highly reliant on their hosts and are thus likely to be affected by host species and host traits [14,15,16,17]. Notwithstanding their economical and conservation importance, the understanding of the wild orchid diversity and ecology under wide range of microclimate conditions in the undisturbed and disturbed forests is still rather poor, especially when the impacts come under different disturbance regimes. A comparative study of different undisturbed and disturbed forest types including scoring for different agents of disturbance will give unbiased and general views on the effect of forest and ecological disturbance on the orchid community. Additionally, if only protected areas are targeted, more wild orchids will be lost, and these species could disappear from large areas of former range. Thus, the present study includes several forest types, vegetation types, and disturbance regimes. This diversity and ecology study determines the suitability of the plants, especially trees as protectors and homes to epiphytic plants along with threats and limiting factors.
Specifically, in this paper, the following questions were addressed: (1) Does the diversity of orchids in undisturbed and disturbed forests of Peninsular Malaysia differ? (2) What are the factors determining diversity and composition of orchids in the undisturbed and disturbed forests? (3) What is the significance of ecological alterations on the orchid community in two adjacent forests that are located next to each other or connected to each other, but with distinct levels of disturbance? (4) How much do the number of host plants and host characteristics influence the number of epiphytic orchid species?
2. Materials and Methods
2.1. Study Area
Samplings were conducted between November 2016 and April 2021 in 18 different localities in Peninsular Malaysia (Figure 1). The systematic sampling involved choosing exploration sites in Peninsular Malaysia. We collected information on each of the site conditions (e.g., area, vegetation type, elevation, disturbance regime, and disturbance origin) (Table 1 and Table 2). Major disturbances were classified as either natural or anthropogenic in origin. To disentangle the effects of anthropogenic and natural disturbance regimes, there were two main forest types varying in vegetation: undisturbed (Table 1) and disturbed (Table 2) forests, with several subsidiary study sites, eight undisturbed and ten disturbed forests, were selected. The sub-classified disturbed forest types were defined according to the vegetation, disturbance, and period of disturbance. Classification of the vegetation types was based on [18] (modified from [19], and [20] for Bukit Pedu, Kedah. These sites represent some of the major ecosystem and vegetation types in Peninsular Malaysia. To characterise and compare disturbance regimes amongst disturbed forest types, the Disturbance Index (DI) based on five variables (Severity (D), Extension (E), Frequency (F), Duration (D), and Patch Type (PT)) were evaluated following a scoring method and ordinal variables modified from [21] (Table 3). The DI was calculated by multiplying the assigned scores describing the five disturbance variables: DI = S × E × F × D × P × T. A low DI indicates a low impact of a given disturbance agent for a particular ecosystem, whereas a high DI indicates a large impact.
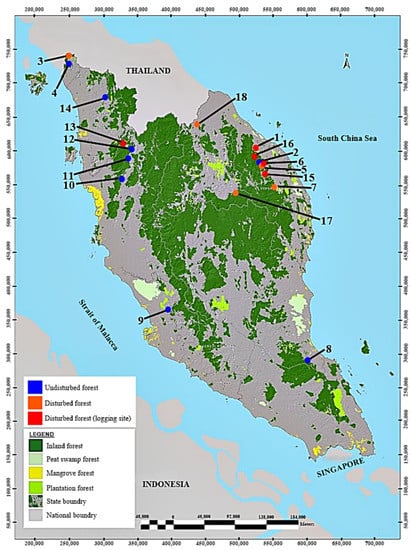
Figure 1.
Map showing the study localities (modified and reproduced with permission from JPSM (2019)): 1: Hulu Setiu, logged lowland dipterocarp forest (LHS), 2: Bukit Batu Kapal, lowland riparian forest (BK), 3: Gunung Perlis, hill dipterocarp forest (GP), 4: Bukit Rongkit, hill limestone forest (BR), 5: Bukit Batu Kapal, forest margin of a mixed dipterocarp forest (BK(M)), 6: Bukit Batu Kapal, logged hill dipterocarp forest (LBK), 7: Bukit Sekayu, lowland riparian forest (BS), 8: Padang 7, hill mixed dipterocarp forest (PG7), 9: Taman Rimba Komanwel, lowland dipterocarp forest (TRK), 10: Lata Kekabu, lowland riparian forest (KB), 11: Lata Lawin, lowland riparian forest (LW), 12: Lata Tanjung Kala, lowland riparian forest (TK), 13: Bintang Hijau, logged lowland dipterocarp forest (LB), 14: Bukit Pedu, conglomerate hill forest (BP), 15: Gawi, logged lowland dipterocarp forest (LG), 16: Petuang, logged lowland dipterocarp forest (LP), 17: Kuala Koh, lowland dipterocarp forest (KH), 18: Tanah Merah, lowland forest (TM). Source: Jabatan Perhutanan Semenanjung Malaysia (JPSM).

Table 1.
General characteristics of undisturbed forests in Peninsular Malaysia considered in this study.
Table 2.
General characteristics of disturbed forests in Peninsular Malaysia considered in this study.
Table 3.
Ordinal variables assessed to characterise and compare disturbance regimes amongst the disturbed studied sites.
2.2. Sampling Design, Samples, and Data Collection
Samplings were carried along the existing and off-road four-wheel trails using the line-transect method designed at accessible trail(s) in the selected study sites and established randomly by selecting a point along the trail in each locality. To accommodate sampling in the large areas such as logging sites and skid trails, the study areas were calculated based on the distance travelled of each trail multiplied by the width of 5–10 metres on each side of the trail, as shown in Figure 2. Observers walk along lines and count all orchid plants observed within the specified width. The length, width, and number of transect plots were not fixed considering the topography, accessibility, time constraint, movement restrictions due to the pandemic, and budget. Due to the limitations, the comparative study focused on different forest types irrespective of the total size area. Square kilometre (km2) is used as the unit for total transect plots’ area for ease of density calculation, graph scaling, and plotting.
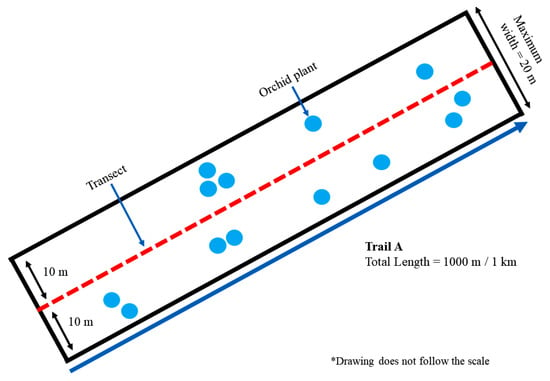
Figure 2.
An example of a line-transect (1000 m × 20 m transect plot). The total transect plots’ area was converted to the unit of square kilometre (km2).
One individual of the flowering specimens was collected and all occurrence pertaining to the same species were successively pooled for each transect and trail. Samples with floral structures were preserved using a standard herbarium technique [22] and voucher specimens were deposited in the Herbarium of Universiti Putra Malaysia (UPM) at the Department of Biology, Faculty of Science. The robust non-flowering individuals were brought back to UPM and transplanted into the ex situ conservatory as living collections. However, some of the non-flowering individuals that are also fragile, minute size, and found growing in few populations were not collected during the first visit until flowering. The flowering individuals were then collected during the revisits. Additionally, some sterile individuals were simply determined at generic level or to the closest affinity on the basis of their vegetative morphological characters. The exact locality is withheld to protect the population from illegal collections.
For species identification and evaluation of each species’ distribution status, reliable taxonomic and floristic literatures were referred and studied, e.g., [23,24,25,26]. Digitalised images of herbarium collections, botanical drawings, and records deposited in the National Herbarium of the Netherlands (NHN) accessed through Browse Dutch Natural History Collections: BioPortal (Naturalis) (http://bioportal.naturalis.nl/ (accessed on 27 October 2022)), Herbarium of Singapore Botanic Gardens (SING) accessed through BRAHMS Online managed by the University of Oxford (http://herbaria.plants.ox.ac.uk/bol/sing (accessed on 27 October 2022)), Swiss Orchid Foundation [https://orchid.unibas.ch/index.php/en/ (accessed on 27 October 2022)), Kew Herbarium Catalogue (http://apps.kew.org/herbcat/gotoSearchPage.do (accessed on 27 October 2022)), Natural History Museum Specimen Collection (https://data.nhm.ac.uk/ (accessed on 27 October 2022)), Herbarium of Aarhus University (AAU)(https://www.aubot.dk/search_form.php (accessed on 27 October 2022)), Museum National D’Histoire Naturelle (MNHN) (https://science.mnhn.fr/all/search (accessed on 27 October 2022)), and Plants of the World Online (POWO) (http://www.plantsoftheworldonline.org/ (accessed on 27 October 2022)) were examined prior to the taxonomic treatment and assessment on the range of distribution for each species. The accepted names were validated via KEW World Checklist of Selected Plant Families (WCSP) [27] and POWO.
Occurrence of the host plants containing epiphytic orchids was recorded within the studied transect plots. For each sampled host plant, parameters, and characteristics: height, bark’s texture, canopy’s structure, crown’s forms and classes, and substrate cover were recorded in a spreadsheet. Height was measured either using clinometer and visual interpretation or measuring tape for fallen trees. Each character was described and photographed on the field. The presence of epiphytic orchids was recorded for the entire host plants. The host species and associated epiphytic orchids were mostly identified in the field and verified using different local floras, e.g., [28,29,30,31,32,33,34,35,36,37,38]. The unidentified species of host plants were photographed in detail along with their flowers, leaves, and fruits (if present) and later identified with expert consultation. Studying epiphyte diversity is challenging because it is difficult to observe and identify the plants in the tree canopy from the ground [39]. Each host plant was examined from the ground at different points, thereby assuring a clear view of all the orchids and host’s parts [4,40]. Hence, for large and tall host plants, the inaccessible epiphyte growing up on it were only photographed and captured using a Nikon D5100 and a macro lens AF-S DX NIKKOR 55–300 mm f/4.5–5.6 G ED VR (very high-powered telephoto zoom) as evidence, and on-field visual identification were employed.
2.3. Ecological Study Design and In Situ Measurement of Microclimate Data
In this study, intact undisturbed area and adjacent disturbed area were compared to understand the significant impacts of ecological dynamics towards orchid diversity. Four ecological parameters were logged from morning (9 am) to afternoon time (4 pm) under the influence of varying spatial-temporal conditions; temperature (°C), relative air humidity (10%–100%), and light intensity (μmol m−2s−1) (number of photons received per unit time (s) on a unit area (m)) of the study areas, and bark moisture (0%–100%) of the host plants. The temperature and humidity were measured with Extech Hygro-Thermometer (Extech Instruments, Inc., Nashua, NH, USA) at 2 m above the ground during the study period [41]. The light intensity was first measured in Lux unit with Milwaukee MW700 Standard Portable Lux Meter (Milwaukee Instruments, Inc., Rocky Mount, NC, USA) and then value was converted to the Photosynthetic Photon Flux Density (PPFD) unit. Given an illuminance value (Lux), similarly, we calculated the PPFD in micromoles per second per square metre (μmol m−2s−1) for the given light source [42,43]. Unit Lux is simply based on visual sensitivity and does not provide information on the energy or photon content of light, which truly influence the photosynthesis or sugar production in the leaf. Hence, in order to understand better the light intensity for a study relating to plant responses, the suitable unit is the μmol m−2s−1. The bark moisture was measured with HoldPeak HP-2GD Wood Moisture Meter (Zhuhai Jida Huapu Instrument Co., Ltd., Zhuhai, Guangdong Province, China) at 2 m above the basal area.
2.4. Diversity Analyses
Recorded information about the total individual species were analysed statistically and mathematically for clearer identification of diversity in the area. Diversity analyses were estimated using PAleontological STatistics Version 4.04 (PAST4) [44,45]. To determine the species richness and species evenness, Shannon–Wiener Diversity Index (H) and Simpson’s Diversity Index (1-D) were used in this study. Inclusion of both diversity indices improves the output information of the dataset, which is unique for each community or sample analysed. The H and 1-D indices allow data on species richness and relative abundance to be combined [46]. The Simpson Index was explained by presence of the predominant species, and the Shannon Index assumed that individuals were randomly selected, and that all species were represented in the sample [47]. The effect of the sample size is generally insignificant for both indices. However, these methods could not tell which factors contributed more to the value. Hence, Evenness (E) was used to determine how close in numbers each species in each studied area were [48]. Higher values indicate greater community diversity and more even distribution of different individuals within a species. Dominance (D) gives the abundance of the most abundant species and ranges from 0 (all taxa are equally present) to 1 (one taxon dominates the community completely) [44].
2.5. Data Analyses
Abundance was calculated as Relative abundance of orchid (%Ao): [Number of clump or population of a particular orchid species within the transect plots ÷ Total number of all orchid clumps or populations of the transect plots] × 100. When sampling design is transect-based, orchid and host plants are sampled in unequal numbers due to the variation in size of the studied areas or trails. Hence, the following parameters were computed; Density of the orchid plants within a transect plot (plants/km2): Number of orchids plants within the transect plot (plants) ÷ Total area of the transect plots (km2). An ordination method, clustering with UPGMA (Unweighted Pair Group Method with Arithmetic Mean) and Q-mode clustering (grouping variables or associations) by transposing the data matrix [45], was used to look at similarity of host plants (bark textures) amongst the studied vegetation alliances. The similarity was tested using Bray–Curtis Similarity Coefficient [49]. Bray–Curtis is a popular similarity index for abundance data (Hammer et al., 2001). The ordination method was carried out using PAleontological STatistics Version 4.04 (PAST4) [45].
2.6. Statistical Analyses
Ecological data obtained were analysed using software package IBM SPSS version 24 (IBM Corp., Chicago, IL, USA). The measured microclimate parameters were subjected to Shapiro–Wilk for a normality distribution test. As the parametric assumptions could not be obtained for all dependant variables even through transformation (Supplementary Table S1a,c,e,g), non-parametric tests were used for all analyses. The Mann–Whitney test was used to determine the significant relationship of the parameters between the undisturbed and disturbed forests. Statistical significance was determined at p < 0.05 (Supplementary Table S1b,d,f). Following the study [50], the significant difference of mean between groups were determined by comparing directly between the minimum values recorded for undisturbed forests with minimum values recorded for disturbed forests and evaluated accordingly for the maximum values. Kruskal–Wallis one-way analysis of variance (ANOVA) was used to determine the significant difference in daily hours’ ecological parameters between undisturbed, disturbed, and disturbed ‘non-logging sites’ forests with distinctively different disturbance types (Supplementary Tables S1h,j,k). The post hoc for Kruskal–Wallis was conducted using the Mann–Whitney test with multiple pairwise comparisons (Supplementary Table S1i,l). To ensure that Type I error does not t exceed 0.05, Bonferroni correction was used by dividing α of 0.05 by the number of tests conducted. Therefore, the statistical significance was determined at p < 0.017. Comparing species richness among sites or samples is a statistical challenge because the observed number of species is sensitive to the number of individuals counted or the area sampled [51]. Hence, to describe the orchid species richness in the different localities, we used a rarefaction function for each region. Rarefaction is a method for comparing the species richness of samples of different sizes by calculating species richness for a given number of individuals based on rarefaction curves, which are plots of the number of species as a function of the number of samples [52], in our case number of clumps (one clump = one individual = one occurrence). We conducted individual-based rarefaction as implemented in EstimateS 9.1.0 [53].
2.7. Terminology
Following the study [54], the terms ‘host’ and ‘host species’ imply that the focal epiphyte taxon has been observed on the plant individual or species. Since hosts are of several growth forms (trees, shrubs, lianas), the term ‘host plants’ is used rather than ‘host tree’. Note that, in unambiguous contexts, ‘host’ is often used as a short form of ‘host species’ or ‘host plant’. Therefore, the unoccupied (by epiphytic orchids) plant species were excluded from the analysis. In the current study, ‘undisturbed forest’ and ‘disturbed forest’ are the general classification used for the forest types. The terms ‘undisturbed forest’ and ‘disturbed forest’ were adopted from [55]. The synonymous terms such as undisturbed and disturbed areas, or undisturbed and disturbed vegetation have been used in other works of literature on biodiversity in the tropics or Malaysia, such as [18,56,57,58,59,60,61]. ‘Undisturbed forest’ are primary or protected forests that have experienced little to no recent human disturbance, whereas ‘disturbed forest’ are forest that have experienced disastrous and large disturbance such as deforestation, extreme floods, tree falls, and human trampling.
3. Results
3.1. Species Richness and Abundance
In total, 389 orchid specimens confined to 239 species and 65 genera were collected (Supplementary Table S2). A plot of species accumulation curve allows comparison of a number of species at different levels of collecting effort. Using the number of individuals, the disturbed forest was the most species-rich for equal sampling effort if compared to the undisturbed forest (Figure 3). Additionally, the curve for the undisturbed forest stopped at 250 individuals and appeared to approach an asymptote, whereas the curve for the disturbed forest is longer but steeper, indicating that there may be more species found in the disturbed forest if the sampling continues. This may indicate a higher abundance of species which can be found in heterogeneous and larger area sites [62], but nothing of the sort for logging areas as the orchid abundance is influenced by the number of fallen trees and ease of collections for epiphytic orchids on the fallen trees along the skid trails [50].
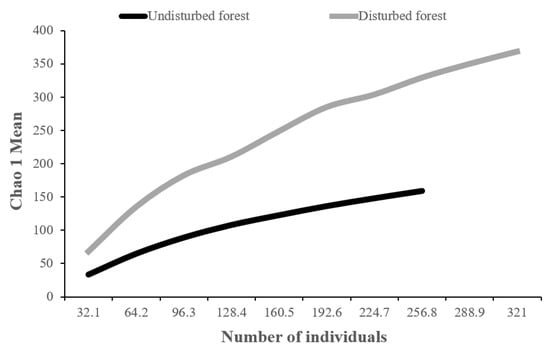
Figure 3.
Orchid species accumulation curves (individual-based rarefaction) within grid points of different land-use intensities.
Species richness in the disturbed forests was 183 species with the logging sites harbouring the most abundant orchids species if compared to the undisturbed forests with 100 species (Figure 4A). Despite that, undisturbed forests had a higher orchid density (2.4 orchids) when compared with the disturbed forests (0.2 orchids). However, disturbed habitats, especially the logging sites, showed greater impoverishment than the undisturbed habitats as very few surviving plants were found or collected. Similarly, Agrostophyllum stipulatum was recorded with highest relative abundance in both undisturbed (3.6%) and disturbed forests (4.6%). The species composition of the orchids differed significantly in between undisturbed and disturbed forests. Disturbed forests comprised mostly epiphytic orchids (Figure 4B), of which the highest collection was from logging sites. Terrestrial species occurred the most in the undisturbed habitats. Only few to none were marginally more frequent in a more open and disturbed habitat. Additionally, species density of orchids based on growth habits was lower in disturbed habitats than in the undisturbed or pristine ones (Figure 4C). The two forest types were Dendrobium and Bulbophyllum as of the most abundant genera which were also mostly epiphytic (Figure 5).
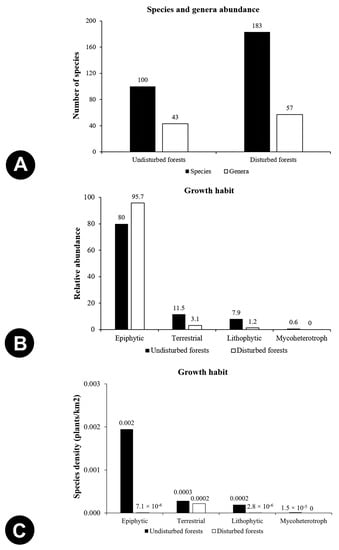
Figure 4.
Comparison of species abundance and density between undisturbed and disturbed forests: (A) Species and genera abundance, (B) Relative abundance based on growth habit, (C) Species density based on growth habit.
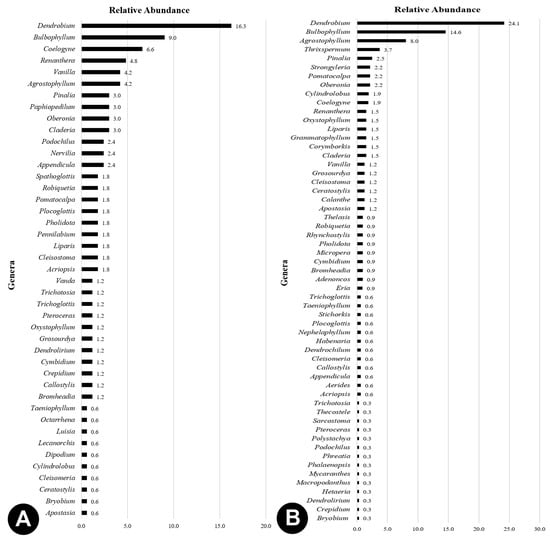
Figure 5.
Comparison of species abundance based on genera for (A) undisturbed and (B) disturbed forests in Peninsular Malaysia.
3.2. Species Diversity
Shannon–Wiener Diversity Index (H) (Figure 6A) and Simpson’s Diversity Index (1-D) (Figure 6B) were showing similar patterns of diversity for the undisturbed and disturbed forests. Both indices showed that disturbed forests harboured higher species diversity than the undisturbed forests. Theoretically, H is strongly influenced by species richness, evenness, and presence of rare species. Meanwhile, 1-D gives more weight to evenness and presence of common species. In this sense, H has an advantage over 1-D because it depends more on species richness and less on abundance, so it is very sensitive to even small diversity changes, and thus is widely used to assess the actual state of environment. Although the finding suggests that disturbed forests could retain a high richness of orchid taxa, there are several detrimental aspects that need consideration—the high abundance of orchids in the disturbed forests might overstate the species richness. The low orchid diversity in the undisturbed forest is supported by the high dominance of the most dominant species (D = 0.015) (Figure 6C), and the slightly higher evenness in the undisturbed forest (E = 0.844) does not seem to affect the H and 1-D values much (Figure 6D). Additionally, the higher diversity of orchids in the disturbed forests might be influenced by the higher number of rare species found on the fallen trees in the logged areas (Figure S1). Moreover, forest margin had lower species diversity (Figure 7A) and density (Figure 7C) than the adjacent undisturbed forest with no terrestrial orchid recorded in the area. It suggests the altered environment caused by the adjacent logging activity reduced the orchid species richness. In logging sites, orchid species were greatest in diversity if compared to the undisturbed forests (Figure 7E), but lower in species density (Figure 7G). It only comprised of epiphytic orchids (Figure 7H). Further details on the occurrence, diversity, and density of orchids in each study site are presented in Table 4.
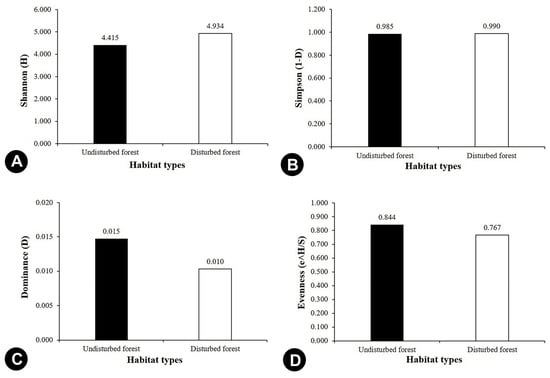
Figure 6.
Overall comparison of species diversity between the undisturbed and disturbed forests: (A) Shannon (H), (B) Simpson (1-D), (C) Dominance, (D) Evenness (e^H/S).
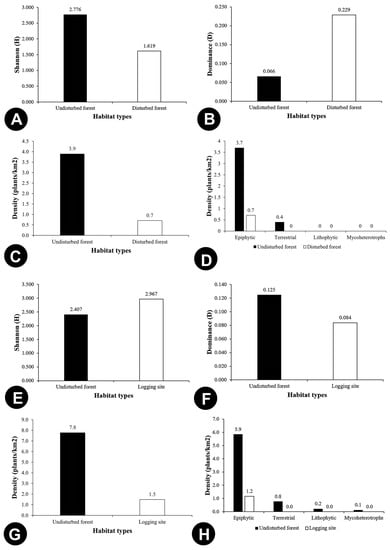
Figure 7.
Comparison of species diversity and density for Adjacent Undisturbed vs. Disturbed Forests and Undisturbed Forests vs. Logging Sites: (A,E) Shannon (H), (B,E) Dominance (D), (C,G) Density (plants/km2) based on forest types, (D,H) Density (plants/km2) based on growth habit.
Table 4.
Comparison based on diversity, density, and distribution of life forms of orchid species in undisturbed and disturbed forests in Peninsular Malaysia.
3.3. Microclimate Conditions
Overall data of the microclimate conditions (Supplementary Table S3) show that maximum temperature for the disturbed forests were significantly higher (p < 0.001, df = 58) than the undisturbed ones. Simultaneously, for the relative air humidity, the disturbed forests had significantly lower values (p < 0.001, df = 58) if compared to the undisturbed forests (Table 5, Figure S2). Additionally, a comparison on microclimate conditions in between two adjacent forests, an undisturbed forest (BK) and a forest margin (BK(M)) located next to a logging site (LBK), was conducted to see whether there were any significant differences in microclimate between the two forests contrasting in level of disturbance. The result shows there were significant differences in maximum temperature (p < 0.001, df = 38), minimum and maximum relative air humidity (p < 0.001, df = 38), and minimum (p = 0.010, df = 38) and maximum (p < 0.001, df = 38) light intensity in between the two adjacent forests (Table 5, Figure S3). It indicates that the disturbance caused by adjacent logging projected extreme microclimate conditions on the environment. Also, the result shows significant differences in temperature (p < 0.001, df = 38), relative air humidity (p < 0.001, df = 38), and light intensity (p < 0.001, df = 38) in between the undisturbed forests and extremely disturbed forests (logging areas) (Table 5, Figure S4). The canopy disruptions caused by the selective-cut and clear-cut logging produced openings in the canopy, which had significantly affected the temperature and the light intensity. The temperature could reach up to 40 °C in the afternoon and consequently reduced the humidity to 51%. It indicates extreme dryness in the logging areas, limiting growth and distribution of orchids.
Table 5.
Summary of topographic and ecological predictors (mean ± SE) of undisturbed and disturbed forests in Peninsular Malaysia.
Figure 8 shows the hourly means for temperature and relative air humidity differed between habitats of different disturbance regimes. Hourly means differed between disturbance regimes during daytime from 10 am to 4 pm. The corresponding hourly temperature for the logging sites were significantly higher at the consecutive period of day (morning, H(1) = 26.38, p = 0.002, df = 2; afternoon, H(1) = 26.75, p < 0.001, df = 2; late afternoon, H(1) = 20.33, p < 0.001, df = 2) if compared to the undisturbed forests (morning, H(1) = 14.00, p = 0.002, df = 2; afternoon, H(1) = 13.10, p < 0.001, df = 2; late afternoon, H(1) = 5.85, p < 0.001, df = 2). Similarly, the result shows significantly higher temperature for the logging site (morning, H(1) = 12.63, p = 0.005, df = 2; afternoon, H(1) = 22.72, p < 0.001, df = 2; late afternoon, H(1) = 19.35, p < 0.001, df = 2) than the non-logging site disturbed forests (morning, H(1) = 5.78, p = 0.005, df = 2; afternoon, H(1) = 11.62, p < 0.001, df = 2; late afternoon, H(1) = 5.33, p < 0.001, df = 2). The result also showed significant difference for relative air humidity in between the logging sites (morning, H(1) = 6.56, p = 0.011, df = 2; afternoon, H(1) = 5.00, p < 0.001, df = 2; late afternoon, H(1) = 5.44, p < 0.001, df = 2) and undisturbed forests (morning, H(1) = 13.10, p = 0.011, df = 2; afternoon, H(1) = 14.50, p < 0.001, df = 2; late afternoon, H(1) = 14.10, p < 0.001, df = 2). The result was also with significant difference for relative air humidity in between the logging sites (morning, H(1) = 7.06, p = 0.050, df = 2; afternoon, H(1) = 5.44, p < 0.001, df = 2; late afternoon, H(1) = 5.33, p < 0.001, df = 2) and non-logging disturbed forests (morning, H(1) = 11.94, p = 0.050, df = 2; afternoon, H(1) = 13.56, p < 0.001, df = 2; late afternoon, H(1) = 13.67, p < 0.001, df = 2), except for morning time.
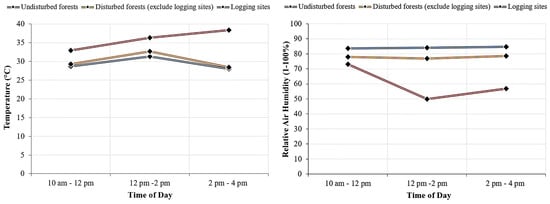
Figure 8.
Daily course of temperature and relative air humidity in undisturbed and disturbed forests for different disturbance regimes in Peninsular Malaysia (2016–2020). Note: Each point represents means of period of day, morning (10 am–12 pm), afternoon (12 pm–2 pm), and late afternoon (2 pm–4 pm).
3.4. Factors Influencing Abundance of Epiphytic Orchids: Host Plants and Bark Moisture
Effects of microclimate change due to the anthropogenic and naturogenic activities are now often examined based on the forest canopy species, especially epiphytic orchid species that rely on the host plants. In this paper, the general host characteristics and bark moisture that may influence the distribution and abundance of epiphytic orchids are mainly explored. In total, 102 host individuals harbouring wild orchids were inspected in all the studied line-transect plots. There are five most significant types that accumulate abundant epiphytic orchids, Type 4, 6, 13, 18, and 19 (Table 6), with wide and interlocking crown roofing the understorey part from sun exposure and dryness. Additionally, the study highlights the importance of the host plants that possess bark texture Type 9 and 12 (Table 6), smooth to fissured textures. The result demonstrated that epiphytic orchid diversity and abundance is favoured by rough-barked host. Only 15 families of host species were present in the study localities. Total host species was undetermined because a number of host individual or the fallen trees recorded in the logging sites were only identified to family level due to the missing of important morphological characters, such as leaves and flowers. In the undisturbed and non-logging disturbed forests, epiphytic orchids were prominently abundant growing on Saraca, Sygyzium, and Neonauclea tree species that were growing along the riparian areas. Many clumps of epiphytic orchids were also recorded growing on large snags (dead standing trees) that were covered by moist substrate (mosses and lichens) in riparian forests. On average, an individual host tree could have harboured more than ten epiphytic orchid species. Species abundance of epiphytic orchids correlated with the bark moisture and texture. A box plot analysis shows that host plants having bark moisture 60%–80% generally harboured more epiphytic orchids than hosts with 0%–20%, 20%–40%, 40%–60%, and 80%–100% bark moisture (Figure 9). Fifteen distinct bark textures were recorded and described based on the surface morphology (Table 7 and Figure 10). Using cluster analysis (Figure 11), the difference of bark textures of host plants, those of undisturbed forest or non-logging disturbed forest or logging site were distinguished. Bark textures are generally grouped well based on the vegetation type and disturbance level. This suggests bark textures could be associated with the microclimate conditions of the forest. Furthermore, this explains that the textures that potentially influenced the abundance of epiphytic orchids were likely to be modulated by climate.
Table 6.
Types of host plants harbouring orchids in the undisturbed and disturbed forests in Peninsular Malaysia.
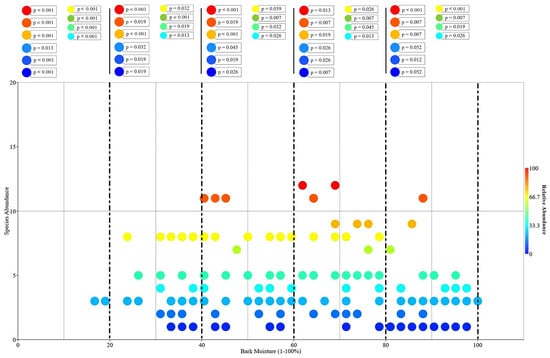
Figure 9.
Relation between total epiphyte species density (species per tree) and host plants’ bark moisture (bubble plot; N = 150). According to their colour, cold to hot colour, bubbles reflect multiple values.
Table 7.
Types of bark texture of the host plants harbouring epiphytic orchids in the undisturbed and disturbed forests in Peninsular Malaysia.

Figure 10.
Types of bark texture of host plants in undisturbed and disturbed forests in Peninsular Malaysia: (A) Type 1, (B) Type 2, (C) Type 3, (D) Type 4, (E) Type 5, (F) Type 6, (G) Type 7, (H) Type 8, (I) Type 9, (J) Type 10, (K) Type 10, (L) Type 11, (M) Type 12, (N) Type 13, (O) Type 14, (P) Type 15.
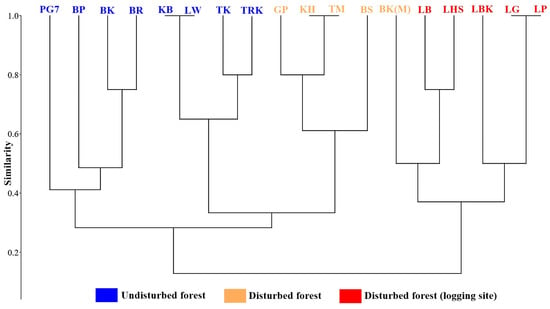
Figure 11.
Similarity of host plants (bark textures) amongst the studied vegetation alliances using Bray–Curtis Similarity Coefficient.
4. Discussion
4.1. Ecological Implications of Current Findings on Orchid Diversity
Species richness, diversity, and abundance of the wild orchids differed in the different localities. It is known that orchid species richness differs across a land-use intensity gradient [3,63]. Generally, increment in the number of orchid individuals at each locality is corresponding to the classical species-area relationship where larger areas tend to contain larger numbers of species [64,65]. Moreover, in certain occasions, in extremely disturbed and dry forests, orchids do not decrease in species diversity and density with increasing disturbance, suggesting that orchids may indeed be comparatively disturbance-resilient [41,50]. In the current study, high abundance of orchids in disturbed forests was majorly comprised of epiphytic orchids whose occurrence was highly influenced by number of fallen trees, accessibility, and ease of human access for collections in the logged forest areas [50,66,67]. The problem of greater access to fallen trees in logged areas seems to be a methodological problem that appears to invalidate the ‘higher diversity’ found in the disturbed forests. Despite the vast area, the number and distribution of wild orchids in the deforested area were influenced by the number of host plants for epiphytic orchids and the microclimate conditions. Additionally, in some logging sites, especially the ones that have been neglected for many years, only one or two fallen trees are found over such a large area. Moreover, since most of the ground vegetation has been cleared for skid trails, ground orchids are rarely seen in the clearing zone and therefore have a low density. As in this study, species density decreased with increased disturbance level. The microclimate of disturbed forests was markedly altered from that of undisturbed forests, similarly, reported in [68] (also cited in [41]). Temperature, air humidity, and light intensity in the undisturbed forests and the extremely disturbed forests (logging sites) were significantly different proving that opening up of the forest canopy resulted in significant microclimate changes, which in turn affected the wild orchids [50,69].
Impoverishment of the logging sites was eminent, greatly contrasting with the non-logging disturbed forests and the undisturbed forests. Logging sites were reported with extremely high temperature and low relative air humidity and host plants’ bark moisture. The abundance of epiphytes usually decreases in sites with lower precipitation [14,41,70]. The smaller number of orchids’ taxa and terrestrial orchids in the non-logging disturbed forests (excluding logging sites) than the undisturbed forests indicates that the study area with extreme microclimate condition is a marginal habitat for wild orchids and seems to confirm the importance of temperature and humidity [41,70,71,72], light intensity, and the ground conditions [50] as a driver of wild orchids’ species richness. Dry microclimate in the secondary forest, caused by the more open canopy of the secondary forest and the stronger radiation drove impoverishment [69]. The authors of [69] report that high temperature, low humidity, and lack of a dense bryophyte cover affected the bark moisture, and apparently had cascading effects on the diversity of epiphytic orchids in these forests. Epiphyte species should show stronger host specificity in habitats where climatic conditions are suboptimal for their performance because the modulating effect of tree traits is stronger under such conditions [73].
Presumably, drier sites featured only the dominant, robust, and drought-tolerant species [41,50,74] such as of Bulbophyllum and Dendrobium species. This explains the dominance of these genera in both undisturbed and disturbed forests. The drought-tolerant orchids tolerate drought instead of avoiding it [75]. They may be limited in their ecological distribution by their inability to maintain a positive carbon balance during repeated cycles of wetting and drying [76]. The lack of broad, thick leaves (thick cuticles) and trichomes (which protect against radiation and reduce water loss by protecting against wind and heat) are associated with their drought avoidance phenology. The plants produce roots at the beginning of the wet season and shed them as the dry season approaches [77,78]. Some plants possess storage organs such as pseudobulbs or pseudobulbous stem and rhizome to store water [79].
The results also show that sites with little anthropogenic influence have higher species densities than those with high anthropogenic influence. Primary or undisturbed forest areas have more natural vegetation and a protected microclimate, are less polluted, less disturbed by humans, including collecting pressures, and have more abundant host tree species with suitable characteristics [80,81]. The contradicting results of orchid diversity between undisturbed and disturbed forests from this study may also have been influenced by the unseen (unmeasured) drivers such as microbiomes [82,83] and soil properties [18,84].
Based on the genera abundance, the orchid flora of disturbed forests was not distinct from the canopy-protected and undisturbed forests, but still, the species richness and density were reduced. Forests that were either regenerating after clear-cutting or surrounded by plantations generally show considerably reduced species number compared to the matured and primary ones. Similar observations have been made in several studies, in which secondary moist forests regenerating after clear-cutting generally have significantly reduced epiphyte species abundance compared to mature forests (vascular plants [14,85,86], bryophytes [2,87,88,89], epiphyte [41]). Three sites we studied, BS, GP, KH, which were regenerated and secondary forests, showed a similar pattern. The authors of [63,81] reported that species richness and abundance of epiphytic orchids gradually declined significantly with increasing habitat modification from unmanaged via remnant forest patches, agricultural land to very strong land-use intensity. This could indicate the regeneration of wild orchid communities in the disturbed forests following disturbance is very slow. However, this depends on how the forests are managed after cutting. For instance, one of our studied sites, PG7, is a regenerated forest that has now gazetted as a High Conservation Value Forest (HCVF) area, showing a considerably high diversity and density of orchids compared to the other studied undisturbed and disturbed sites. Different management practices influenced the orchid species and their composition and density.
4.2. Applications of the Current Findings for Informing Conservation and Restoration of Orchids
This investigation also gives prominence to the importance of the adjacent forests or the remaining natural forests for restoration as the forests constitute biological sources that enable succession from species-poor systems to highly diverse ones [90,91]. However, the disturbance projected by the forest clearing affected the ecological conditions and environment of the canopy-covered adjacent area. Forest clearing created forest margin or forest edge, where a large area of the forests was opened and disturbed, and a number of fallen trees cut during logging fell into the adjacent area. The adjacent area experienced increased temperature and light intensity. Consequently, it had lower diversity and density, too, if compared to the adjacent undisturbed forest. Similarly reported in studies [41,92,93], edge habitats experienced increased solar radiation, air temperature, vapour pressure deficit, and wind speed compared to the forest interior. Edge habitat fostered lower total species richness than any other habitat, but was similar in composition to closed forest [41]. Projected warming threatens the persistence of a population unless these populations possess a high micro-evolutionary potential that aid them in tolerating the environmental changes [94]. This includes species or certain species from genera Bulbophyllum, Coelogyne, Cymbidium, Eria, Liparis, Oberonia, Paphiopedilum, Panisea, Pholidota, and Vanilla [95]. Based on the comparative study on ecological and forest structure in Bukit Batu Kapal, Terengganu, a lowland to hill forest area, the undisturbed patches were home to abundant orchids and mature tree species, whereas the adjacent logged and secondary forest was composed of Bulbophyllum and Coelogyne species, and invasive and pioneer ‘non-orchid’ plant species. Bulbophyllum and Coelogyne species employed ‘drought escape’ adaptive strategy by shedding leaves and roots in dry conditions and leaving dormant pseudobulbs or stems to minimise transpiration.
The species composition of epiphytic orchids depends on several factors other than temperature, humidity, and light intensity, such as host plant species and bark characteristics as shown in [15,63]. Associations between epiphytic orchids and host characteristics are among the main factors affecting the diversity and distribution of epiphytes [3]. Along with altered microclimate, this has been attributed to reduced surface area for colonisation, and the lack of late successional substrates on young, small, and fast-growing host trees. In comparison with study [87], an analysis of tree-wide cortical moss diversity in primary rainforests, specialists recovered more slowly than generalists. In addition to this, the dominance of invasive tree and shrub species may restrict colonisation by epiphytic species. Exotic species such Acacia, invasive species such as Bambusa, and native pioneer species such as Macaranga have smooth bark texture, less favourable for the epiphyte colonisation, including the mosses and lichens. In forest areas that have been subjected to disturbance, Acacia, Macaranga, bamboos, and several species of early successional woody shrubs, such as Melastomataceae are common. Other than that, Macaranga, Arenga, and Acacia have a dominant presence in the secondary or converted forests. Epiphytic orchid specificity towards certain or single species of host plants was rather weak, in fact, their preference was more on the host characteristics regardless of species. According to studies [54,96], host specificity exists when mutual trade-offs prevent a host-dependent species from adapting equally well to the full range of conditions found in different host species. The same species of orchids found on a variety of host plant species have similar characteristics in undisturbed lowland forests. This is a ‘parallel host-specific scenario’, a term coined in [54], in which all epiphyte species exhibit the same low or high performance on a particular tree species. Moreover, previous studies of few and small populations of an epiphytic orchid species increases the likelihood that limitation to a single tree species was simply due to randomness in combination with host biases [8,97]. For instance, the epiphytic orchids preferred host plant species, such as Tristaniopsis, Syzygium, and Eurya species growing along the riparian area, but not in the dry habitat as in the heath vegetation and belukar (bushes). In the riparian area, overtopped and intermediate trees were canopy-protected and constantly moist with thick substrate covering the trunk and branches. Host plants with smooth and dry bark texture, such as Leptospermum javanica, Calophyllum species, and Tristaniopsis merguensis, were abundant in Padang 7 which is peaked with coastal heath vegetation. Fewer epiphytes growing on these trees was not just because of the unsuitable bark texture, but was also highly affected by the extreme hot and dry environment of the heath vegetation. In fact, epiphytic orchids’ associations with many host bark textures varied between localities indicating that some of the bark textures are locality-specific. Moreover, certain host traits may change during ontogeny which, in turn, may lead to differential ‘life stage biases’ [54]. These traits include leaf size and deciduousness [98] and bark properties [99].
Although species specificity (host-orchid species specificity) was not proved here in the current study, previous works have shown that epiphytes depend to some degree on potential host species [40] and some orchid species are restricted to certain host tree species [63,100,101]. In the protected forests, tall and large trees with Type 6 bark texture (soft, small fissured, peeled off, warty) were common, but rare in the secondary and intensively used forests. Tall and large trees in disturbed forests are often isolated. The crowns of such isolated large trees are also isolated from each other, meaning that large parts of the crowns are exposed to sun, favouring the orchids which use pseudobulbs as a water reservoir [4]. This was also observed in the current study where only the Bulbophyllum and Coelogyne were commonly dominating the isolated trees. Thus, large and old host tree individuals and host plants with rough or fissured bark are essential for the protection of epiphytic biodiversity.
However, not only the tree size but also the suitability of the stand to provide a microhabitat for the epiphytes is important. Some epiphytic orchids are more common in smaller trees and shrubs, similar to what was observed in [81,102]. Small trees with spreading crown along the riparian areas would accumulate as much as the tall trees. High abundance of species was not only observed on tall trees but also on smaller trees with spreading crown along the riparian area because, similarly, they are exposed to light and moisture. Host plants growing in the riparian area offer epiphytes specific microclimatic conditions. Similarly, the authors of [103,104,105,106] observed that the most preferred tree species occupied riparian areas, lower slopes, and valleys near swamps. These type of host plants were observed with very low abundance in disturbed forests. As reported in study [54], there are numerous instances of extreme basic host specificity that are not the typical ‘tree’ growth morphology. A notable example here is liana. Lianas such as Spatholobus species also accumulate epiphytic orchids, but the orchids were not recorded as being abundant as on trees.
Previous studies indicate that host species with rough bark are more frequently colonised by epiphytes [15,63,107,108]. The reasons for more epiphytes occurring on rough and fissured bark are that they retain moisture for longer and seedling recruitment is better as seed lodges more easily in the crevices in bark than on smooth bark [109,110]. Host plants with either flaking, large peeling or smooth bark, and low substrate cover are observed as poor hosts. The colonisation of bryophytes and lichens may influence substrate suitability for vascular epiphytes [8,71,111,112]. However, quantitative evidence is lacking to support this claim. Rarity of the epiphytic orchids in young secondary forests was influenced by bark characteristics and tree architecture such as having little-branched crowns with oblique instead of horizontal branches [69]. This type of crown architecture is unfavourable for the establishment of epiphytes ([113]; cited in [69]). In secondary forests in Peninsular Malaysia, trees with round and overtopped crowns with a height of 6 m to 10 m are common. The hosts’ bark texture significantly increased towards higher human impact, showing significant differences between all intensities [63]. Previous studies indicate that the diversity of epiphytes is positively correlated with host characteristics of bark texture [15,63], type of host (deciduous or evergreen) [114], pH of bark, light, wind, and tree diameter [115]. Unfortunately, in this study, the hosts’ ages and sizes (diameter at breast height, tree height, and canopy volume) were not fully accounted for, as the age and size of some individuals of the fallen trees and lianas are difficult to verify. Additionally, the substrate cover was recorded only based on their general distribution on the hosts.
Terrestrial and mycoheterotroph orchids were found to be sensitive indicators of microclimate and human disturbance in the disturbed forests. Since most of the terrestrial and mycoheterotroph orchid species lack water storage organs and leaf cuticles, they are highly dependent on the soil conditions, more sensitive to forest disturbance, and favoured in forests with a dense tree canopy. Based on the findings, the terrestrial orchid species were underrepresented in disturbed forests. Additionally, the predominance of lithophytic orchids growing in rocky outcrop habitats, such as conglomerate hill and limestone hill forests, is significant. Some orchid species in the rocky outcrops are capable of living in two growth forms, growing as epiphytic and lithophytic. Many orchids, such as Bulbophyllum, Coelogyne, and Dendrobium species, which are normally epiphytic, but also grow on the ground or on limestone cliffs, similar to those reported in [116], are on the summit areas with high light levels. Their adaptation increases the likelihood that the rocky outcrop is lacking alternative hosts [54]. As reported in study [117], some shrubby or arborescent, mat-forming monocots have been found as alternative hosts on rocky outcrops in Africa and South America, including several Velloziaceae [118,119,120,121] and one Cyperaceae [117]. In this study, the alternative hosts could be bamboos (Poaceae) and Eugeissona or bertam (Arecaceae) growing at the bottom of rocky outcrops, but wild orchids were not seen growing there. Bases of limestone hills and ravines are usually shaded and have high humidity, which often creates a microhabitat that is suitable for terrestrial orchids [18,116], including Paphiopedilum niveum (Rchb.f.) Stein, Nervilia concolor (Blume) Schltr., and a Crepidium species.
5. Conclusions
Conclusively, microclimatic changes, anthropogenic disturbance, and low abundance of suitable host plants were key determinants of wild orchid composition and colonisation. We suggest these criteria should be considered in forest restoration to reduce disturbance impact on the associated orchid community. The study reported high diversity of epiphytic orchids and big differences in species richness, abundance, and species composition of orchid communities in different localities across disturbance gradients in Peninsular Malaysia. Wild orchids showed sensitivity to habitat quality and disturbance. Generally, wild orchid species density was decreasing with increasing human impact, especially with removal of host plants and extreme high temperature and light, and low humidity and bark moisture. Occurrence of terrestrial and mycoheteretroph orchids are indicators of microclimatic conditions and human disturbance. In non-logging forests, orchid abundance changes along the disturbance gradient. Epiphytic assemblages exhibit correlation to the type, bark texture, and bark moisture of the host plants. Conservation values of the suitable host plants for epiphytic orchids’ diversification is worth further study. The significant difference in microclimate of the adjacent undisturbed and disturbed forests appears to underline the extreme effect of the disturbance which happened in the disturbed area towards the intact adjacent area. Forest and ecological alterations caused a shift in patterns of orchid species composition, where the robust and sun-loving species are dominating the forests. However, the considerable effect of the disturbance towards the community in the specific vegetation types requires further research because our current study was limited to the pre-created trails only.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14030544/s1, Table S1: Test of normality and test statistics for ecological parameters recorded in undisturbed and disturbed forests in Peninsular Malaysia; Table S2: List of orchid species collected from the undisturbed and disturbed forests in Peninsular Malaysia; Table S3: In situ ecology parameters recorded in undisturbed and disturbed forests in Peninsular Malaysia; Figure S1: Comparison of species abundance based on distribution: (A) Undisturbed forests and (B) Disturbed forests; Figure S2: In situ ecology parameters recorded for the Undisturbed Forests vs. Disturbed Forests. Different superscripts indicate significant differences (p < 0.05) in Mann–Whitney test with multiple pairwise comparisons; Figure S3: In situ ecology parameters recorded for the Adjacent Undisturbed Forests vs. Disturbed Forests. Different superscripts indicate significant differences (p < 0.05) in Mann–Whitney test with multiple pairwise comparisons; Figure S4: In situ ecology parameters recorded for the Undisturbed Forests vs. Extremely Disturbed Forests (Logging Sites). Different superscripts indicate significant differences (p < 0.05) in Mann–Whitney test with multiple pairwise comparisons.
Author Contributions
Conceptualization, E.E.B. and R.G.; methodology, E.E.B. and R.G.; validation, E.E.B. and R.G.; formal analysis, E.E.B.; investigation, E.E.B.; resources, E.E.B. and R.G., data curation, E.E.B. and R.G.; writing—original draft preparation, E.E.B. and R.G.; writing—review and editing, E.E.B., R.G., M.M. and C.S.Y.Y.; visualization, E.E.B.; supervision, R.G., M.M. and C.S.Y.Y.; project administration, R.G.; funding acquisition, E.E.B. and R.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the UPM-KRIBB (Korea Research Institute of Bioscience and Biotechnology) grant (Vot. 6384300), FRGS-MOHE (Vot. 5524110), UPM-Putra IPS Grant (Vot. 9695200), Tenaga Akademik Muda (TAM) issued by Universiti Putra Malaysia (UPM), and Skim Latihan Akademik IPTA (SLAI) issued by the Ministry of Higher Education Malaysia (MOHE).
Data Availability Statement
Data are contained within the article or Supplementary Materials.
Acknowledgments
We sincerely thank the Forest Department Peninsular Malaysia who granted us the access permit to the studied area. We also gratefully acknowledge the help provided by our staff and friends in the department. We thank our fellow experts for their help with the plant identification and consultation, Mohd Norfaizal Ghazalli (Plant Taxonomist, Malaysian Agricultural Research and Development Institute (MARDI), Malaysia) and Ahmad Fitri Zohari (Plant Taxonomist, Universiti Kebangsaan Malaysia (UKM), Malaysia). Our gratitude also goes to the reviewers and to Jean W.H. Yong (Plant Biologist, Swedish University of Agricultural Sciences (SLU), Sweden), Mohd Nazre Saleh@Japri (Plant Biologist, Universiti Putra Malaysia (UPM)), and Faridah Qamaruzzaman (Plant Conservation Biologist, Universiti Putra Malaysia (UPM)) for their valuable comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Cribb, P.J.; Kell, S.P.; Dixon, K.W.; Barrett, R.L. Orchid Conservation: A Global Perspective. In Orchid Conservation; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2003; pp. 1–24. [Google Scholar]
- Nöske, N.; Hilt, N.; Werner, F.; Brehm, G.; Fiedler, K.; Sipman, H.; Gradstein, S. Disturbance effects on diversity in montane forest of Ecuador: Sessile epiphytes versus mobile moths. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Timsina, B.; Rokaya, M.B.; Münzbergová, Z.; Kindlmann, P.; Shrestha, B.; Bhattarai, B.; Raskoti, B.B. Diversity, distribution and host-species associations of epiphytic orchids in Nepal. Biodivers. Conserv. 2016, 25, 2803–2819. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S.; Rokaya, M.B.; Bhattarai, P.; Gruppe, A. Diversity, composition and host-species relationships of epiphytic orchids and ferns in two forests in Nepal. J. Mt. Sci. 2017, 14, 1065–1075. [Google Scholar] [CrossRef]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of tree age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Flores-Palacios, A.; García-Franco, J.G. The relationship between tree size and epiphyte species richness: Testing four different hypotheses. J. Biogeogr. 2006, 33, 323–330. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S. Epiphytic orchids and their ecological niche under anthropogenic influence in central Himalayas, Nepal. J. Mt. Sci. 2016, 13, 774–784. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Zimmerman, J.K.; Lebrón, L.; Bayman, P.; Sastre, I.; Axelrod, F.; Alers-García, J. Host specificity and low reproductive success in the rare endemic Puerto Rican orchid Lepanthes caritensis. Biol. Conserv. 1998, 85, 297–304. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef]
- Colwell, R.K.; Dunn, R.R.; Harris, N.C. Coextinction and persistence of dependent species in a changing world. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 183–203. [Google Scholar] [CrossRef]
- Steege, H.T.; Cornelissen, J.H.C. Distribution and ecology of vascular epiphytes in lowland rain forest of Guyana. Biotropica 1989, 21, 331–339. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, J.; He, J.S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological diversity of orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef]
- Hietz, P. Diversity and Conservation of Epiphytes in a Changing Environment. In Proceedings of the Invited Lecture at the International Conference on Biodiversity and Bioresources: Conservation and Utilization, Phuket, Thailand, 23–27 November 1997; (Published in 1999). Available online: https://media.iupac.org/symposia/proceedings/phuket97/hietz.pdf (accessed on 10 July 2022).
- Callaway, R.M.; Reinhart, K.O.; Moore, G.W.; Moore, D.J.; Pennings, S.C. Epiphyte host preferences and host traits: Mechanisms for species-specific interactions. Oecologia 2002, 132, 221–230. [Google Scholar] [CrossRef]
- Partomihardjo, T.; Eizi, S.; Junichi, Y. Development and distribution of vascular epiphytes communities on the Krakatau Islands, Indonesia. South Pac. Stud. 2004, 25, 7–26. [Google Scholar]
- Hsu, R.C.C.; Wolf, J.H.; Tamis, W.L. Regional and elevational patterns in vascular epiphyte richness on an East Asian Island. Biotropica 2014, 46, 549–555. [Google Scholar] [CrossRef]
- Saw, L.G. Vegetation of Peninsular Malaysia. Seed Plants 2010, 1, 21–41. [Google Scholar]
- Whitmore, T.C. Tropical Rain Forests of the Far East, 2nd ed.; ELBS/Oxford University Press: Oxford, UK, 1984; p. 295. [Google Scholar]
- Kiew, R. A Flora Survey of Limestone Hills in Perlis; Project No. 272/93; World Wildlife Fund: Selangor, Malaysia, 1993; p. 58. [Google Scholar]
- Calderon-Aguilera, L.E.; Rivera-Monroy, V.H.; Porter-Bolland, L.; Martínez-Yrízar, A.; Ladah, L.B.; Martínez-Ramos, M.; Alcocer, J.; Santiago-Pérez, A.L.; Hernandez-Arana, H.A.; Reyes-Gómez, V.M.; et al. An assessment of natural and human disturbance effects on Mexican ecosystems: Current trends and research gaps. Biodivers. Conserv. 2012, 21, 589–617. [Google Scholar] [CrossRef]
- Bridson, D.; Forman, L. The Herbarium Handbook, 3rd ed.; Royal Botanic Gardens: Kew, UK, 2000; p. 348. [Google Scholar]
- Seidenfaden, G.; Wood, J.J. The Orchids of Peninsular Malaysia and Singapore; Royal Botanical Garden: Kew, UK, 1992; p. 779. [Google Scholar]
- Turner, I.M. A catalogue of the vascular plants of Malaya: Orchidaceae. Gard. Bull. Singap. 1995, 47, 559–620. [Google Scholar]
- Comber, J.B. Orchids of Sumatra; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2001; p. 1026. [Google Scholar]
- Ong, P.T.; O’Bryne, P.; Saw, L.G.; Chung, R.C.K. Checklist of Orchids of Peninsular Malaysia; Research Pamphlet (No. 136); Perpustakaan Negara Malaysia: Kuala Lumpur, Malaysia, 2017; p. 169. [Google Scholar]
- Govaerts, R.; Bernet, P.; Kratochvil, K.; Gerlach, G.; Carr, G.; Alrich, P.; Pridgeon, A.M.; Pfahl, J.; Campacci, M.A.; Baptista, D.H.; et al. World Checklist of Orchidaceae; The Board of Trustees of the Royal Botanic Gardens: Kew, UK; Available online: http://apps.kew.org/wcsp/ (accessed on 22 November 2021).
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Cooperation: Kuala Lumpur, Malaysia, 1966; Volume 1 (A–H), p. 1240.
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Cooperation: Kuala Lumpur, Malaysia, 1966; Volume 2 (I–Z), p. 2444.
- Ng, F.S.P. Tree Flora of Malaya; Volume 3; Forest Research Institute of Malaysia: Kepong, Malaysia, 1978; p. 339.
- Ng, F.S.P. Tree Flora of Malaya; Volume 4; Forest Research Institute of Malaysia: Kepong, Malaysia, 1989; p. 549.
- Ridley, H.N. Introduction. In Flora of Malay Peninsula; Ridley, H.N., Ed.; Volume, 1; Reeve & Co.: London, UK, 1922; pp. ix–xx. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 1, Polypetalae; L. Reeve & Company, Limited: London, UK, 1922; p. 470. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 2, Gamopetalae; L. Reeve & Company, Limited: London, UK, 1923; p. 672. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 3, Apetalae; L. Reeve & Company, Limited: London, UK, 1924; p. 406. [Google Scholar]
- Ridley, H.N. The Flora of the Malay Peninsula; Volume 4, Monocotyledones; L. Reeve & Company, Limited: London, UK, 1924; p. 383. [Google Scholar]
- Whitmore, T.C. Tree Flora of Malaya; Volume 1; Forest Research Institute of Malaysia: Kepong, Malaysia, 1972; p. 473.
- Whitmore, T.C. Tree Flora of Malaya; Volume 2; Forest Research Institute of Malaysia: Kepong, Malaysia, 1973; p. 444.
- Ingram, S.W.; Lowman, M.D. The collection and preservation of plant material from the tropical forest canopy. For. Canopies 1995, 22, 587–603. Available online: https://canopymeg.com/PDFs/papers/0047.pdf (accessed on 12 June 2021).
- Migenis, L.E.; Ackerman, J.D. Orchid—Phorophyte relationships in a forest watershed in Puerto Rico. J. Trop. Ecol. 1993, 9, 231–240. [Google Scholar] [CrossRef]
- Werner, F.A.; Gradstein, S.R. Diversity of dry forest epiphytes along a gradient of human disturbance in the tropical Andes. J. Veg. Sci. 2009, 20, 59–68. [Google Scholar] [CrossRef]
- McCree, K.J. Tests of Current Definitions of Photosynthetically Active Radiation Against Leaf Photosynthesis Data. Agric. For. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- Thimijan, R.W.; Heins, R.D. Photometric, Radiometric, and Quantum Light Units of Measure: A Review of Procedures for Interconversion. Hortic. Sci. 1983, 18, 818–822. [Google Scholar] [CrossRef]
- Harper, D.A.T. Numerical Palaeobiology–Computer-Based Modelling and Analysis of Fossils and Their Distributions; John Wiley & Sons: Oxford, UK, 1999; p. 468. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 12 June 2021).
- Gorelick, R. Combining richness and abundance into a single diversity index using matrix analogues of Shannon’s and Simpson’s indices. Ecography 2006, 29, 525–530. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Biodiversity; Blackwell Publishing: Malden, MA, USA, 2004; p. 256. [Google Scholar]
- Help, C. A new index measuring evenness. J. Mar. Biol. Assoc. United Kingd. 1974, 54, 555–557. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Besi, E.E.; Nikong, D.; Mustafa, M.; Go, R. Orchid diversity in anthropogenic-induced degraded tropical rainforest, an extrapolation towards conservation. Lankesteriana 2019, 19, 107–124. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9; User’s Guide and application; 2013. Available online: http://purl.oclc.org/estimates (accessed on 13 December 2021).
- Wagner, K.; Mendieta-Leiva, G.; Zotz, G. Host specificity in vascular epiphytes: A review of methodology, empirical evidence and potential mechanisms. AoB PLANTS 2015, 7, plu092. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: New York, NY, USA, 1985; p. 472. [Google Scholar] [CrossRef]
- Turner, M.G.; Dale, V.H.; Everham, E.H. Fires, hurricanes, and volcanoes: Comparing large disturbances. BioScience 1997, 47, 758–768. [Google Scholar] [CrossRef]
- Turner, M.G.; Dale, V.H. Comparing large, infrequent disturbances: What have we learned? Ecosystems 1998, 1, 493–496. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Tecco, P.A.; Díaz, S. Plant Invasions in Undisturbed Ecosystems: The Triggering Attribute Approach. J. Veg. Sci. 2005, 16, 723–728. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Gandaseca, S.; Abd Wahab, N.L.; Pazi, A.M.; Rosli, N.; Zaki, P.H. Comparison of water quality status of undisturbed and disturbed mangrove forest at Awat-Awat Lawas Sarawak. Open J. For. 2016, 6, 14–18. [Google Scholar] [CrossRef]
- Jamaluddin, J.; Azhar, B.; Chong, P. Estimating Wildlife Occurrence in Peninsular Malaysia Using GWR4. Malays. For. 2018, 81, 55–63. [Google Scholar]
- MacArthur, R.H. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, H.S.; Fischer, A. Host tree utilization by epiphytic orchids in different land-use intensities in Kathmandu valley, Nepal. Plant Ecol. 2012, 213, 1393–1412. [Google Scholar] [CrossRef]
- Preston, F.W. The canonical distribution of commonness and rarity: Part I. Ecology 1962, 43, 185–215. [Google Scholar] [CrossRef]
- Connor, E.F.; McCoy, E.D. Species–Area Relationships. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2002; p. 10. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P.; Larsen, T.H.; Docherty, T.D.; Ansell, F.A.; Hsu, W.W.; Derhé, M.A.; Hamer, K.C.; Wilcove, D.S. Degraded lands worth protecting: The biological importance of Southeast Asia’s repeatedly logged forests. Proc. R. Soc. B Biol. Sci. 2011, 278, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hohnwald, S. Beiträge zum Mikroklima interandiner Trockentäler Boliviens. Geoökodynamik 1999, 20, 221–229. [Google Scholar]
- Gradstein, S.R. Epiphytes of Tropical Montane Forests—Impact of Deforestation and Climate Change. In The Tropical Mountain Forest: Patterns and Processes in a Biodiversity Hotspot; Gradstein, S.R., Homeier, J., Gansert, D., Eds.; Biodiversity and Ecology Series: Göttingen, Germany, 2008; pp. 51–65. [Google Scholar]
- Gentry, A.H.; Dodson, C.H. Contribution of non-trees to species richness of a tropical rain forest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Wolf, J.H.D. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the northern Andes. Ann. Mo. Bot. Gard. 1993, 80, 928–960. [Google Scholar] [CrossRef]
- Kreft, H.; Köster, N.; Küper, W.; Nieder, J.; Barthlott, W. Diversity and biogeography of vascular epiphytes in Western Amazonia, Yasuni, Ecuador. J. Biogeogr. 2004, 31, 1463–1476. [Google Scholar] [CrossRef]
- Sanford, W.W. The Ecology of Orchids. In The Orchids (Scientific Studies); Withner, C.L., Ed.; Wiley: New York, NY, USA, 1974; p. 100. [Google Scholar]
- Gradstein, S.R.; Churchill, S.P.; Salazar Allen, N. Guide to the bryophytes of tropical America. Mem. N. Y. Bot. Gard. 2001, 86, 577. [Google Scholar]
- Richardson, D.H.S. The Biology of Mosses; Halsted Press: New York, NY, USA, 1981; p. 220. [Google Scholar]
- Alpert, P.; Oechel, W.C. Carbon balance limits the microdistribution of Grimmia laevigata, a desiccation-tolerant plant. Ecology 1985, 66, 660–669. [Google Scholar] [CrossRef]
- Cribb, P.; Butterfield, I. The Genus Pleione; Royal Botanic Gardens: Victoria, Australia, 1999; p. 129. [Google Scholar]
- Zhang, W.; Hu, H.; Zhang, S.B. Divergent adaptive strategies by two co-occurring epiphytic orchids to water stress: Escape or avoidance? Front. Plant Sci. 2016, 7, 588. [Google Scholar] [CrossRef]
- Stratton, L.; Goldstein, G.; Meinzer, F.C. Stem water storage capacity and efficiency of water transport: Their functional significance in a Hawaiian dry forest. Plant Cell Environ. 2000, 23, 99–106. [Google Scholar] [CrossRef]
- Köster, N.; Nieder, J.; Barthlott, W. Effect of host tree traits on epiphyte diversity in natural and anthropogenic habitats in Ecuador. Biotropica 2011, 43, 685–694. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Fischer, H.S. Micro-site conditions of epiphytic orchids in a human impact gradient in Kathmandu valley, Nepal. J. Mt. Sci. 2012, 9, 331–342. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Lobakova, E.S.; Kolomeitseva, G.L.; Netrusov, A.I. Microbiota of the orchid rhizoplane. Microbiology 2001, 70, 492–497. [Google Scholar] [CrossRef]
- Herrera, H.; Fuentes, A.; Soto, J.; Valadares, R.; Arriagada, C. Orchid-Associated Bacteria and Their Plant Growth Promotion Capabilities. In Orchids Phytochemistry, Biology and Horticulture; Merillon, J.M., Kodja, H., Eds.; Reference Series in Phytochemistry; Springer: New York, NY, USA, 2021; p. 26. [Google Scholar] [CrossRef]
- Bates, B.; Kundzewicz, Z.; Wu, S. Climate Change and Water; Intergovernmental Panel on Climate Change Secretariat: Geneva, Switzerland, 2008; p. 210. [Google Scholar]
- Barthlott, W.; Schmit-Neuerburg, V.; Nieder, J.; Engwald, S. Diversity and abundance of vascular epiphytes: A comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecol. 2001, 152, 145–156. [Google Scholar] [CrossRef]
- Krömer, T.; Gradstein, S.R. Species richness of vascular epiphytes in two primary forests and fallows in the Bolivian Andes. Selbyana 2003, 24, 190–195. [Google Scholar]
- Acebey, A.; Gradstein, S.R.; Krömer, T. Species richness and habitat diversification of corticolous bryophytes in submontane rain forest and fallows of Bolivia. J. Trop. Ecol. 2003, 19, 9–18. [Google Scholar] [CrossRef]
- Drehwald, U. Biomonitoring of disturbance in neotropical rainforests using bryophytes as indicators. J. Hattori Bot. Lab. 2005, 97, 117–126. [Google Scholar] [CrossRef]
- Holz, I.; Gradstein, S.R. Phytogeography of the bryophyte floras of oak forests and pâramo of the Cordillera de Talamanca, Costa Rica. J. Biogeogr. 2005, 32, 1591–1609. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Masino, S.A.; Faison, E.K. Intact forests in the United States: Proforestation mitigates climate change and serves the greatest good. Front. For. Glob. Change 2019, 2, 27. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F. Forest-climate interactions in fragmented tropical landscapes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.D. The microevolutionary consequences of climate change. Trends Ecol. Evol. 1990, 5, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Feng, C.L.; Luo, Y.B.; Chen, B.S.; Wang, Z.S.; Gu, H.Y. Potential challenges of climate change to orchid conservation in a wild orchid hotspot in southwestern China. Bot. Rev. 2010, 76, 174–192. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. Parasitic plants: Parallels and contrasts with herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef]
- Crain, B.J.; Tremblay, R.L. Update on the distribution of Lepanthes caritensis, a rare Puerto Rican endemic orchid. Endanger. Species Res. 2012, 18, 89–94. [Google Scholar] [CrossRef]
- Merwin, M.C.; Rentmeester, S.A.; Nadkarni, N.M. The influence of host tree species on the distribution of epiphytic bromeliads in experimental monospecific plantations, La Selva, Costa Rica. Biotropica 2003, 35, 37–47. [Google Scholar] [CrossRef]
- Johansson, D. Ecology of vascular epiphytes in West African rain forests. Acta Phytogeogr. Suec. 1974, 59, 1–129. Available online: https://www.diva-portal.org/smash/get/diva2:565496/FULLTEXT01.pdf (accessed on 12 May 2020).
- Frei, J.K.; Dodson, C.H. The chemical effect of certain bark substrates on the germination and early growth of epiphytic orchids. Bull. Torrey Bot. Club 1972, 99, 301–307. [Google Scholar] [CrossRef]
- Laube, S.; Zotz, G. Neither host-specific nor random: Vascular epiphytes on three tree species in a Panamanian lowland forest. Ann. Bot. 2006, 97, 1103–1114. [Google Scholar] [CrossRef]
- Zotz, G. Johansson revisited: The spatial structure of epiphyte assemblages. J. Veg. Sci. 2007, 18, 123–130. [Google Scholar] [CrossRef]
- Annaselvam, J.; Parthasarathy, N. Diversity and distribution of herbaceous vascular epiphytes in a tropical evergreen forest at Varagalaiar, Western Ghats, India. Biodivers. Conserv. 2001, 10, 317–329. [Google Scholar] [CrossRef]
- Mucunguzi, P. Diversity and distribution of epiphytic orchids in Kibale National Park, Uganda. Selbyana 2008, 29, 217–225. [Google Scholar]
- Ling, C.Y.; Sang, J. Diversity of Orchidaceae from Murum Dam, Belaga, Sarawak, Borneo. Sibbaldia 2018, 16, 67–85. Available online: https://core.ac.uk/download/pdf/267015996.pdf (accessed on 3 January 2020).
- Besi, E.E.; Nikong, D.; Pungga, R.S.; Go, R. Wild orchid diversity of highland forest in the Heart of Borneo: Long Banga and Tama Abu, Sarawak. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 125–135. [Google Scholar] [CrossRef]
- Tupac Otero, J.; Aragón, S.; Ackerman, J.D. Site variation in spatial aggregation and phorophyte preference in Psychilis monensis (Orchidaceae). Biotropica 2007, 39, 227–231. [Google Scholar] [CrossRef]
- de la Rosa-Manzano, E.; Andrade, J.L.; Zotz, G.; Reyes-García, C. Epiphytic orchids in tropical dry forests of Yucatan, Mexico–Species occurrence, abundance and correlations with host tree characteristics and environmental conditions. Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 100–109. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Olmsted, I.C. Host tree utilization by vascular epiphytes in a seasonally inundated forest (Tintal) in Mexico. Biotropica 1992, 24, 402–407. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A. Distribution pattern of the epiphytic orchid Rhynchostylis retusa under strong human influence in Kathmandu valley, Nepal. Bot. Orient. J. Plant Sci. 2011, 8, 90–99. [Google Scholar] [CrossRef]
- Callaway, R.M.; Reinhart, K.O.; Tucker, S.C.; Pennings, S.C. Effects of epiphytic lichens on host preference of the vascular epiphyte Tillandsia usneoides. Oikos 2001, 94, 433–441. [Google Scholar] [CrossRef]
- Watthana, S.; Pedersen, H.Æ. Phorophyte diversity, substrate requirements and fruit set in Dendrobium scabrilingue Lindl. (Asparagales: Orchidaceae): Basic observations for re-introduction experiments. Trop. Nat. Hist. 2008, 8, 135–142. Available online: https://li01.tci-thaijo.org/index.php/tnh/article/view/102971 (accessed on 15 January 2020).
- Ibisch, P. Neotropische Epiphytendiversität—Das Beispiel Bolivien; Galunder Verlag: Wiehl, Germany, 1996; p. 357. [Google Scholar]
- Wolseley, P.A.; Aguirre-Hudson, B. The ecology and distribution of lichens in tropical deciduous and evergreen forests of northern Thailand. J. Biogeogr. 1997, 24, 327–343. [Google Scholar] [CrossRef]
- Adhikari, Y.P.; Fischer, A.; Pauleit, S. Sustainable conservation perspectives for epiphytic orchids in the central Himalayas, Nepal. Appl. Ecol. Environ. Res. 2015, 13, 753–767. [Google Scholar] [CrossRef]
- Ong, P.T. Limestone orchids of Peninsular Malaysia. Orchid Rev. 2015, 123, 28–36. [Google Scholar]
- Porembski, S. Epiphytic orchids on arborescent Velloziaceae and Cyperaceae: Extremes of phorophyte specialisation. Nord. J. Bot. 2003, 23, 505–512. [Google Scholar] [CrossRef]
- Morris, B. The epiphytic orchids of the Shire Highlands, Malawi. Proc. Linn. Soc. Lond. 1968, 179, 51–66. [Google Scholar] [CrossRef]
- La Croix, I.F.; La Croix, E.A.S.; La Croix, T.M. Orchids of Malawi: The Epiphytic and Terrestrial Orchids from South and East Central Africa; Taylor & Francis: Rotterdam, The Netherlands, 1991; p. 358. [Google Scholar]
- Matias, L.Q.; Braga, P.I.S.; Freire, A.G. Reproductive biology of Constantia cipoensis Porto & Brade (Orchidaceae), an endemic species from the Serra do Cipo, Minas Gerais. Rev. Bras. De Botânica 1996, 19, 119–125. [Google Scholar]
- da Mota, R.C.; de Barros, F.; Stehmann, J.R. Two new species of Orchidaceae from Brazil: Bulbophyllum carassense and Lepanthopsis vellozicola. Novon J. Bot. Nomencl. 2009, 19, 380–387. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).