
Morphological, Physiological, and Biochemical Composition of Mulberry (Morus spp.) under Drought Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Evaluation of Growth
2.3.1. Morphological and Biomass Indicators
2.3.2. Photosynthetic and Chlorophyll Fluorescence
2.3.3. Biochemical Indices
2.4. Statistical Analysis
3. Results
3.1. Effects of Drought Stress on Photosynthetic Parameters
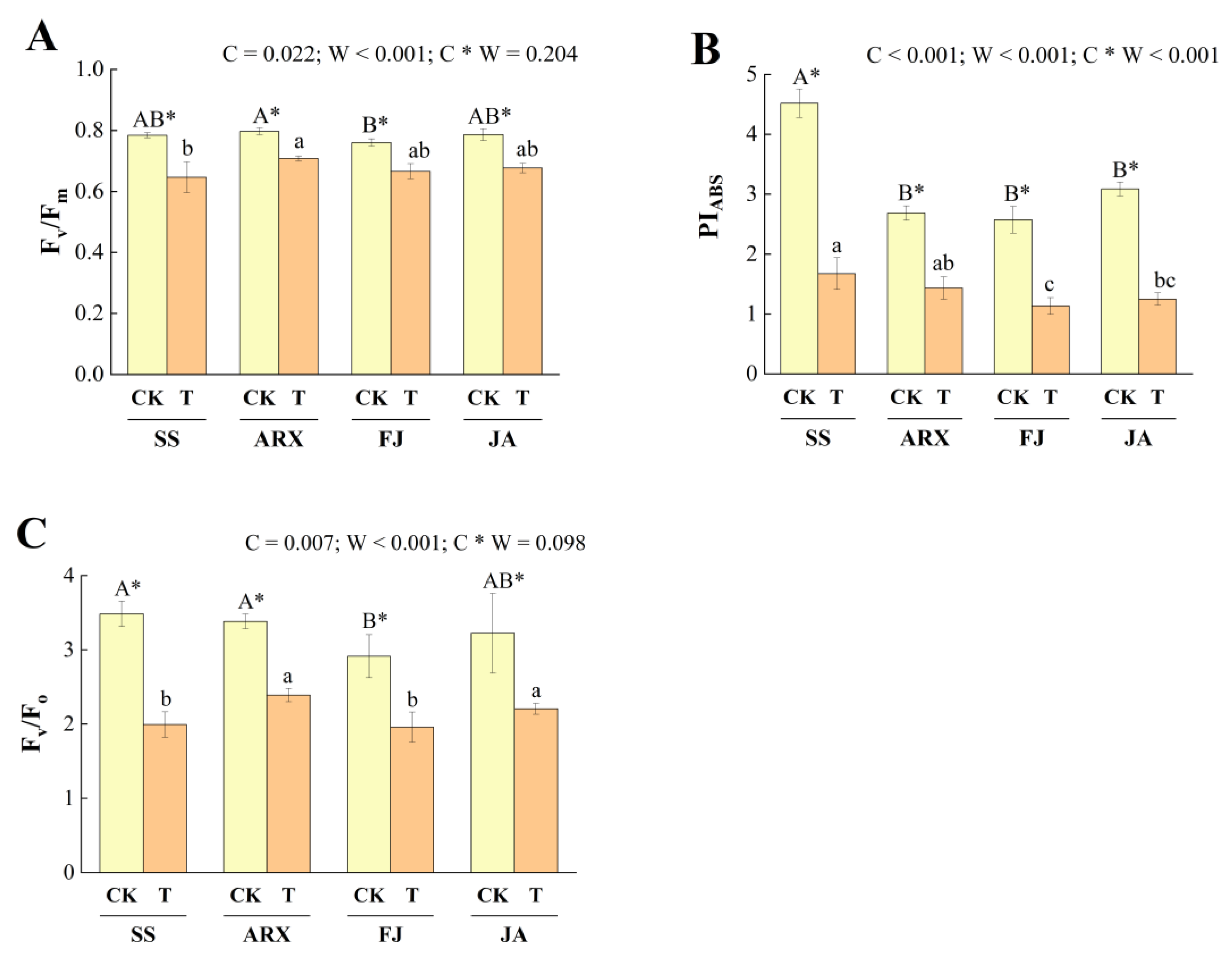
3.2. Effects of Drought Stress on Fluorescence Parameters
3.3. Effects of Drought Stress on Biochemical Composition
3.4. Effects of Drought Stress on Biomass
3.5. Correlation Coefficients of Growth Morphology with Photosynthesis, Fluorescence, Biochemical Index, and Biomass
3.6. Principal Components Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mukarram, M.; Choudhary, S.; Kurjak, D.; Petek, A.; Khan, M.M.A. Drought: Sensing, signaling, effects and tolerance in higher plants. Physiol. Plant. 2021, 172, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, Y.; Zhu, J.K. Thriving under stress: How plants balance growth and the stress response. Dev. Cell. 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Chen, S. Response mechanism of plants to drought stress. Horticulture 2021, 7, 50. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Wang, J.; Wu, X.; Ma, S.; Xu, Z.; Zhou, T.; Xu, N.; Tang, X.; An, B. Increased CO2 concentrations increasing water use efficiency and improvement PSII function of mulberry seedling leaves under drought stress. J. Plant Interact. 2019, 14, 213–223. [Google Scholar] [CrossRef]
- Zhang, H.H.; Xu, N.; Sui, X.; Zhong, H.X.; Yin, Z.P.; Li, X.; Sun, G.Y. Photosynthesis response to drought stress in leaves of two alfalfa (Medicago sativa) varieties. Int. J. Agric. Biol. 2018, 20, 1012–1020. [Google Scholar]
- Liu, L.; Cao, X.; Zhai, Z.; Ma, S.; Tian, Y.; Cheng, J. Direct evidence of drought stress memory in mulberry from a physiological perspective: Antioxidative, osmotic and phytohormonal regulations. Plant Physiol. Biochem. 2022, 186, 76–87. [Google Scholar] [CrossRef]
- Reddy, K.S.; Sekhar, K.M.; Sreeharsha, R.V.; Reddy, A.R. Hydraulic dynamics and photosynthetic performance facilitate rapid screening of field grown mulberry (Morus spp.) genotypes for drought tolerance. Environ. Exp. Bot. 2019, 157, 320–330. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T.; Muller, B. The physiological basis of drought tolerance in crop plants: A scenario-dependent probabilistic approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Zhuang, J.; Wang, Y.; Chi, Y.; Zhou, L.; Chen, J.; Zhou, W.; Ding, J.; Song, J.; Zhao, N. Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits. PeerJ 2020, 8, e10046. [Google Scholar] [CrossRef]
- Mahmood, T.; Abdullah, M.; Ahmar, S.; Yasir, M.; Iqbal, M.S.; Yasir, M.; Mora-Poblete, F.; Ur Rehman, S.; Ahmed, S.; Rana, R.M.; et al. Incredible role of osmotic adjustment in grain yield sustainability under water scarcity conditions in wheat (Triticum aestivum L.). Plants 2020, 9, 1208. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.Y.; Beckles, D.M. Dynamic changes in the starch-sugar interconversion within plant source and sink tissues promote a better abiotic stress response. J. Plant Physiol. 2019, 234, 80–93. [Google Scholar] [CrossRef]
- Martínez-Noël, G.M.; Tognetti, J.A. Sugar signaling under abiotic stress in plants. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: New York, NY, USA, 2018; pp. 397–406. [Google Scholar] [CrossRef]
- Wen, P.; Hu, T.G.; Linhardt, R.J.; Liao, S.T.; Wu, H.; Zou, Y.X. Mulberry: A review of bioactive compounds and advanced processing technology. Trends Food Sci. Technol. 2019, 83, 138–158. [Google Scholar] [CrossRef]
- Dhiman, S.; Kumar, V.; Mehta, C.M.; Gat, Y.; Kaur, S. Bioactive compounds, health benefits and utilization of Morus spp.—A comprehensive review. J. Hortic. Sci. Biotechnol. 2020, 95, 8–18. [Google Scholar] [CrossRef]
- Awasthi, A.K.; Nagaraja, G.M.; Naik, G.V.; Kanginakudru, S.; Thangavelu, K.; Nagaraju, J. Genetic diversity and relationships in mulberry (genus Morus) as revealed by RAPD and ISSR marker assays. BMC Genet. 2004, 5, 1. [Google Scholar] [CrossRef]
- Sarkar, T.; Mogili, T.; Sivaprasad, V. Improvement of abiotic stress adaptive traits in mulberry (Morus spp.): An update on biotechnological interventions. 3 Biotech. 2017, 7, 214. [Google Scholar] [CrossRef]
- Lu, L.; Tang, Y.; Xie, J.-S.; Yuan, Y.L. The role of marginal agricultural land-based mulberry planting in biomass energy production. Renew. Energy 2009, 34, 1789–1794. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, Y.; Yin, Y.; Zhu, X.; Zhu, W.; Zhou, Y. Comparison of soil bacterial community and functional characteristics following afforestation in the semi-arid areas. PeerJ 2019, 7, e7141. [Google Scholar] [CrossRef]
- Reddy, K.S.; Sekhar, K.M.; Reddy, A.R. Genotypic variation in tolerance to drought stress is highly coordinated with hydraulic conductivity-photosynthesis interplay and aquaporin expression in field-grown mulberry (Morus spp.). Tree Physiol. 2017, 37, 926–937. [Google Scholar] [CrossRef]
- Cao, X.; Shen, Q.; Liu, L.; Cheng, J. Relationships of growth, stable carbon isotope composition and anatomical properties of leaf and xylem in seven mulberry cultivars: A hint towards drought tolerance. Plant Biol. 2020, 22, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvent. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Rukmangada, M.S.; Sumathy, R.; Kruthika, H.S.; Naik, V.G. Mulberry (Morus spp.) growth analysis by morpho-physiological and biochemical components for crop productivity enhancement. Sci. Hortic. 2020, 259, 108819. [Google Scholar] [CrossRef]
- Lei, P.; Liu, Z.; Li, J.; Jin, G.; Xu, L.; Ji, X.; Zhao, X.; Tao, L.; Meng, F. Integration of the Physiology, Transcriptome and Proteome Reveals the Molecular Mechanism of Drought Tolerance in Cupressusgigantea. Forests 2022, 13, 401. [Google Scholar] [CrossRef]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. J. Agron. Crop Sci. 2018, 204, 505–514. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop. J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Chen, H.; Huang, J.; Zhang, Y.; Qi, M.; Liu, Y.; Li, T. Photosynthetic response mechanism of soil salinity-induced cross-tolerance to subsequent drought stress in tomato plants. Plants 2020, 9, 363. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Li, P.D.; Zhu, Y.F.; Song, X.L.; Song, F.P. Negative effects of long-term moderate salinity and short-term drought stress on the photosynthetic performance of Hybrid Pennisetum. Plant Physiol. Biochem. 2020, 155, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.X.; Li, J.; Bi, W.S.; Zuo, S.Y.; Li, L.J.; Li, W.L.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; Van der Tol, C.; Flexas, J.; Pfündel, E.E.; Berry, J.A.; Moreno, J.; Frankenberg, C. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. An integrative approach to photoinhibition and photoprotection of photosynthesis. Environ. Exp. Bot. 2018, 154, 1–3. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazar, D.; Kromdijk, J.; Govindjee. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Alemu, S.T. Photosynthesis limiting stresses under climate change scenarios and role of chlorophyll fluorescence: A review article. Cogent Food Agric. 2020, 6, 1785136. [Google Scholar] [CrossRef]
- Rosanna, D.; Martina, P.; Chiara, C.; Giacomo, G.; Riccardo, M.; Filippo, B. Chlorophyll a fluorescence analysis along a vertical gradient of the crown in a poplar (Oxford clone) subjected to ozone and water stress. Tree Physiol. 2012, 32, 8. [Google Scholar] [CrossRef]
- Force, L.; Critchley, C.; Rensen, J.J.S.V. New fluorescence parameters for monitoring photosynthesis in plants. Photosynth. Res. 2003, 78, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Silva, P.E.; Sanglard, L.M.V.P.; Ávila, R.T.; Morais, L.E.; Martins, S.C.V.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R.; et al. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef]
- Živčák, M.; Brestič, M.; Olšovská, K.; Slamka, P. Performance index as a sensitive indicator of water stress in Triticum aestivum L. Plant Soil Environ. 2008, 54, 133–139. [Google Scholar] [CrossRef]
- Li, Y.; Fang, A.; Zhang, T.; Zhang, S.; Zhu, W.; Zhou, Y. Exogenous Calcium Improves Photosynthetic Capacity of Pinus sylvestris var. mongolica under Drought. Forests 2022, 13, 2155. [Google Scholar] [CrossRef]
- Strasser, B.J.; Strasser, R.J. Measuring Fast Fluorescence Transients to Address Environmental Questions: The JIP-Test. Photosynth. Light Biosph. 1995, 5, 977–980. [Google Scholar] [CrossRef]
- Thomas, F.M.; Yu, R.; Schaefer, P.; Zhang, X.; Lang, P. How diverse are Populus “diversifolia” leaves? Linking leaf morphology to ecophysiological and stand variables along water supply and salinity gradients. Flora 2017, 233, 68–78. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of plant responses to water stress and related genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Pellis, A.; Laureysens, I.; Ceulemans, R. Growth and production of a short rotation coppice culture of poplar I. Clonal differences in leaf characteristics in relation to biomass production. Biomass Bioenergy 2004, 27, 9–19. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Connections between abscission, dehiscence, pathogen defense, drought tolerance, and senescence. Plant Sci. 2019, 284, 25–29. [Google Scholar] [CrossRef]
- Bhusal, N.; Kim, H.S.; Han, S.G.; Yoon, T.M. Photosynthetic traits and plant-water relations of two apple cultivars grown as bi-leader trees under long-term waterlogging conditions. Environ. Exp. Bot. 2020, 176, 104111. [Google Scholar] [CrossRef]
- Marron, N.; Villar, M.; Dreyer, E.; Delay, D.; Boudouresque, E.; Petit, J.-M.; Delmotte, F.M.; Guehl, J.-M.; Brignolas, F. Diversity of leaf traits related to productivity in 31 Populus deltoides × Populus nigra clones. Tree Physiol. 2005, 25, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.L.; Li, D.H.; Wang, L.H.; Ding, X.; Zhang, Y.X.; Gao, Y.; Zhang, X.R. Morpho-anatomical and physiological responses to waterlogging of sesame (Sesamum indicum L.). Plant Sci. 2013, 208, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Soluble Sugar (mg·g−1) | Starch (mg·g−1) | NSC (mg·g−1) | Chl (a + b) (mg·g−1) | |
|---|---|---|---|---|---|
| SS | CK | 68.03 ± 1.02 B | 86.65 ± 1.16 B * | 154.68 ± 1.68 A * | 2.40 ± 0.04 A* |
| T | 71.43 ± 0.43 c | 49.15 ± 1.03 c | 120.58 ± 0.76 c | 1.54 ± 0.10 a | |
| ARX | CK | 48.28 ± 0.44 C * | 75.51 ± 2.17 C | 123.79 ± 2.43 C * | 2.11 ± 0.03 B * |
| T | 54.81 ± 1.13 d | 81.33 ± 1.63 d | 136.14 ± 2.46 ab | 1.69 ± 0.07 a | |
| FJ | CK | 69.16 ± 0.56 B * | 61.02 ± 1.31 B * | 130.18 ± 1.57 C | 1.83 ± 0.09 C * |
| T | 75.38 ± 0.49 b | 51.79 ± 1.82 b | 127.18 ± 2.19 bc | 1.47 ± 0.01 a | |
| JA | CK | 77.44 ± 1.96 A * | 64.24 ± 0.88 A * | 141.68 ± 1.73 B | 1.98 ± 0.12 BC * |
| T | 90.25 ± 0.80 a | 49.70 ± 2.41 a | 139.94 ± 3.13 a | 1.43 ± 0.10 a | |
| C | * | * | * | * | |
| W | * | * | * | * | |
| C × W | * | * | * | * | |
| Genotypes | Morphology Traits | Distribution of Biomass | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BL | TLA | SLA | DLW | DBW | DRW | TB | RCR | ||
| SS | CK | 78 ± 1.15 A | 1034.93 ± 54.24 B | 125.89 ± 1.63 B | 8.21 ± 0.33 B | 7.41 ± 0.53 A | 51.83 ± 2.5 A | 65.01 ± 3.31 A | 3.23 ± 0.06 B |
| T | 58 ± 1.15 a | 415.08 ± 16.12 b | 127.25 ± 5.33 a | 3.47 ± 0.3 b | 4.65 ± 0.41 a | 31.25 ± 4.16 a | 39.37 ± 4.87 a | 3.82 ± 0.19 ab | |
| ARX | CK | 44.33 ± 3.18 B | 620.18 ± 11.1 C | 131.97 ± 4.81 B | 4.72 ± 0.24 C | 2.56 ± 0.08 C | 30.25 ± 3.15 C | 37.52 ± 3.47 C | 4.14 ± 0.25 A |
| T | 32.33 ± 1.45 c | 405.42 ± 11.51 b | 111.71 ± 3.04 bc | 3.63 ± 0.13 b | 1.71 ± 0.18 bc | 20.86 ± 1.43 bc | 26.2 ± 1.7 bc | 3.9 ± 0.1 a | |
| FJ | CK | 52 ± 3.46 B | 1137.38 ± 65.15 B | 146.19 ± 2.49 A | 7.8 ± 0.58 B | 4.34 ± 0.36 B | 39.72 ± 2.12 B | 51.86 ± 3.03 B | 3.28 ± 0.07 B |
| T | 39 ± 1.53 b | 586.93 ± 25.06 a | 125.73 ± 5.86 ab | 4.69 ± 0.34 a | 2.86 ± 0.38 b | 27.56 ± 1.54 ab | 35.11 ± 2.25 ab | 3.67 ± 0.14 a | |
| JA | CK | 47 ± 1.73 B | 1402.25 ± 53.91 A | 134.28 ± 2.46 B | 10.44 ± 0.21 A | 4.35 ± 0.19 B | 32.82 ± 0.86 BC | 47.6 ± 1.26 B | 2.22 ± 0.01 C |
| T | 34 ± 2.08 bc | 565.24 ± 27.3 a | 100.89 ± 3.26 c | 5.63 ± 0.44 a | 1.27 ± 0.43 c | 16.75 ± 2 c | 23.65 ± 2.87 c | 2.43 ± 0.02 b | |
| C | * | * | * | * | * | * | * | * | |
| W | * | * | * | * | * | * | * | * | |
| C × W | * | * | * | * | * | ||||
| DLW | DBW | DRW | TB | RCR | |
|---|---|---|---|---|---|
| BL | 0.42 * | 0.95 ** | 0.90 ** | 0.89 ** | -- |
| TLA | 0.97 ** | 0.58 ** | 0.62 ** | 0.72 ** | −0.59 ** |
| SLA | -- | 0.45 * | 0.52 ** | 0.52 ** | -- |
| PIABS | ABS/RC | TRo/RC | Gs | NSC | Chl (a + b) | |
|---|---|---|---|---|---|---|
| SLA | -- | -- | -- | -- | -- | -- |
| BL | 0.43 ** | 0.49 ** | 0.31 * | 0.32 * | -- | 0.62 * |
| TLA | -- | -- | -- | -- | 0.47 * | 0.53 * |
| Genotypes | PC1 63.9% | PC2 22.4% | Species | PC1 66.0% | PC2 14.2% | ||
|---|---|---|---|---|---|---|---|
| Morphology traits | BL | 0.37 | 0.28 | Photosynthetic parameters | Pn | 0.33 | −0.03 |
| TLA | 0.37 | −0.37 | Gs | 0.30 | 0.21 | ||
| SLA | 0.26 | 0.11 | Tr | 0.31 | −0.23 | ||
| Distribution of biomass | DLW | 0.35 | −0.44 | IWUE | −0.13 | 0.63 | |
| DBW | 0.41 | 0.20 | Fluorescence parameters | PIABS | 0.28 | 0.37 | |
| DRW | 0.41 | 0.24 | Fv/Fm | 0.33 | 0.06 | ||
| TB | 0.43 | 0.13 | Fv/Fo | 0.32 | 0.15 | ||
| RCR | −0.11 | 0.68 | Vj | −0.22 | −0.46 | ||
| Mo | −0.25 | −0.08 | |||||
| ABS/RC | −0.30 | 0.24 | |||||
| TRo/RC | −0.32 | 0.18 | |||||
| ETo/RC | 0.20 | 0.20 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Yin, Y.; Zhu, W.; Zhou, Y. Morphological, Physiological, and Biochemical Composition of Mulberry (Morus spp.) under Drought Stress. Forests 2023, 14, 949. https://doi.org/10.3390/f14050949
Sun Z, Yin Y, Zhu W, Zhou Y. Morphological, Physiological, and Biochemical Composition of Mulberry (Morus spp.) under Drought Stress. Forests. 2023; 14(5):949. https://doi.org/10.3390/f14050949
Chicago/Turabian StyleSun, Zhiyu, You Yin, Wenxu Zhu, and Yongbin Zhou. 2023. "Morphological, Physiological, and Biochemical Composition of Mulberry (Morus spp.) under Drought Stress" Forests 14, no. 5: 949. https://doi.org/10.3390/f14050949