Abstract
Spermosphere microorganisms are affected by seed exudates (carbon deposits) and interact with each other around the germinating seeds. The interactions in this region will have an important impact on seed germination, plant growth, and development. Ormosia henryi Prain is a rare and endangered tree species with hardseed in China that generally shows low levels of natural regeneration. However, good levels of natural regeneration were observed under a few mother trees in Guanling County (Guizhou Province, China). Based on this phenomenon: we raise the question: Is the seed germination of Ormosia henryi in high-level natural regeneration areas driven by the spermosphere microbe and what is the driving mechanism? The purpose of this study was to explore the promoting role of spermosphere bacteria in different stages of germination, so as to provide a theoretical basis for screening strong colonization ability spermosphere functional bacteria. Thus, in this study, we designed a germination experiment simulating the natural soil (sterilized and unsterilized) of Guanling County. We used 16S rRNA gene sequencing and widely targeted multiple reaction monitoring (MRM) metabolomic analysis to analyze bacterial microbial diversity and the metabolic profiles of seed exudates at the four stages of seed germination: imbibition, radicle protrusion, radicle elongation, and cotyledon protrusion. The results indicated that some spermosphere bacteria: taxa of Bacillus, Acidobacteriales, Nitrospira, and Ellin6067, and other functional bacteria may play important roles in promoting seed germination depending largely on their functional characteristics and abundance. The critical period for spermosphere bacteria to promote seed germination was at the radicle protrusion stage in the unsterilized soil group, levels of metabolites such as sugars and amino acids of seed exudates were increased in this stage, more respiratory pathways were activated and seed respiratory metabolism was enhanced. Spermosphere bacteria of wild Ormosia henryi in the Guanling area appeared to promote the germination of O. henryi seeds, mainly through activating galactose metabolism and the lysine degradation pathway to provide energy for germination.
1. Introduction
Seed germination is a key step in plant development and agricultural production. The spermosphere is “a microbiologically dynamic zone” of soil within 1~10 mm of the surface of a germinating seed [1]. Many aspects of seed biology, including seed germination, dormancy, storage, and dispersal, are influenced by spermosphere microorganisms [2,3,4]. Germinating seeds release exudates into the spermosphere, providing a suitable environment for microbial growth [5], and playing an important role in the germination of many crop seeds. For example, Bacillus velezensis acts as an antifungal agent in the spermosphere of germinating Zea mays seeds [6]. Acinetobacter sp. and Sphingomonas melonis in the spermosphere and on Oryza sativa seeds promote the germination and disease resistance of O. sativa seeds [7,8]. Methylobacterium oryzae in the spermosphere of tomato (Solanum lycopersicum) seeds plays a role in seed germination and plant growth promotion [9]. In addition, Pseudomonas sp. [10], Massilia sp. [11], Pseudarthrobacter sp., Bacteroidetes sp. [12], and Azospirillum brasilense [13] have been found in the spermosphere of various crops and play a role in promoting seed germination. Spermosphere microorganisms are known to drive the germination of crop seeds to some extent, but few studies have been completed on the microbial species and mechanisms that drive the germination of woody plant seeds under natural conditions.
Previous investigations have reported that spermosphere bacteria can increase seed germination rate and seedling vigor, which may be due to their ability to produce hormones. Bacillus, for example, can produce exogenous gibberellin and reduce the levels of jasmonic acid and abscisic acid to promote the germination of rice seeds [14]. Methylotrophic bacteria that produce hormones, such as cytokinins and indole-3-acetic acid (IAA), are abundant in spinach seeds, seed development, and cotyledon elongation [2]. Enterobacter sp. strain NIASMVII produces IAA in seeds of Psoralea corylifolia, which promotes cell elongation and growth and may increase its germination rate [15]. In addition to participating in hormone synthesis, spermosphere bacteria can also promote seed germination under various abiotic stresses by inducing physiological protection against oxidative damage. For example, abundant Bacillus strains are in the spermosphere of peanut (Arachis hypogaea) radicles, germination, and seedling growth under salt stress [11]. Some spermosphere bacteria, such as Pseudomonas chlororaphis, can also produce antifungal metabolites with spectral activity that can effectively control seed-borne diseases [16].
Metabolomics involves studying and can accurately reflect the pathological and physiological state of organisms. Seed germination requires an energy supply, and at the metabolic level, gibberellin induces the expression of a variety of hydrolytic enzymes to re-activate reserves in aleurone cells, including starch, lipids, and proteins, to sustain embryonic growth [17]. Therefore, this involves various metabolic pathways, such as plant hormone synthesis, glucose metabolism, lipid metabolism, and glutathione metabolism [18]. Many studies have investigated interactions between root exudates and rhizosphere microorganisms [19]; however, few studies have investigated interactions between seed exudates and spermosphere microorganisms. Studying microorganisms in the spermosphere broadens our understanding of the regulatory mechanisms of seed dormancy, germination, and subsequent seedling establishment.
Ormosia henryi Prain is a member of the Papilionoideae. Its seeds are generally hard and need to be soaked in hot water (>90 °C) or scarified with a 95% sulfuric acid solution before sowing to promote seed swelling in artificial cultivation. A previous study reported that only 16% of untreated O. henryi seeds buried in soil for a year germinated [20]. Therefore, a low seed germination rate may be an important limiting factor for the natural regeneration of O. henryi. However, while investigating O. henryi's natural regeneration, we observed that seeds under the mother tree germinated well in some places. This observation prompted us to consider whether spermosphere microorganisms were aiding germination. The spermosphere of O. henryi can promote seed germination, such as Trichoderma and Penicillium sclerotiorum [21]. However, to date, neither the effect of bacteria on the germination of O. henryi seeds nor the mechanism driving seed germination has been investigated.
Based on the great difference in the natural regeneration of wide Ormosia henryi Prain and the hardiness characteristic of seed, which leads us to wonder if it has something to do with the spermosphere soil microorganisms. The purpose of this study was to explore the promoting role of spermosphere bacteria. Thus, we proposed two hypotheses: First of all, we hypothesized that (H1) seed-related microorganisms play an important role in germination driving, especially the spermosphere bacteria, which are more diverse and more powerful than the seed endophytes and are the first to contact the hard seed coat of Ormosia henryi. Therefore, we selected the soil under the mother tree of Ormosia henryi in the Guanling area, which had the best natural regeneration effect, for the seed-sowing experiment. In order to eliminate differences in soil factors, previous studies [22] have proved that a short time of soil autoclave sterilization (1 h) has little effect on soil physical and chemical properties, and can produce a large difference in soil microorganisms, thus affecting seed germination and seedling growth. Therefore, we conducted a short time (1 h) sterilization and unsterilization test on the soil under the mother tree of Ormosia henryi in the Guanling area. This comparative test can produce a great difference in the soil microorganisms, so as to verify the important influence of the spermosphere bacteria in the seed germination process. However, seed exudates will be affected by changes in the soil microbial community [3], we further hypothesized (H2) that germinating seed exudates are different due to changes in the spermosphere microbial community. Through the interaction between spermosphere bacteria and seed exudates, the metabolic pathways of key bacteria involved in seed germination can be explored. Since the seed exudates change rapidly depending on the time of seed germination [23] in order to better understand the interaction mechanism between bacteria and seed exudates, we must study their time-course relationship, that is, the interaction at different stages of germination. We used 16S rRNA gene sequencing to analyze the microbial diversity and the metabolic profiles of seed exudates at the four stages of seed germination: imbibition, radicle protrusion, radicle elongation, and cotyledon protrusion. Correlations between differential bacteria and differential metabolites of seed exudates at different stages of germination were explored to determine metabolic pathways of key differential microorganisms involved in seed germination.
2. Materials and Methods
2.1. Seeds and Soil
In November 2019, mature seeds were collected from a single mother plant of O. henryi in Huishui County, Qiannan Buyi, and Miao Autonomous Prefecture (105°39′3.69″ W, 25°58′24.88″ N) in China, the weight of 1000 seeds was 350 g, and which were stored at 4 °C. The seed sowing substrate was 0~20 cm of soil collected from under the same mother plant of wild O. henryi growing in Guanling County, Guizhou Province (Table 1) (105°26′12″ W, 25°58′30″ E) where abundant naturally regenerated seedlings were observed. Impurities and stones were removed, and then the soil was mixed evenly. The analysis of soil properties was completed by the method of soil agricultural chemistry analysis book [24].

Table 1.
Effects of sterilized (CK) and unsterilized (T) soil on Ormosia henryi seed germination and seedling growth (unpaired t-test).
2.2. Sowing Experiment Using Natural Soil as Medium
Half of the soil was sterilized by the method of high-pressure sterilization (CK), and half was not sterilized (T). The sterilization method used in this study is high-pressure steam sterilization. In the current study, long-term sterilization (4 h) with this method will significantly change the concentration of nitrogen, phosphorus, and potassium in the soil, while short-term autoclave sterilization (within 1 h) has little effect on soil nutrient status [22]. Soil sterilization method: The soil sample was put into a 50 × 70 cm sterilization bag and sterilized in a vertical high-pressure steam sterilization pot. The condition of sterilization was 121 °C (0.1 Mpa) high-temperature sterilization for 60 min and dried for later use. Sterilized and unsterilized soil was placed in seed trays (80 × 35 × 20 cm) and 100 healthy undamaged seeds of uniform size were sown in each tray, with three replicates of each treatment. After sowing, a thin layer of soil was covered, and then the topsoil was covered with pine needles to simulate seeding and germination. Regularly to keep the soil moist and germinate in time.
Soil samples (5 g) containing seed exudates and spermosphere bacteria were collected from the spermosphere at four germination stages: imbibition (I), radicle protrusion (II), radicle elongation (III), and cotyledon protrusion (IV). Each duplicate sample of the same treatment was mixed evenly, labeled, and immediately stored at −80 °C until analysis.
2.3. Index of Seed Germination and Morphological Analysis
The germination rate (seed germination/number of seeds sown) × 100% was calculated for each treatment at stage IV. Determination of seedling growth indicators started two months after stage IV; at that point, the seedling’s age was three months. Seedling height, ground diameter, and root length were measured with vernier calipers. Roots and shoots were then dried at 80 ± 2 °C oven until they reached a constant weight, and the dry weight was recorded.
2.4. Analysis of Spermosphere Bacterial Community
Microbiome analysis was conducted by Majorbio Bio-Technology Company. Ltd. (Shanghai, China). DNA was extracted from the spermosphere soil samples using a FastDNA® Spin Kit for Soil (MP Biomedicals, Santa Ana, CA, USA) as described by Bai et al. [25]. The V3–V4 region of the 16S rDNA was amplified using primer pairs 338F: (ACTCCTACGGGAGGCAGCAG) and 806R: (GGACTACHVGGGTWTCTAAT). The amplified PCR products were purified using an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA). The library was constructed using a NEXTFLEX Rapid DNA-Seq Kit (PerkinElmer, Waltham, MA, USA), and sequenced using Illumina’s Miseq PE300 platform. The quality control software FASTQ (https://maq.sourceforge.net/fastq.shtml, accessed on 23 April 2023) was used to the ensure quality of the sequencing reads, FLASH software (https://ccb.jhu.edu/software/FLASH/, accessed on 23 April 2023) was used for splicing, and UPARSE software (https://drive5.com/uparse/, accessed on 23 April 2023) was used to perform operational taxonomic unit (OTU) clustering on the sequence reads based on a 97% sequence similarity. Chimeric sequences were removed during the OTU clustering process and chloroplast and mitochondrial sequences were removed from all samples.
2.5. Analysis of Spermosphere Soil Samples of Seed Exudates
Metabolomic analysis was conducted by Biotree Bio-Technology Company Ltd. (Shanghai, China). Seed exudates among soil samples were determined by performing ultra-high-performance liquid chromatography. Firstly, the samples were freeze-dried and then grounded (60 Hz, 30 s) before weighing 1000 mg of the sample into a centrifuge tube. To extract the exudates, 3500 mL of methanol: water (volume ratio of 3:1, pre-cooled to −40 °C, containing an internal standard) was added to the tube, which was then vortexed for 30 s, homogenized at 40 Hz for 4 min, placed in an ice water bath and then subjected to ultrasound for 5 min. The steps of homogenization and ultrasound were repeated three times, followed by centrifugation, and dried by nitrogen blowing. The supernatant was filtered using a 0.22 μm microporous membrane and then analyzed by using an EXION LC System (SCIEX, Toronto, ON, Canada) ultra-high-performance liquid chromatography machine. The target compounds were separated using a Waters UPLC liquid chromatography column (Waters, Milford, MA, USA). The high-performance liquid chromatography A phase was a 0.1% formic acid aqueous solution; the B phase was acetonitrile. The column oven temperature was 40 °C, the temperature of the autosampler was 4 °C, and the injection volume was 2 μL. All mass spectrometry data acquisition and quantitative analysis of target compounds were performed using SCIEX Analyst Work Station Software (Version 1.6.3). After the mass spectrometry data were converted into mzXML format by ProteoWizard software (https://proteowizard.sourceforge.io/, accessed on 23 April 2023), the self-written R package (version 3.6.2, kernel is XCMS [26]) was used for peak identification, peak extraction, peak alignment, and integration, and then matched with BiotreeDB self-built secondary mass spectrometry database for material annotation [27,28].
2.6. Statistical Analysis
Eight treatments (sterilized and unsterilized spermosphere soil treatments at four stages of germination) were designed, and the microbiome and metabolome data of 48 samples were obtained. The data obtained from three replicates were expressed as mean value ± standard deviation. An independent sample t-test was used to compare the significant differences in germination indexes and seedling morphology indexes between the sterilized and unsterilized soil treatment groups. p value less than 0.05 was considered statistically significant. PE reads sequenced by Illumina was first spliced according to overlap, and sequence quality was controlled and filtered at the same time. Statistical analysis of community structure was conducted after samples were distinguished. Student’s t-test was used for species difference analysis, Spearman correlation coefficient statistical method was used for correlation heatmap analysis to determine the correlation between germination indexes and spermosphere bacterial community abundance, and between bacterial abundance and differential metabolite contents, and the images were drawn on the majorbio cloud platform (https://cloud.majorbio.com, accessed on 28 July 2022). Through metabolomics detection, 1104 features were identified, and the data were preprocessed, including the elimination of outliers identified by the interquartile range method, missing values in the metabolic raw data were replaced by numbers corresponding to half of the minimum value. The internal standard normalization method was used for data analysis, and three-dimensional data were obtained, including peak number, sample name, and normalized peak area. Orthogonal projections to latent structures-discriminate analysis (OPLS-DA) were carried out on the preprocessed data. Based on the OPLS-DA model and Student’s t test statistical method, the VIP value was set to be greater than 1, and the p-value was set to be less than 0.05, thus, the differential metabolites of different stages of germination were obtained. Advanced volcano plot (ggplot 2, R version 4.1.3), Lollipop plot, and KEGG enrichment scatter plot were performed using the OmicStudio tools (omicstudio.cn). The visualized network diagram was drawn according to the KEGG enrichment pathway in PPT software (https://www.genome.jp/kegg/pathway.html, accessed on 23 April 2023).
3. Results
3.1. Effects of Spermosphere Microorganisms on O. henryi Seed Germination and Seedling Growth
The unpaired t-test analysis (Table 1) was conducted on the indexes of sterilized soil and unsterilized soil, and the results showed that there were significant differences in germination rate, germination time, seedling height, root length, shoot dry weight, and root dry weight between sterilized and non-sterilized groups (p < 0.05). For diameter, there was no significant difference (p > 0.05). Seeds grown in unsterilized soil (T treatment) germinated 19 d earlier than seeds grown in sterilized soil (CK), and the germination rate (30.33% ± 1.53) is in the increased percentage that of the CK seed group (13.33% ± 0.8; Table 1). Furthermore, the shoot dry weight and root dry weight were higher than those obtained for the CK. The main difference between the sterilized soil and the unsterilized soil was the microbial community (for example, the species, abundance, and diversity), indicating that the differential microorganisms between the two treatment groups may be key to shortening seed germination time, enhancing germination rate and seedling growth.
3.2. Association Analysis of Dominant Bacteria
Bacterial sequences obtained from unsterilized soil and sterilized soil samples collected at the four seed germination stages were grouped into 6246 OTUs, which were assigned to 36 phyla, 101 classes, 241 orders, 422 families, and 846 genera. The correlation analysis of the top 150 spermospheres bacterial abundance at germination stage IV with germination rate and seedling growth indicators was conducted (Table S1), and 51 bacteria were screened to be significantly correlated with germination rate (Table 2). The results revealed that the dominant taxa such as Bacillus, Gemmatimonadaceae, Acidobacteriales, Nitrospira, Kitasatospora, Ellin6067, and Micromonosporaceae were significantly positively correlated (p ≤ 0.05, p < 0.01 or p < 0.001) with germination rate. The taxa of Bacillus, Nitrospira, Ellin6067, Kitasatospora, Gemmatimonadaceae, and Micromonosporaceae were significantly positively correlated with seedling height and root length, while the taxa of Citrifermentans, Microterricola, Variovorax, Beijerinckiaceae, Leifsonia, Labrys, Anaerosinus, Phenylobacterium, and Magnetospirillaceae were significantly negatively correlated with germination rate, seedling height and shoot dry weight. These data indicated that spermosphere bacteria may promote or inhibit seed germination and seedling growth.
Table 2.
The correlation between spermosphere bacteria at the genus level (top 150 most abundant taxa positively correlated with germination rate) and seed germination rate and seedling growth indexes in the cotyledon protrusion stage of germination.
3.3. Analysis O. henryi Seed Exudates
Samples of unsterilized and sterilized soil were analyzed by widely targeted multiple reaction monitoring (MRM) metabolomic (Figure 1A). In total, 1104 chemical compounds were identified in O. henryi seed exudates, including 198 alkaloids, 144 flavonoids, 133 terpenoids, 51 amino acids, and their derivatives, and 14 sugars. The principal component analysis score plot of the metabolites revealed that there were only small differences between biological replicates and that the three replicates of each treatment group clustered together (Figure 1B), indicating the repeatability and reliability of the experiment. Metabolites detected in sterilized soil formed separate clusters from those detected in unsterilized soil at the same germination stage, indicating that metabolites secreted by seeds were strongly influenced by the soil treatment methods. As germination progressed, the number of upregulated metabolites increased (Figure 1C–F), indicating that the metabolites secreted by seeds were greatly affected by the germination stage.
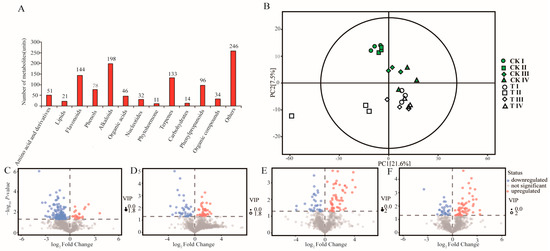
Figure 1.
Metabolic compounds detected in seed exudates. (A) Identified metabolites. (B) Principal component analysis (PCA) score plot of metabolites presents in unsterilized and sterilized (CK) soil at stages I to IV of seed germination (imbibition, radicle protrusion, radicle elongation, and cotyledon protrusion). (C–F) Volcano plots display the differential abundance of metabolites from seed exudates grown in unsterilized and sterilized soil at different germination stages. Each point in the volcanic map represents a metabolite. Red: metabolites enriched in unsterilized soil, blue: metabolites enriched in sterilized soil. The abscissa represents the multiple changes of unsterilized soil treatment compared with each substance (logarithm based on 2), the ordinate represents the p-value of the t-test (negative logarithm based on 10), and the scatter size represents the VIP value of OPLS-DA model. The larger the scatter, the greater the VIP value.
3.4. Different Stages of O. henryi Seed Germination
There were 140 significantly different microorganisms in stage I, 250 in stage II, 223 in stage III, and 266 in stage IV screened by the Student’s t-test method. Compared with the sterilization treatment (Figure 2), the taxa of Acidobacteriales, Ellin6067, Nitrospira, Micromonosporaceae, and Kitasatospora were significantly more abundant in the spermosphere microbial community throughout the germination process of seeds grown in unsterilized soil. The abundance of Bacillus in the unsterilized soil was significantly higher at germination stages II, III, and IV than in the sterilized soil, and the abundance of Acidobacteriales in the unsterilized soil was significantly higher at stages I, II, and III than in the sterilized soil, and the abundance of Nitrospira in the unsterilized soil was significantly higher at stages I, II, and IV than in the sterilized soil. These bacteria were not only dominant bacteria in the spermosphere microbial community (relative abundance ≥ 1%) but also were significantly positively correlated with the germination rate (Table 2). Furthermore, bacterial abundance in unsterilized soil was significantly higher than that in sterilized soil. By contrast, the abundance of Leifsonia and Labrys was significantly lower in unsterilized soil than in sterilized soil throughout the whole seed germination period. Likewise, the abundance of Magnetospirillaceae in the unsterilized soil was significantly lower at stages I, II, and IV than in the sterilized soil, and the abundance of Microterricola and Variovorax in the unsterilized soil were significantly lower at stages II, III, and IV than in the sterilized soil, and the abundance of Phenylobacterium in the unsterilized soil was significantly lower at stages III and IV than in the sterilized soil. These bacteria taxa were also significantly negatively correlated with germination rate (Table 2).
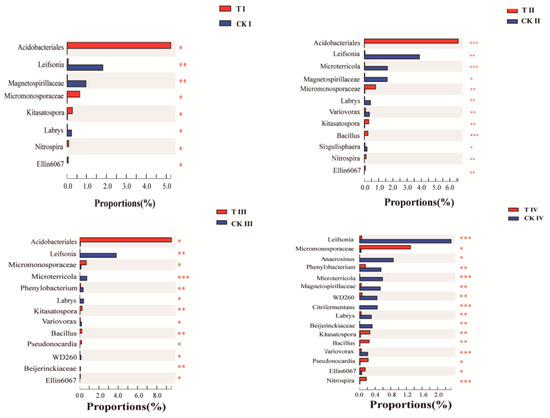
Figure 2.
Differential Bacterial taxa detected in unsterilized soil (T) and sterilized soil (CK) at each stage (imbibition (I), radicle protrusion (II), radicle elongation (III), and cotyledon protrusion (IV)) of O. henryi seed germination. *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
The metabolic profiles of exudates of seeds grown in unsterilized or sterilized soil were screened by the widely targeted multiple reaction monitoring(MRM) metabolomic analysis. Differential metabolites were analyzed using the following criteria: a p-value of ≤0.05 and a threshold of variable importance in projection (VIP) value of ≥1. Compared with the sterilized soil treatment group, the number of differential metabolites was 23 up-regulated and 132 down-regulated in the first stage, 42 up-regulated and 40 down-regulated in the second stage, 55 up-regulated and 32 down-regulated in the third stage, 58 up-regulated and 32 down-regulated in the fourth stage in the unsterilized soil treatment group, respectively. Over the course of germination from stages I to IV, the number of upregulated metabolites gradually increased (Figure 3, red). In stage I, more flavonoids and alkaloids were enriched, such as Abyssinone V, Pinocembrin, 7-Hydroxyflavone, Methyl 3-indolylacetate. In stages II, III, and IV, metabolites involved in providing seed germination energy and respiration were enriched, such as carbohydrates (alpha-D-Glucose, Maltotetraose), amino acids and derivatives (L-Lysine, 5-Hydroxylysine, L-Glutamine, L-Isoleucine), Organooxygen compounds (Beta-D-Fructose 2-phosphate, Betadex, Raffinose), Keto acids and derivatives (2-Keto-6-acetamidocaproate), and Nucleotide and its derivates (Thymidine, Uracil), etc.
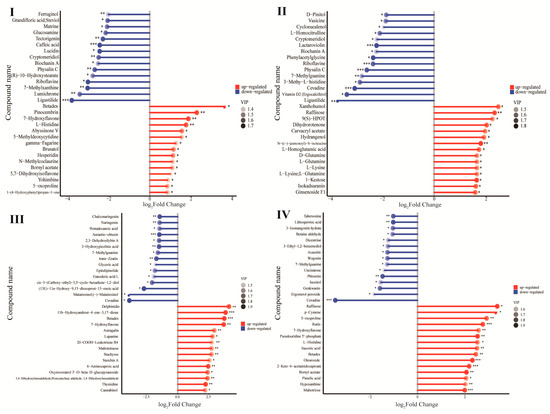
Figure 3.
Differential metabolites of O. henryi seeds exudates (seeds grown in unsterilized soil vs. sterilized soil) at different germination stages (imbibition (I), radicle protrusion (II), radicle elongation (III), and cotyledon protrusion (IV)). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001; VIP, variable importance in projection.
3.5. Correlation between Differential Metabolites and Differential Spermosphere Bacteria
The correlation heat map analysis was performed on the differential metabolites and differential spermosphere bacteria in each stage, and the first 70 differential metabolites were selected for image drawing. Heatmap analysis of the correlation between differential metabolites and the differential abundance of the bacterial genus in the spermosphere at different seed germination stages showed that at the initial stage of germination (Figure 4I), the taxa of Ellin6067 and Nitrospira were significantly positively correlated with most upregulated metabolites, which mainly comprised fatty acids, flavonoids and other metabolites, such as 8,9-DiHETrE and itaconic acid. The number of carbohydrate metabolites increased at stage II of germination (Figure 4II). the abundance of the Acidobacteriales taxon showed significant positive correlations with the contents of glucose, fructotriose, raffinose, kojibiose, and other carbohydrates. The taxa of Bacillus and Micromonosporaceae showed significant positive correlations with the contents of L-homoglutamic acid, lysine, and glutamine, as well as amino acids and carbohydrate metabolites such as kojibiose and raffinose. The abundance of Nitrospira was significantly positively correlated with the contents of 2-keto-6-acetamidocaproate, α-D-glucose. The abundance of Microterricola, Variovorax, Leifsonia showed significant positive correlations with the contents of metabolites, such as phenylalanine, benzene and substituted derivatives, and alkaloids. At stage III of germination (Figure 4III), the abundance of Bacillus, Kitasatospora, and Micromonosporaceae taxa was significantly positively correlated with the contents of glucose, β-D-fructose 2-phosphate, pyrimidine, thymidine, and other metabolites. The abundance of Bacillus and Ellin6067 was significantly positively correlated with the contents of maltose, stachyose, and other carbohydrates. The abundance of acidobacteriales taxa was significantly positively correlated with plant hormones and keto-acid metabolites such as trans-zeatin riboside, indole-3-carboxaldehyde, and 2-keto-6-acetamidocaproate. The abundance of spermosphere bacteria that were lower in the unsterilized soil than in sterilized soil, such as Beijerinckiaceae, Variovorax, and Leifsonia, were significantly positively correlated with most phenols and alkaloids. At stage IV (Figure 4IV), the abundance of Bacillus was significantly positively correlated with the contents of sugars, fatty acyls, flavonoids, and other metabolites, such as itaconic acid, pimelic acid, rutin, and 1-kestose. The abundance of Ellin6067 was significantly positively correlated with most alkaloids, terpenoids, and other metabolites, while the effect of Nitrospira was not significant at this stage.
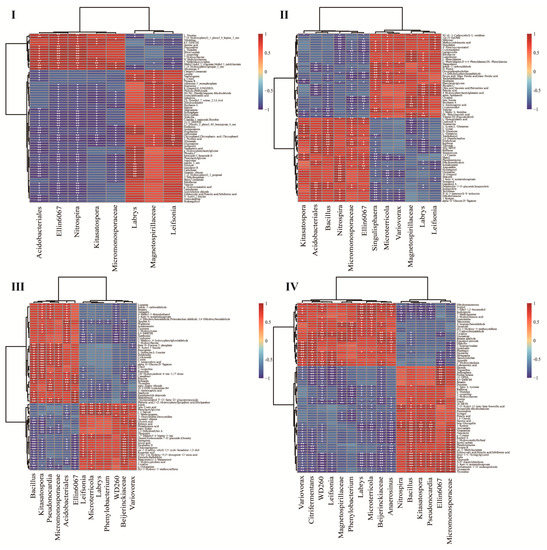
Figure 4.
Correlation heatmap of differential bacteria in the spermosphere and differential metabolites in seed exudates detected at each stage of seed germination (imbibition (I), radicle protrusion (II), radicle elongation (III), and cotyledon protrusion (IV)) of O. henryi seeds grown in unsterilized soil vs. sterilized soil. *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01.
These analyses indicated that at the initial stage of germination (I), the seed coat might soften due to the actions of Ellin6067 and Nitrospira, which starts the O. henryi seed germination process. At the middle and late stages of germination (II, III, IV), seeds consume metabolites such as sugars and amino acids as a source of energy due to the interactions of the taxa of Acidobacteriales, Bacillus, Micromonosporaceae, and Nitrospira. At the radicle protrusion stage in the unsterilized soil group, levels of metabolites such as sugars and amino acids increased in seeds and were significantly positively correlated with the more dominant bacterial taxa, indicating that these metabolites might act as a nutrient carbon source for these microorganisms, thus attracting spermosphere bacteria to gather around the seed, thereby activating more respiratory metabolic pathways, and enhancing seed respiratory metabolism, so this stage may therefore be a critical period for initiating seed germination. At the radicle protrusion stage in the unsterilized soil group, hormone metabolites in seeds, particularly cytokinins, were significantly upregulated, indicating that the speed of embryo division and differentiation was accelerated at this stage.
3.6. KEGG Enrichment Pathway Analysis
The up-regulated differential metabolites in the different germination stages of the unsterilized treatment group were analyzed by the KEGG enrichment pathway. The result showed that at stage I, the seed coat began to rupture, and a large number of fatty acyl metabolites flowed out (Figure 5). The taxa of Ellin6067, Nitrospira, and Acidobacteriales activated significant enrichment of C5 branched-chain dibasic acid metabolism and β-alanine metabolism. At the middle stages of germination (stages II and III), the taxa of Acidobacteriales, Bacillus, and Nitrospira mainly activated and significantly enriched metabolic pathways such as galactose metabolism, lysine degradation, amino acid biosynthesis, and metabolism, plant hormone signal transduction, fructose, and mannose metabolism, and the degradation of the branched-chain amino acids (BCAAs) valine, leucine, and isoleucine, suggesting that during this stage, sugar metabolism, amino acid metabolism, and hormone transduction provide energy and signal transmission for seed germination, and that seed germination respiration was enhanced, which was considered to be the critical period of germination. At stage IV, the taxa of Bacillus and Ellin6067 might mainly activate the β-alanine metabolism, pyrimidine metabolism, and arachidonic acid metabolism pathways, and were significantly enriched (p < 0.05). In summary, our analyses suggested that the spermosphere bacteria in unsterilized soil (e.g., the taxa of Bacillus, Acidobacteriales, Ellin6067, and Nitrospira) played an important role in promoting seed germination.
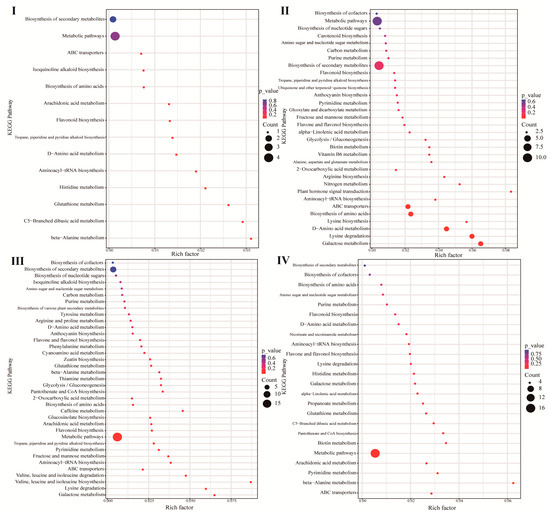
Figure 5.
KEGG enrichment pathway analysis at stages (I–IV) (imbibition, radicle protrusion, radicle elongation, and cotyledon protrusion) of germination.
3.7. Visual Network Diagram
A visual network diagram was constructed (Figure 6) according to the enrichment metabolic pathways of the upregulated metabolites in the unsterilized soil. With regard to metabolites related to the upstream pathways of glycolysis, the pentose phosphate pathway, and the citrate cycle, such as raffinose, α-D-glucose, and D-tagatose, the relative contents of these metabolites in the spermosphere of unsterilized soil were higher than those in sterilized soil at each germination stage, in addition, the contents of 5-hydroxy-L-lysine, 2-Keto-6-acetamidohexanoate, 5-Aminopentanoate, L-2-Aminoadipate in the unsterilized soil at the middle stages (radicle protrusion and radicle elongation) of germination were higher than in the sterilized soil. The results indicated that the interaction of the taxa of Bacillus, Acidobacteriales, Ellin6067, and Nitrospira activated the respiratory metabolism in seeds to accelerate seed germination and activated the degradation of lysine metabolic pathway for seed germination energy utilization.
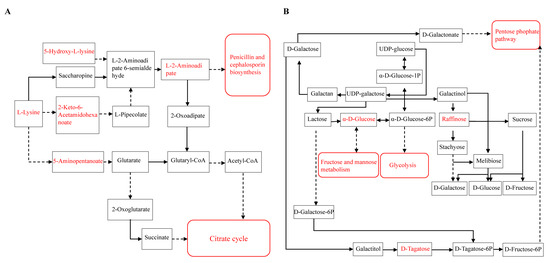
Figure 6.
Visualized network diagram of lysine degradation (A) and galactose metabolism (B). Red represents upregulated metabolites and metabolic pathways in the unsterilized soil treatment.
4. Discussion
4.1. The Role of Spermosphere Bacterial Functions in Seed Germination
Seed germination is a critical stage in the life cycle of a plant. By definition, germination begins with seed imbibition and ends with the emergence of the embryonic axis (usually radicles), during which a series of ordered physiological and morphogenetic processes occur [29]. Seed germination is inherently related to metabolism. Metabolites in seeds change during maturation, drying, and germination. A seed’s metabolite content and germination ability depend on its potential genetic structure and the impact of environmental factors during development [30]. In this study, the germination rate, germination time, and seedling growth index of seeds grown in unsterilized soil were better than those of seeds grown in sterilized soil. The biggest difference between the sterilized soil and unsterilized soil was the microorganisms, indicating that microorganisms may be a key factor affecting the germination time, germination rate, and seedling growth of O. henryi seeds. Among the bacteria detected in the spermosphere, the taxa of Bacillus, Acidobacteriales, Nitrospira, Kitasatospora, Ellin6067, and Micromonosporaceae showed a significant positive correlation (p ≤ 0.05, p < 0.01, or p < 0.001) with germination rate. Bacillus sp. is a common Gram-positive thick-walled bacterium. Many species of Bacillus are involved in plant growth promotion functions. Among their core genes, genes responsible for mineral phosphate dissolution, siderophores, extracellular polysaccharides, flagella biosynthesis, biofilm formation, and IAA production have been reported [31]. Bacillus subtilis can increase the concentration of active oxygen in seeds through specific components of its own extracellular matrix, amyloid protein TasA, and fengycin, and induce specific changes in lipid metabolism and glutathione-related molecular accumulation by targeting oil bodies in the seed endosperm to stimulate radicle development and promote seed germination [32]. Furthermore, Bacillus. subtilis can promote germination by inducing α-amylase activity [33]. Some members of Acidobacteria are capable of degrading complex biopolymers, including those in plant litter, such as xylans, pectin, and chitin [34], and play an important role in maintaining soil ecosystem stability. Ellin6067, which is a member of the Nitrosomonadaceae, is the most representative in terms of degrading heterologous and other complex organic compounds, which can reduce the toxicity of heavy metals [35]. Nitrospira can perform complete nitrification and, therefore, play a key role in nitrogen-cycling microbial communities [36]. Furthermore, studies have revealed that complete ammonia oxidizers, comammox Nitrospira, are dominant in soil nitrification [37] and have a cellulose-degrading function [38]. Therefore, many of the functions of these spermosphere bacteria could play an important role in promoting seed germination.
4.2. Metabolic Pathways Activated by Spermosphere Bacteria Involved in Seed Germination
Biopriming is a new seed treatment technology that combines the biological treatment of seed with a beneficial microorganism with controlled water absorption before sowing to not only improve the seed germination rate and seedling uniformity but also protect seeds from soil and seed-borne diseases [39]. Biopriming is one type of seed priming. No matter what kind of seed priming technology is used, the regulation of seed germination metabolism is directly activated by controlled seed hydration. In the early stages of seed hydration, oxygen is absorbed, and carbon dioxide is released, indicating that respiration is activated to provide energy for subsequent seed germination [40,41]. In addition, seeds can also provide energy for germination through anaerobic respiration or other pathways. When pyruvate in seeds is blocked, anaerobic respiration is activated [42]. In conclusion, the activation of respiratory metabolism is key for successful germination [43].
Our analyses suggest that the taxa of Acidobacteriales, Ellin6067, and Nitrospira in unsterilized soil at stage I of germination mainly activate C5 branched-chain dibasic acid metabolism and the β-alanine metabolism pathway and are significantly more abundant than in sterilized soil. Nitrification performed by Ellin6067 and Nitrospira can lead to soil acidification [44]. The taxa of Acidobacteriales are acidophilic bacteria, whose presence indicates soil acidification. In addition, both the taxa of Acidobacteriales and Nitrospira can degrade cellulose or act as symbionts involved in the metabolism of refractory nutrients [34,38]. The seed coat structure of O. henryi comprises palisade tissue and stone cells. Excessive cellulose and lignin structures lead to hard seeds and impervious seed coats. At the stage of imbibition, few metabolic regulatory pathways are activated inside seeds, so the initiation of O. henryi seed germination may be related to bacteria in the seed coat. Under suitable environmental conditions, nitrification of the soil by the taxa of Nitrospira and Acidobacteriales may create an acid environment in the spermosphere, causing the seed coat to soften, which initiates germination. Although β-alanine is not involved in the synthesis of proteins or enzymes, it is important for the later synthesis of coenzyme A (CoA) because it is an important metabolic hub for the entry of three major nutrients into the citrate cycle (TCA) [45]. The taxa of Acidobacteriales, Bacillus, Micromonosporaceae, Nitrospira, and Ellin6067 activate and significantly enrich galactose metabolism, fructose and mannose metabolism, lysine degradation metabolism, and BCAA metabolism through interactions at stages of radicle protrusion and elongation. The enrichment of more carbohydrate metabolic pathways indicates that the increase in the contents of sugar and amino acid at the early stage of germination plays a vital role in promoting the germination of O. henryi seeds, which is consistent with data obtained by Zhao et al. and Yan et al. [46,47]. Lysine can directly enter the TCA cycle through catabolism, and its degradation helps to maintain mitochondrial respiration under carbon-limited conditions during aging [48]. This is similar to the effect achieved when using hydro-electro hybrid priming to promote carrot seed germination [43]. During germination under abiotic stress, sugar metabolism in seeds may be activated to respond to the abiotic stress and ensure seed germination, plant growth, and development, therefore, sugar metabolism plays an active role in the ecological adaptation of plants. At the stage of cotyledon protrusion, Bacillus and other spermosphere bacteria activate the significant enrichment of β-alanine metabolism, pyrimidine metabolism, and arachidonic acid metabolism. β-alanine is important for the synthesis of pantothenic acid and subsequently CoA. CoA is an essential cofactor for many metabolic and energy reactions and is a key component of life [45], providing a new energy supply for seedling development and growth after germination.
4.3. Effect of Spermosphere Bacteria Abundance on Seed Germination and Plant Growth
Regarding the related research on the effect of bacterial abundance on seed germination rate and seedling emergence, we found that most members of Acidobacteria can decompose soil organic matter, which is important in the carbon cycle and promotes seed germination and plant performance [49]. Studies have found that the abundance of Acidobacteria and Nitrospirae play an important role in promoting seed germination and the pot planting experiments of sterilized soil and unsterilized soil were carried out. The relative abundance of Acidobacteria and Nitrospirae in unsterilized soil was significantly higher than that in sterilized soil, and the seed germination rate of unsterilized soil was higher than that of sterilized soil, which indicated that the high abundance of Acidobacteria and Nitrospirae promoted seed germination. Climate warming can affect seed germination and seedling growth by changing plant-soil microbial interactions, and Acidobacteria members play a key role in plant-soil microbial interactions, demonstrating that the abundance of Acidobacteria and Nitrospirae can improve the adverse effects of climate warming on seed germination and plant performance [50]. As the main member of plant PGPR (plant growth-promoting rhizobacteria), Bacillus is famous for its nitrogen fixation ability. It has the functions of solubilizing phosphorus, reducing iron and sulfur, and showing good growth promotion and stress resistance for many plants [51,52]. Therefore, the increase in the abundance of Bacillus in plants will have a positive impact on nutrient cycling and improving plant performance.
In summary, the spermosphere bacteria of O. henryi seeds play a vital role in seed germination. Under appropriate climatic conditions, some spermosphere bacteria can activate more energy metabolism pathways in the seed to maintain seed development through their functional advantages. This study outlines the roles played by spermosphere bacteria in the germination of O. henryi seeds, providing a theoretical basis for improving the interaction between spermosphere microorganisms and seed germination. The next step will be to screen, isolate and purify spermosphere functional bacteria for application in the artificial seed breeding practice of O. henryi, and explore the ability of these bacteria to colonize and chemotaxis to key differential metabolites of seed exudates, thereby permanently recruiting them to play a beneficial role on the seed coat.
5. Conclusions
The abundance and functional characteristics of some spermosphere bacteria play important roles in promoting the germination of the hard seeds of O. henryi. The key spermosphere microbial groups are the taxa of Bacillus, Acidobacteriales, Ellin6067, and Nitrospira. The critical period when spermosphere bacteria promote seed germination is at the radicle protrusion stage, during which stage energy metabolism and hormone signal transduction are enhanced, and the seed respiration metabolism is accelerated. Spermosphere functional bacteria such as taxa of Acidobacteriales, Bacillus, Ellin6067, and Nitrospira may break through the hard seed coat and germinate through conditions such as their own functions and soil acid environment (suitable climatic conditions), and then activate the plant hormone signal transduction metabolic pathways for signal transmission, as well as activating galactose metabolism, lysine degradation, Valine, leucine and isoleucine degradation and other significant enrichment metabolic pathways in the seed to provide energy for germination.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14061136/s1, Table S1: The correlation analysis between spermosphere bacterial at the genus level (top 150 most abundant taxa) and seed germination rate and seedling growth indexes in the cotyledon protrusion stage of germination.
Author Contributions
M.G. and X.W. conceived this study; M.G. carried out the experiment, analyzed the results, and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (grant number: 32260384); Guizhou Province High-level Innovative Talents Training Plan Project (grant number: (2016)5661) and Guizhou Graduate Research Fund (grant number: YJSCXJH(2020)064).
Data Availability Statement
The Link to the original data of the spermosphere bacteria in this manuscript has been created and is available at: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA935683, accessed on 16 February 2023.
Acknowledgments
We thank Mei Fan and Xueqing Tian for assistance with conducting the experiments. All individuals have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Nelson, E.B. Microbial dynamics and interactions in the spermosphere. Annu. Rev. Phytopathol. 2004, 42, 271–309. [Google Scholar] [CrossRef]
- Lopez-Velasco, G.; Carder, P.A.; Welbaum, G.E.; Ponder, M.A. Diversity of the spinach (Spinacia oleracea) spermosphere and phyllosphere bacterial communities. FEMS Microbiol. Lett. 2013, 346, 146–154. [Google Scholar] [CrossRef]
- Schiltz, S.; Gaillard, I.; Pawlicki-Jullian, N.; Thiombiano, B.; Mesnard, F.; Gontier, E.A. Review: What is the spermosphere and how can it be studied? J. Appl. Microbiol. 2015, 119, 1467–1481. [Google Scholar] [CrossRef]
- Pitzschke, A. Developmental peculiarities and seed-borne endophytes in quinoa: Omnipresent, robust Bacilli contribute to plant fitness. Front. Microbiol. 2016, 7, 2. [Google Scholar] [CrossRef]
- Nelson, E.B. The seed microbiome: Origins, interactions, and impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Pal, G.; Kumar, K.; Verma, A.; Verma, S.K. Seed inhabiting bacterial endophytes of maize promote seedling establishment and provide protection against fungal disease. Microbiol. Res. 2021, 255, 126926. [Google Scholar] [CrossRef]
- Shalini, D.; Benson, A.; Gomathi, R.; Henry, A.J.; Jerritta, S.; Joe, M.M. Isolation, characterization of glycolipid type biosurfactant from endophytic Acinetobacter sp. ACMS25 and evaluation of its biocontrol efficiency against Xanthomonas oryzae. Biocatal. Agric. Biotechnol. 2017, 11, 252–258. [Google Scholar] [CrossRef]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.L.; Ma, B.; et al. Bacterial seed endophyte shapes disease resistance in rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef]
- Chanratana, M.; Han, G.H.; Joe, M.M.; Choudhury, A.R.; Sundaram, S.; Halim, M.A.; Sa, T. Evaluation of chitosan and alginate immobilized Methylobacterium oryzae CBMB20 on tomato plant growth. Arch. Agron. Soil Sci. 2018, 64, 1489–1502. [Google Scholar] [CrossRef]
- Chahtane, H.; Nogueira Füller, T.; Allard, P.-M.; Marcourt, L.; Queiroz, E.F.; Shanmugabalaji, V.; Falquet, J.; Wolfender, J.-L.; Lopez-Molina, L. The plant pathogen Pseudomonas aeruginosa triggers a DELLA-dependent seed germination arrest in Arabidopsis. eLife 2018, 7, e37082. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, D.; Dai, L.X.; Ding, H.; Ci, D.W.; Qin, F.F.; Zhang, G.C.; Zhang, Z.M. Influence of salt stress on growth of spermosphere bacterial communities in different peanut (Arachis hypogaea L.) cultivars. Int. J. Mol. Sci. 2020, 21, 2131. [Google Scholar] [CrossRef]
- Dai, Y.; Li, X.Y.; Wang, Y.; Li, C.X.; He, Y.; Lin, H.H.; Wang, T.; Ma, X.R. The differences and overlaps in the seed-resident microbiome of four Leguminous and three Gramineous forages. Microb. Biotechnol. 2020, 13, 1461–1476. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.N.; Osman, M.E.; Kasim, W.A.; Salama, S. Characterization of two drought-tolerant PGPB. Azospirillum brasilense NO40 and Stenotrophomonas maltophilia. J. Environ. Sci. 2017, 46, 121–136. [Google Scholar]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Asaf, S.; Khan, M.A.; Kang, S.M.; Yun, B.W.; Lee, I.J. Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol. Biochem. 2016, 106, 236–243. [Google Scholar] [CrossRef]
- Sorty, A.M.; Meena, K.K.; Choudhary, K.; Bitla, U.M.; Minhas, P.S.; Krishnani, K.K. Effect of plant growth promoting bacteria associated with halophytic weed (Psoralea corylifolia L) on germination and seedling growth of wheat under saline conditions. Appl. Biochem. Biotechnol. 2016, 180, 872–882. [Google Scholar] [CrossRef]
- Johnsson, L.; Hökeberg, M.; Gerhardson, B. Performance of the Pseudomonas chlororaphis biocontrol agent MA 342 against cereal seed borne diseases in field experiments. Eur. J. Plant Pathol. 1998, 104, 701–711. [Google Scholar] [CrossRef]
- Lovegrove, A.; Hooley, R. Gibberellin and abscisic acid signalling in aleurone. Trends Plant Sci. 2000, 5, 102–110. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Y.; Zhang, Y.C.; Lin, C.; Gong, D.T.; Guan, Y.J.; Hu, J. Reactive oxygen species and gibberellin acid mutual induction to regulate tobacco seed germination. Front. Plant Sci. 2018, 9, 1279. [Google Scholar] [CrossRef]
- Iannucci, A.; Canfora, L.; Nigro, F.; Vita, P.D.; Beleggia, R. Relationships between root morphology, root exudate compounds and rhizosphere microbial community in durum wheat. Appl. Soil Ecol. 2021, 158, 103781. [Google Scholar] [CrossRef]
- Wei, X.L.; Meng, X.S.; Deng, Z. Relation between being endangered and seed reproductive ecology of a rare species Ormosia henryi. Seed 2014, 33, 82–86. [Google Scholar]
- Wang, T.T.; Wei, X.L. Isolation, screening and identification of spermosphere fungi from Ormosia henryi. J. Northwest For. Univ. 2021, 36, 167–172. (In Chinese) [Google Scholar]
- Hu, W.T.; Wei, S.Z.; Chen, H.; Tang, M. Effect of Sterilization on Arbuscular Mycorrhizal Fungal Activity and Soil Nutrient Status. J. Soil Sci. Plant Nutr. 2019, 20, 684–689. [Google Scholar] [CrossRef]
- Windstam, S.; Nelson, E.B. Temporal release of fatty acids and sugars in the spermosphere: Impacts on Enterobacter cloacae-induced biological control. Appl. Environ. Microbiol. 2008, 74, 4292–4299. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.D. Soil Agricultural Chemistry Analysis, 3rd ed.; Agriculture Press: Beijing, China, 2000; pp. 22–110. (In Chinese) [Google Scholar]
- Bai, J.; Xu, D.M.; Xie, D.M.; Wang, M.; Li, Z.Q.; Guo, X.S. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfafa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, C.; Tautenhahn, R.; Böttcher, C.; Larson, T.; Neumann, S. CAMERA: An Integrated Strategy for Compound Spectra Extraction and Annotation of Liquid Chromatography/Mass Spectrometry Data Sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Tong, X.; Peng, Y.; Ma, P.; Zhang, M.J.; Lu, H.M.; Chen, X.Q.; Liang, Y.Z. Multiscale peak detection in wavelet space. Analyst 2015, 140, 7955–7964. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Kazmi, R.H.; Willems, L.A.J.; Joosen, R.V.L.; Khan, N.; Ligterink, W.; Hilhorst, H.W.M. Metabolomic analysis of tomato seed germination. Metabolomics 2017, 13, 145. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Bakshi, U.; Mallick, I.; Mukherji, S.; Bera, B.; Ghosh, A. Genome-guided insights into the plant growth promotion capabilities of the physiologically versatile Bacillus aryabhattai strain AB211. Front. Microbiol. 2017, 8, 411. [Google Scholar] [CrossRef]
- Berlanga-Clavero, M.V.; Molina-Santiago, C.; Caraballo-Rodríguez, A.M.; Petras, D.; Díaz-Martínez, L.; Pérez-García, A.; de Vicente, A.; Carrión, V.J.; Dorrestein, P.C.; Romero, D. Bacillus subtilis biofilm matrix components target seed oil bodies to promote growth and anti-fungal resistance in melon. Nat. Microbiol. 2022, 7, 1001–1015. [Google Scholar] [CrossRef]
- Li, Y.J.; Hu, Q.P. Studying of the promotion mechanism of Bacillus subtilis QM3 on wheat seed germination based on β-amylase. Open Life Sci. 2020, 15, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Eichorst, S.A.; Kuske, C.R.; Schmidt, T.M. Influence of plant polymers on the distribution and cultivation of bacteria in the phylum Acidobacteria. Appl. Environ. Microbiol. 2011, 77, 586–596. [Google Scholar] [CrossRef]
- Liu, H.W.; Yuan, R.L.; Sarkodie, E.K.; Tang, J.H.; Jiang, L.H.; Miao, B.; Liu, X.D.; Zhang, S.Y. Insight into functional microorganisms in wet–dry conversion to alleviate the toxicity of chromium fractions in red soil. Front. Microbiol. 2022, 13, 977171. [Google Scholar] [CrossRef]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.J.; Zhao, Y.X.; Yao, X.W.; Wang, J.Q.; Zheng, P.; Xi, C.W.; Hu, B.L. Dominance of comammox Nitrospira in soil nitrification. Sci. Total Environ. 2021, 780, 146558. [Google Scholar] [CrossRef]
- Chen, C.L.; Gan, Z.H.; Xu, R.H.; Meng, F.G. Cellulose-induced shifts in microbial communities and microbial interactions in an anoxic/aerobic membrane bioreactor. J. Water Process. Eng. 2021, 42, 102106. [Google Scholar] [CrossRef]
- Forti, C.; Shankar, A.; Singh, A.; Balestrazzi, A.; Prasad, V.; Macovei, A. Hydropriming and biopriming improve Medicago truncatula seed germination and upregulate DNA repair and antioxidant genes. Genes 2020, 11, 242. [Google Scholar] [CrossRef]
- Bello, P.; Bradford, K.J. Single-seed oxygen consumption measurements and population-based threshold models link respiration and germination rates under diverse conditions. Seed Sci. Res. 2016, 26, 199–221. [Google Scholar] [CrossRef]
- Płażek, A.; Dubert, F.; Kopeć, P.; Dziurka, M.; Kalandyk, A.; Pastuszak, J.; Wolko, B. Seed hydropriming and smoke water significantly improve low-temperature germination of Lupinus angustifolius L. Int. J. Mol. Sci. 2018, 19, 992. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef]
- Zhao, S.; Garcia, D.; Zhao, Y.L.; Huang, D.F. Hydro-electro hybrid priming promotes carrot (Daucus carota L.) seed germination by activating lipid utilization and respiratory metabolism. Int. J. Mol. Sci. 2021, 22, 11090. [Google Scholar] [CrossRef]
- Cáceres, R.; Malińska, K.; Marfà, O. Nitrification within composting: A review. Waste Manag. 2018, 72, 119–137. [Google Scholar] [CrossRef]
- Perchat, N.; Dubois, C.; Mor-Gautier, R.; Duquesne, S.; Lechaplais, C.; Roche, D.; Fouteau, S.; Darii, E.; Perret, A. Characterization of a novel β-alanine biosynthetic pathway consisting of promiscuous metabolic enzymes. J. Biol. Chem. 2022, 298, 102067. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.F.; Liu, Y.; Huang, Y.Q.; Liang, Q.Y.; Cai, S.G.; Zhang, G.P. Time-course comparative metabolome analysis of different barley varieties during malting. J. Agric. Food Chem. 2022, 70, 2051–2059. [Google Scholar] [CrossRef]
- Yan, H.F.; Nie, Y.T.; Cui, K.L.; Sun, J. Integrative transcriptome and metabolome profiles reveal common and unique pathways involved in seed initial imbibition under artificial and natural salt stresses during germination of halophyte quinoa. Front. Plant Sci. 2022, 13, 853326. [Google Scholar] [CrossRef]
- Kamranfar, I.; Xue, G.P.; Tohge, T.; Sedaghatmehr, M.; Fernie, A.R.; Balazadeh, S.; Mueller-Roeber, B. Transcription factor RD 26 is a key regulator of metabolic reprogramming during dark-induced senescence. New Phytol. 2018, 218, 1543–1557. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Wang, J.J.; Bonser, S.P.; Liu, K.; Liu, Z.Y.; Gao, H.N.; Cui, H.W.; Chen, J.W.; Wang, Y.J.; Song, H.X.; Meng, L.H.; et al. Warming affects herbaceous germination, early survival, and growth by shifting plant-soil microbe interactions in an alpine ecosystem. Plant Soil 2023, 1–17. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Mahmoud, A.B.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.K.; Li, Y.Y.; Yao, H.Y. Biochar Amendment Stimulates Utilization of Plant-Derived Carbon by Soil Bacteria in an Intercropping System. Front. Microbiol. 2019, 10, 1361. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).