
Attraction of Bees to Native and Introduced Willows (Salix spp.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System and Design
2.2. Salix Taxa
2.3. Bee Collection
2.4. Statistical Analyses
3. Results and Discussion
3.1. Bee Diversity
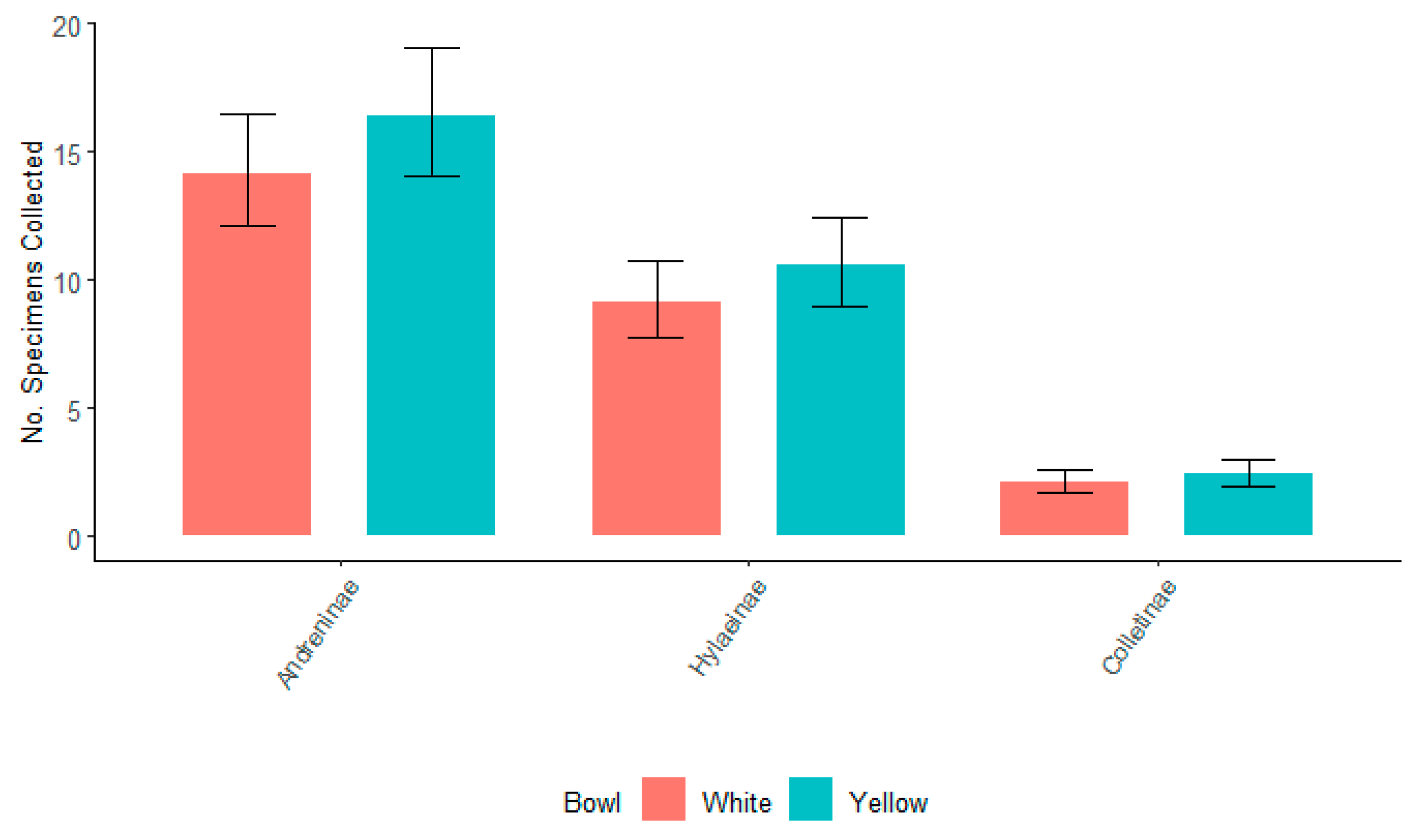
3.2. Bowl Color
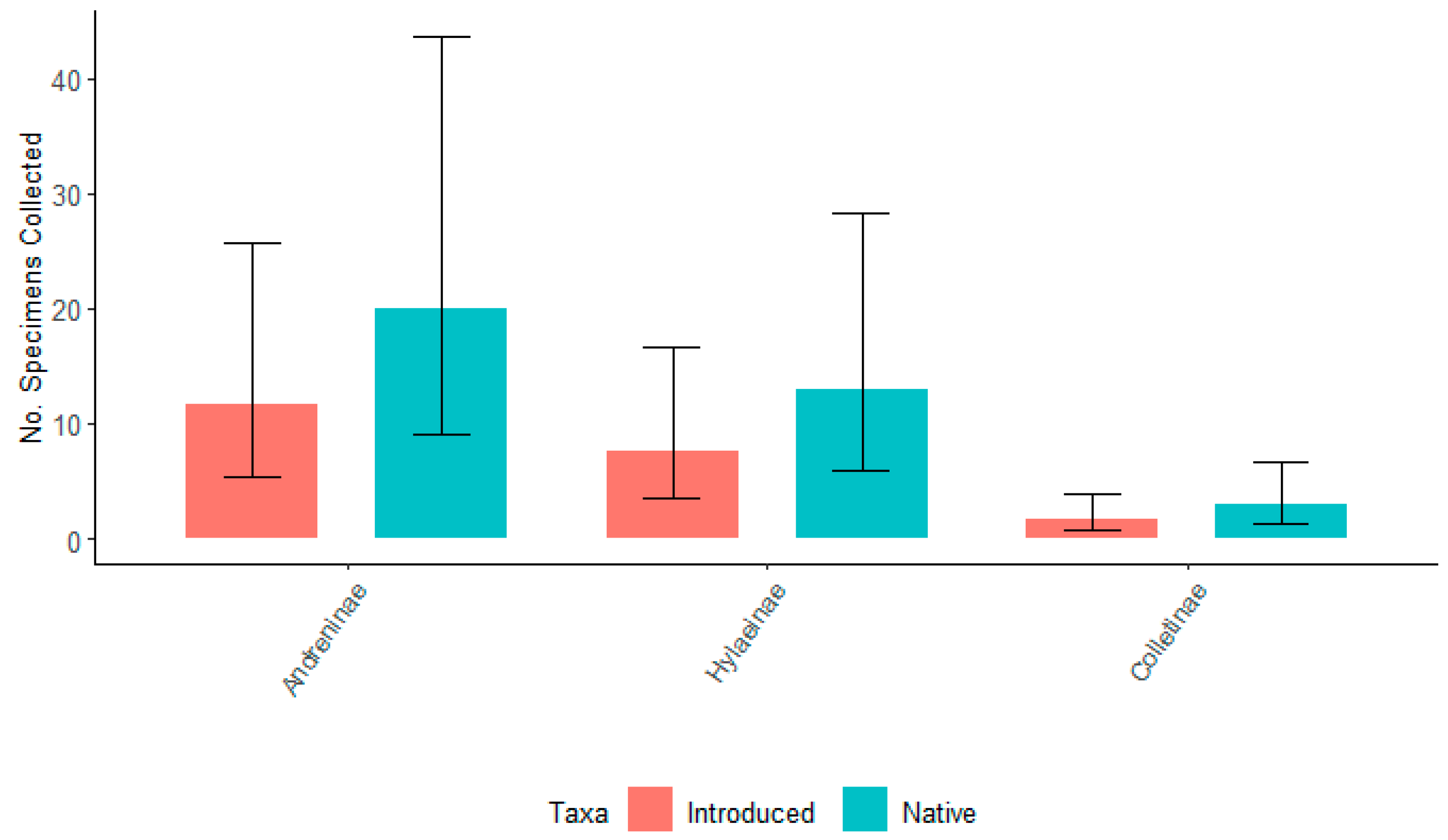
3.3. Total Number of Bees Associated with Native and Introduced Willows
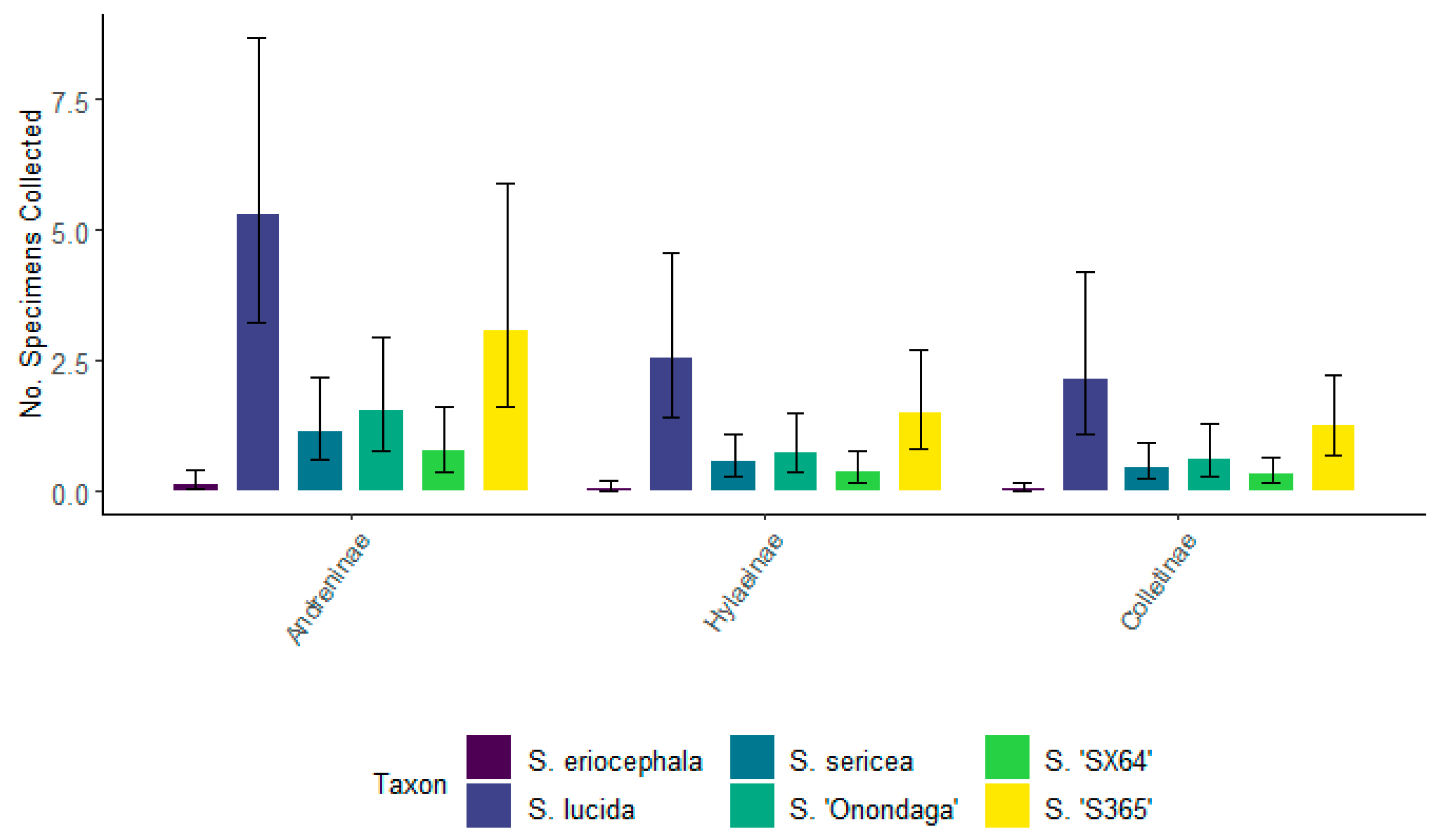
3.4. Total Number of Bees Associated with Each Willow Taxa
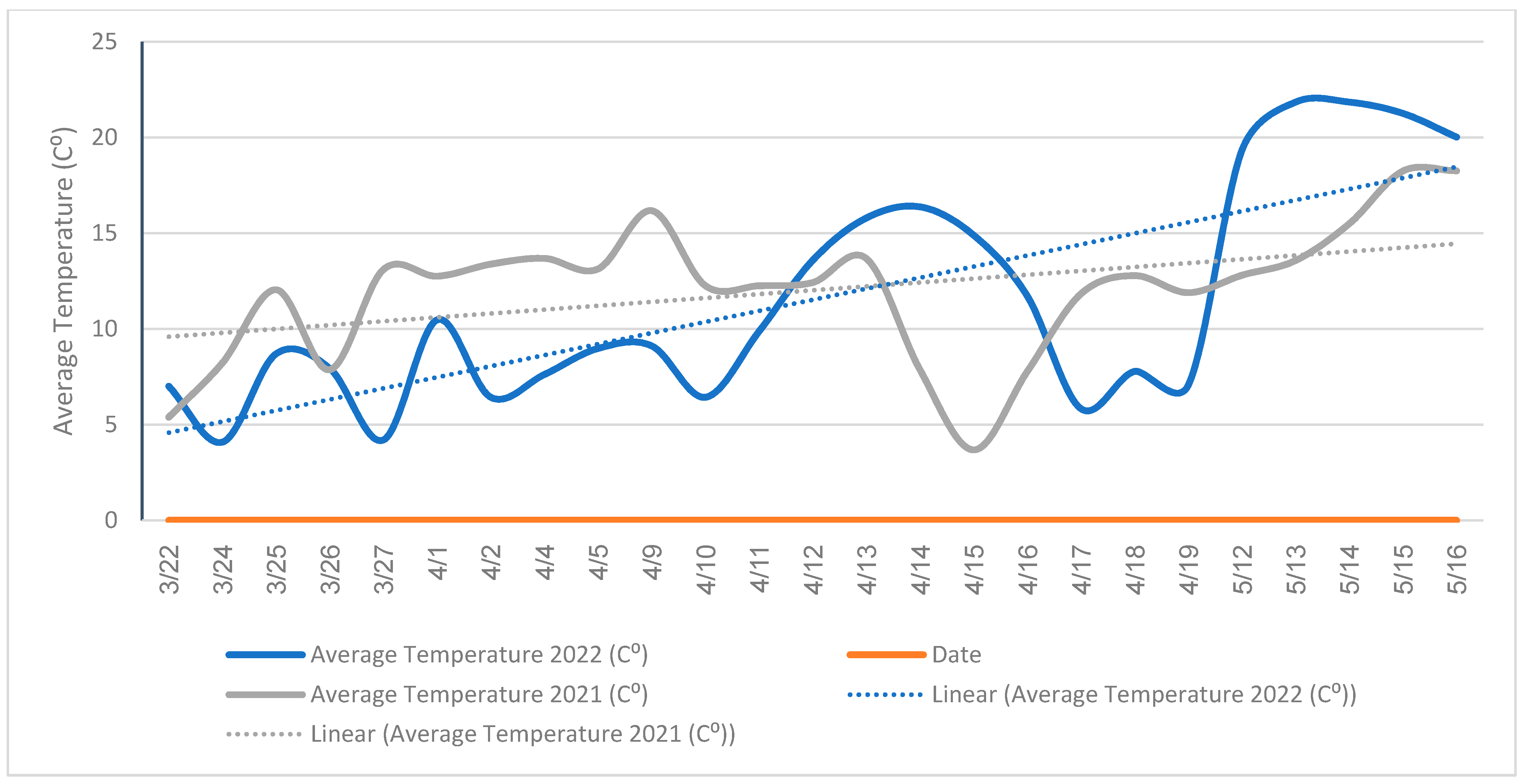
3.5. Overlapping Blooming Periods
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Langholtz, M.; Eaton, L.; Davis, M.; Shedden, M.; Brandt, C.; Volk, T.; Richard, T. Economic comparative advantage of willow biomass in the Northeast USA. Biofuels Bioprod. Biorefin. 2019, 13, 74–85. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Volk, T.A. The characterization of willow (Salix L.) varieties for use in ecological engineering applications: Co-ordination of structure, function, and autecology. Ecol. Eng. 2009, 35, 1178–1189. [Google Scholar] [CrossRef]
- Zumpf, C.; Quinn, J.; Cacho, J.; Grasse, N.; Negri, M.C.; Lee, D. Invertebrate and plant community diversity of an Illinois corn–soybean field with integrated shrub willow bioenergy buffers. Sustainability 2021, 13, 12280. [Google Scholar] [CrossRef]
- Rönnberg-Wästljung, A.C.; Dufour, L.; Gao, J.; Hansson, P.A.; Herrmann, A.; Jebrane, M.; Johansson, A.C.; Kalita, S.; Molinder, R.; Nordh, N.E.; et al. Optimized utilization of Salix—Perspectives for the genetic improvement toward sustainable biofuel value chains. GCB Bioenergy 2022, 14, 1128–1144. [Google Scholar] [CrossRef]
- Reddersen, J. SRC-willow (Salix viminalis) as a resource for flower-visiting insects. Biomass Bioenergy 2001, 20, 171–179. [Google Scholar] [CrossRef]
- Volk, T.A.; Berguson, B.; Daly, C.; Halbleib, M.; Miller, R.; Rials, T.; Abrahamson, L.; Buchman, D.; Buford, M.; Cunningham, M.; et al. Poplar and shrub willow energy crops in the United States: Field trial results from the multiyear regional feedstock partnership and yield potential maps based on the PRISM-ELM Model. GCB Bioenergy 2018, 10, 735–751. [Google Scholar] [CrossRef]
- Frank, J.; Therasme, O.; Volk, T.A.; Brown, T.; Malmsheimer, R.W.; Fortier, M.O.; Eisenbies, M.H.; Ha, H.; Heavey, J. Integrated stochastic life cycle assessment and techno-economic analysis for shrub willow production in the Northeastern United States. Sustainability 2022, 14, 9007. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Weih, M.; Abalos Romero, M.; Charles, J.; Hurst, S.; McIvor, I.; Karp, A.; Trybush, S.; Labrecque, M.; Teodorescu, T.; et al. Salix: Botany and global horticulture. Hortic. Rev. 2008, 34, 447–489. [Google Scholar]
- Rosa, D.; Clausen, J.; Kuzovkina, Y.A. Water quality changes in a short rotation woody crop riparian buffer. Bioenergy Biomass 2017, 107, 370–375. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Schulthess, C.; Zheng, D. Influence of soil chemical and physical characteristics on willow yield in Connecticut. Biomass Bioenergy 2018, 108, 297–306. [Google Scholar] [CrossRef]
- Yang, S.; Volk, T.A.; Fortier, M.O.P. Willow biomass crops are a carbon negative or low-carbon feedstock depending on prior land use and transportation distances to end users. Energies 2020, 13, 4251. [Google Scholar] [CrossRef]
- Tumminello, G.; Volk, T.A.; McArt, S.H.; Fierke, M.K. Maximizing pollinator diversity in willow biomass plantings: A comparison between willow sexes and among pedigrees. Biomass Bioenergy 2018, 117, 124–130. [Google Scholar] [CrossRef]
- Baril, L.M.; Hansen, A.J.; Renkin, R.; Lawrence, R. Songbird response to increased willow (Salix spp.) growth in Yellowstone’s Northern Range. Ecol. Appl. 2011, 21, 2283–2296. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.R.; DeBano, S.J.; Rowland, M.M.; Burrows, S. Feed the bees and shade the streams: Riparian shrubs planted for restoration provide forage for native bees. Restor. Ecol. 2022, 30, e13525. [Google Scholar] [CrossRef]
- Dauber, J.; Jones, M.B.; Stout, J.C. The impact of biomass crop cultivation on temperate biodiversity. GCB Bioenergy 2010, 2, 289–309. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Verheyen, K.; Buggenhout, M.; Vangansbeke, P.; De Dobbelaere, A.; Verdonckt, P.; Bonte, D. Potential of short rotation coppice plantations to reinforce functional biodiversity in agricultural landscapes. Biomass Bioenergy 2014, 67, 435–442. [Google Scholar] [CrossRef]
- Rowe, R.L.; Goulson, D.; Doncaster, C.P.; Clarke, D.J.; Taylor, G.; Hanley, M.E. Evaluating ecosystem processes in willow short rotation coppice bioenergy plantations. GCB Bioenergy 2013, 5, 257–266. [Google Scholar] [CrossRef]
- Rowe, R.L.; Hanley, M.E.; Goulson, D.; Clarke, D.J.; Doncaster, C.P.; Taylor, G. Potential benefits of commercial willow Short Rotation Coppice (SRC) for farm-scale plant and invertebrate communities in the agri-environment. Biomass Bioenergy 2011, 35, 325–336. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.; Ostaff, D.; Ascher, J. Bee foraging preferences on three willow (Salix) species: Effects of species, plant sex, sampling day and time of day. Ann. Appl. Biol. 2020, 177, 333–345. [Google Scholar] [CrossRef]
- Kollberg, I.; Weih, M.; Glynn, C. The effect of willow diversity on insect herbivory and predation. Agric. For. Entomol. 2022, 24, 27–39. [Google Scholar] [CrossRef]
- Prđun, S.; Svečnjak, L.; Valentić, M.; Marijanović, Z.; Jerković, I. Characterization of bee pollen: Physico-chemical properties, headspace composition and FTIR spectral profiles. Foods 2021, 10, 2103. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.J.; Keefover-Ring, K.; Park, Y.L.; Wimp, G.; Grady, J.; DiFazio, S.P. Characterization of Salix nigra floral insect community and activity of three native Andrena bees. Ecol. Evol. 2021, 11, 4688–4700. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.; Marquardt, M.; Babucke, K.; Heuel, K.C.; Ayasse, M.; Dötterl, S.; Galizia, C.G. Neural and behavioural responses of the pollen-specialist bee Andrena vaga to Salix odors. J. Exp. Biol. 2021, 224, jeb242166. [Google Scholar] [CrossRef] [PubMed]
- Dötterl, S.; Glück, U.; Jürgens, A.; Woodring, J.; Aas, G. Floral reward, advertisement, and attractiveness to honeybees in dioecious Salix caprea. PLoS ONE 2014, 9, e93421. [Google Scholar] [CrossRef]
- Füssel, U.; Dötterl, S.; Jürgens, A.; Aas, G. Inter-and intraspecific variation in floral scent in the genus Salix and its implication for pollination. J. Chem. Ecol. 2007, 33, 749–765. [Google Scholar] [CrossRef]
- Konatowska, M.; Rutkowski, P.; Wendzonka, J. The impact of willow flowering time on species composition and the number of Apoidea pollinators. J. Biosci. Med. 2021, 9, 89–100. [Google Scholar]
- Isaacs, R.; Tuell, J.K.; Fiedler, A.K.; Gardiner, M.; Landis, D.A. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Jones, A.T.; Hayes, M.J.; Sackville Hamilton, N.R. The effect of provenance on the performance of Crataegus monogyna in hedges. J. Appl. Ecol. 2001, 38, 952–962. [Google Scholar] [CrossRef]
- Baisden, E.C.; Tallamy, D.W.; Narango, D.L.; Boyle, E. Do cultivars of native plants support insect herbivores? HortTechnology 2018, 28, 596–606. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Lau, J.A.; Hambäck, P.A. Community heterogeneity and the evolution of interactions between plants and insect herbivores. Q. Rev. Biol. 2006, 81, 349–376. [Google Scholar] [CrossRef]
- Burghardt, K.T.; Tallamy, D.W. Not all non-natives are equally unequal: Reductions in herbivore β-diversity depend on phylogenetic similarity to native plant community. Ecol. Lett. 2015, 18, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Gouker, F.E.; Fabio, E.S.; Serapiglia, M.J.; Smart, L.B. Yield and biomass quality of shrub willow hybrids in differing rotation lengths and spacing designs. Biomass Bioenergy 2021, 146, 105977. [Google Scholar] [CrossRef]
- Montes, F.; Fabio, E.S.; Smart, L.B.; Kemanian, A.A. A semi-commercial case study of willow biomass production in the northeastern USA. Agron. J. 2021, 113, 1287–1302. [Google Scholar] [CrossRef]
- Fletcher, R.J., Jr.; Robertson, B.A.; Evans, J.; Doran, P.J.; Alavalapati, J.R.; Schemske, D.W. Biodiversity conservation in the era of biofuels: Risks and opportunities. Front. Ecol. Environ. 2011, 9, 161–168. [Google Scholar] [CrossRef]
- Fabio, E.S.; Volk, T.A.; Miller, R.O.; Serapiglia, M.J.; Gauch, H.G.; Van Rees, K.C.; Smart, L.B. Genotype× environment interaction analysis of North American shrub willow yield trials confirms superior performance of triploid hybrids. Bioenergy 2017, 9, 445–459. [Google Scholar] [CrossRef]
- Cameron, K.; Smart, L.; Ballard, B.; Volk, T.; Abrahamson, L. (Eds.) SUNY ESF. 2007a. Salix purpurea ‘Onondaga’. 2007. Available online: https://www.esf.edu/willow/documents/Onondaga.pdf (accessed on 25 March 2023).
- Cameron, K.; Smart, L.; Volk, T.; Abrahamson, L. (Eds.) SUNY ESF. 2007b. Salix miyabeana ‘SX64’. 2007. SUNY ESF. Available online: https://www.esf.edu/willow/documents/SX64.pdf (accessed on 25 March 2023).
- Cameron, K.; Smart, L.; Ballard, B.; Volk, T.; Abrahamson, L. (Eds.) SUNY ESF. 2007c. Salix caprea Hybrid ‘S365’. 2007. SUNY ESF. Available online: https://www.esf.edu/willow/documents/S365.pdf (accessed on 25 March 2023).
- St. Clair, A.L.; Dolezal, A.G.; O’Neal, M.E.; Toth, A.L. Pan traps for tracking honeybee activity-density: A case study in soybeans. Insects 2020, 11, 366. [Google Scholar] [CrossRef]
- Templ, B.; Mózes, E.; Templ, M.; Földesi, R.; Szirák, Á.; Báldi, A.; Kovács-Hostyánszki, A. Habitat-dependency of transect walk and pan trap methods for bee sampling in farmlands. J. Apic. Sci. 2019, 63, 93–115. [Google Scholar] [CrossRef]
- Buffington, M.L.; Garretson, A.; Kula, R.R.; Gates, M.W.; Carpenter, R.; Smith, D.R.; Kula, A.A. Pan trap color preference across Hymenoptera in a forest clearing. Entomol. Exp. Appl. 2021, 169, 298–311. [Google Scholar] [CrossRef]
- Borror, D.J.; White, R.E. A Field Guide to Insects: America North of Mexico; Houghton Mifflin Harcourt: Boston, MA, USA, 1998; Volume 19. [Google Scholar]
- Cadotte, M.W.; Cadotte, I.J.; MacIvor, J.S. The Bees in Your Backyard: A Guide to North America’s Bees: Andrenidae; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Dibble, A.C.; Drummond, F.A.; Averill, A.L.; Bickerman-Martens, K.; Bosworth, S.C.; Bushman, S.L.; Hoshide, A.K.; Leach, M.E.; Skyrm, K.; Venturini, E.; et al. Bees and Their Habitats in Four New England States. MR448; Maine Agricultural and Forest Experiment Station: Orono, ME, USA, 2018. [Google Scholar]
- Zarrillo, T.A.; Ascher, J.S.; Gibbs, J.; Stoner, K.A. New and noteworthy records of bees (Hymenoptera: Apoidea: Anthophila) for Connecticut. J. Kans. Entomol. Soc. 2016, 89, 138–157. [Google Scholar] [CrossRef]
- Wilson, J.S.; Carril, O.M. The Bees in Your Backyard. A Guide to North America’s Bees: Colletidae; Princeton University Press: Princeton, NJ, USA, 2015. [Google Scholar]
- Suni, S.S.; Scott, Z.; Averill, A.; Whiteley, A. Population genetics of wild and managed pollinators: Implications for crop pollination and the genetic integrity of wild bees. Conserv. Genet. 2018, 18, 667–677. [Google Scholar] [CrossRef]
- Berkley, N.A.; Hanley, M.E.; Boden, R.; Owen, R.S.; Holmes, J.H.; Critchley, R.D.; Carroll, K.; Sawyer, D.G.M.; Parmesan, C. Influence of bioenergy crops on pollinator activity varies with crop type and distance. GCB Bioenergy 2018, 10, 960–971. [Google Scholar] [CrossRef]
- Droege, S.; Tepedino, V.J.; LeBuhn, G.; Link, W.; Minckley, R.L.; Chen, Q.; Conrad, C. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Ostaff, D.P.; Mosseler, A.; Johns, R.C.; Javorek, S.; Klymko, J.; Ascher, J.S. Willows (Salix spp.) as pollen and nectar sources for sustaining fruit and berry pollinating insects. Can. J. Plant Sci. 2015, 95, 505–516. [Google Scholar] [CrossRef]
- Filipiak, Z.M.; Denisow, B.; Stawiarz, E.; Filipiak, M. Unravelling the dependence of a wild bee on floral diversity and composition using a feeding experiment. Sci. Total Environ. 2022, 820, 153326. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salix Taxon | Common Name | Native Status | Blooming Period | Blooming Time | Distribution | Pedigree/Provenance | Source | Height | Resistance |
|---|---|---|---|---|---|---|---|---|---|
| S. eriocephala | Heart-leaf willow | Yes | Mid-season | Mid-April | Eastern and Central North America | Unknown | Vermont Willow Nursery, Vermont | 4.57 m–6.09 m | Resistant to pests/disease |
| S. sericea | Silky willow | Yes | Mid-season | Mid-April | Eastern North America | Wild, collected in Connecticut | Wild collected | 2.13 m–3.66 m | Resistant to pests/disease |
| S. lucida | Shining willow | Yes | Late season | Beginning of May | Eastern and Central North America | Unknown | Vermont Willow Nursery, Vermont | 4.00 m–7.50 m | Resistant to pests/disease |
| S. ‘Onondaga’ | Onondaga purple osier | No | Early season | End of March | Introduced, Europe | S. purpurea ‘SH3’ × S. purpurea ‘94002’ | SUNY/Double A Willow (NY) | 4.57 m–6.09 m | Resistant to pests/disease |
| S. ‘SX64’ | SX64 Miyabe willow | No | Mid-season | Beginning of April | Introduced, Asia | Wild collected in Japan | SUNY/Double A Willow (NY) | 4.57 m–6.09 m | Resistant to pests/disease |
| S. ‘S365′ | Hybrid of Goat willow | No | Mid-season | Mid-April | Introduced, Europe | Most likely S. caprea × S. cinerea | SUNY/Double A Willow (NY) | 3.05 m–4.57 m | Resistant to pests/disease |
| Willow Taxon | Hymenoptera Taxon | Count |
|---|---|---|
| Salix eriocephala | Andrenidae (Andreninae) | 226 |
| Colletidae (Hylaeinae) | 169 | |
| Colletidae (Colletinae) | 56 | |
| Halictidae (Halictinae) | 1 | |
| Apidae (Bombus) | 3 | |
| Sphecidae | 26 | |
| Vespidae | 1 | |
| Total | 482 | |
| Salix sericea | Andrenidae (Andreninae) | 270 |
| Colletidae (Hylaeinae) | 145 | |
| Colletidae (Colletinae) | 49 | |
| Halictidae (Halictinae) | 2 | |
| Apidae (Bombus) | 0 | |
| Sphecidae | 17 | |
| Vespidae | 7 | |
| Total | 490 | |
| Salix lucida | Andrenidae (Andreninae) | 329 |
| Colletidae (Hylaeinae) | 134 | |
| Colletidae (Colletinae) | 21 | |
| Halictidae (Halictinae) | 4 | |
| Apidae (Bombus) | 1 | |
| Sphecidae | 6 | |
| Vespidae | 10 | |
| Total | 505 | |
| S. ‘SX64’ | Andrenidae (Andreninae) | 232 |
| Colletidae (Hylaeinae) | 179 | |
| Colletidae (Colletinae) | 37 | |
| Halictidae (Halictinae) | 0 | |
| Apidae (Bombus) | 1 | |
| Sphecidae | 8 | |
| Vespidae | 2 | |
| Total | 459 | |
| S. ‘S365’ | Andrenidae (Andreninae) | 209 |
| Colletidae (Hylaeinae) | 203 | |
| Colletidae (Colletinae) | 27 | |
| Halictidae (Halictinae) | 1 | |
| Apidae (Bombus) | 1 | |
| Sphecidae | 12 | |
| Vespidae | 1 | |
| Total | 454 | |
| S. ‘Onondaga’ | Andrenidae (Andreninae) | 49 |
| Colletidae (Hylaeinae) | 21 | |
| Colletidae (Colletinae) | 49 | |
| Halictidae (Halictinae) | 1 | |
| Apidae (Bombus) | 0 | |
| Sphecidae | 0 | |
| Vespidae | 0 | |
| Total | 120 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grandstaff, G.; Kuzovkina, Y.A.; Legrand, A. Attraction of Bees to Native and Introduced Willows (Salix spp.). Forests 2023, 14, 959. https://doi.org/10.3390/f14050959
Grandstaff G, Kuzovkina YA, Legrand A. Attraction of Bees to Native and Introduced Willows (Salix spp.). Forests. 2023; 14(5):959. https://doi.org/10.3390/f14050959
Chicago/Turabian StyleGrandstaff, Gabrielle, Yulia A. Kuzovkina, and Ana Legrand. 2023. "Attraction of Bees to Native and Introduced Willows (Salix spp.)" Forests 14, no. 5: 959. https://doi.org/10.3390/f14050959
APA StyleGrandstaff, G., Kuzovkina, Y. A., & Legrand, A. (2023). Attraction of Bees to Native and Introduced Willows (Salix spp.). Forests, 14(5), 959. https://doi.org/10.3390/f14050959