
Ecological Factors Driving Tree Diversity across Spatial Scales in Temperate Forests, Northeast China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region and Sample Sites
2.2. Tree Diversity Scaling
2.3. Explanatory Variable Selection
2.4. Data Analysis
3. Results
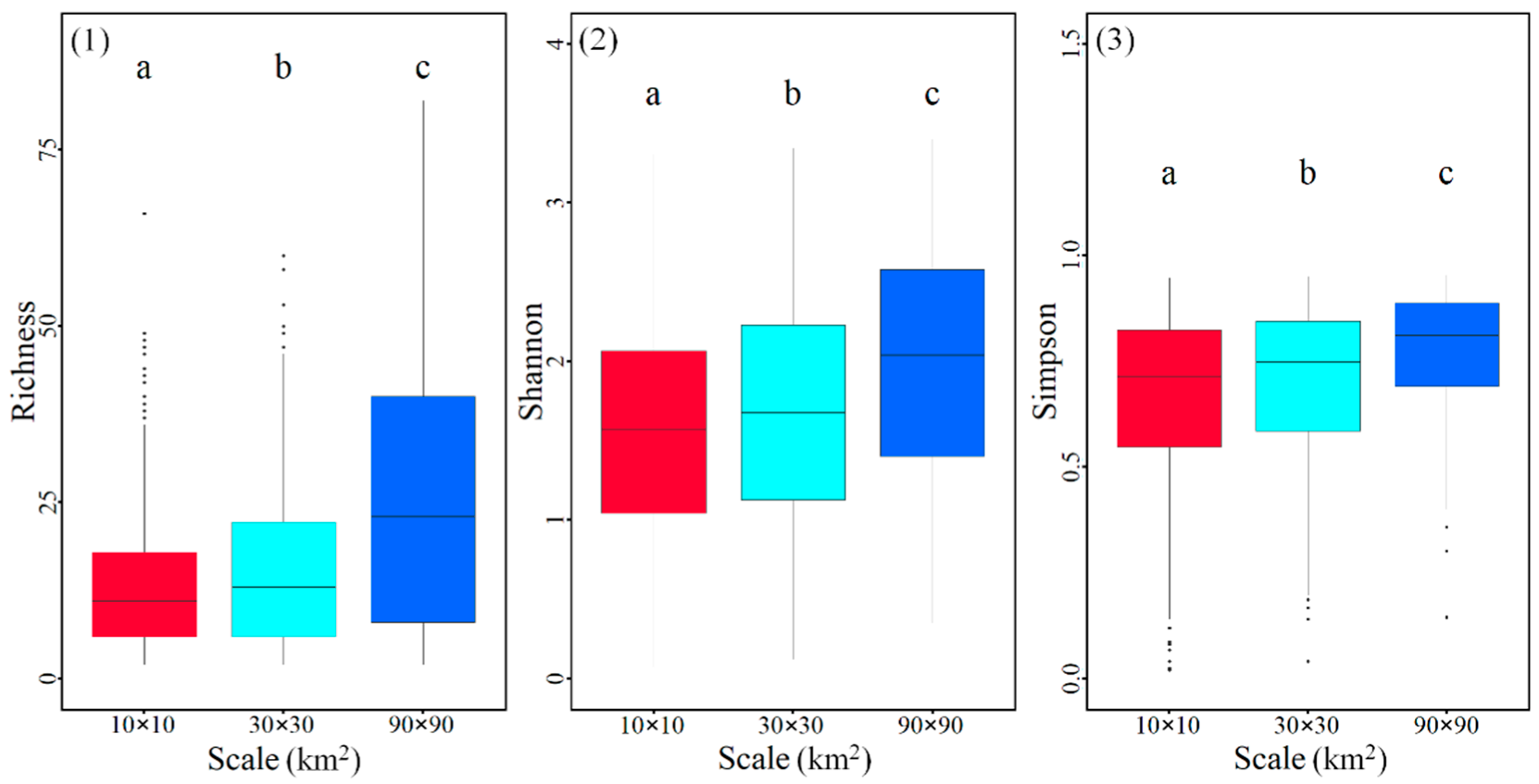
3.1. Tree Diversity at Different Spatial Scales
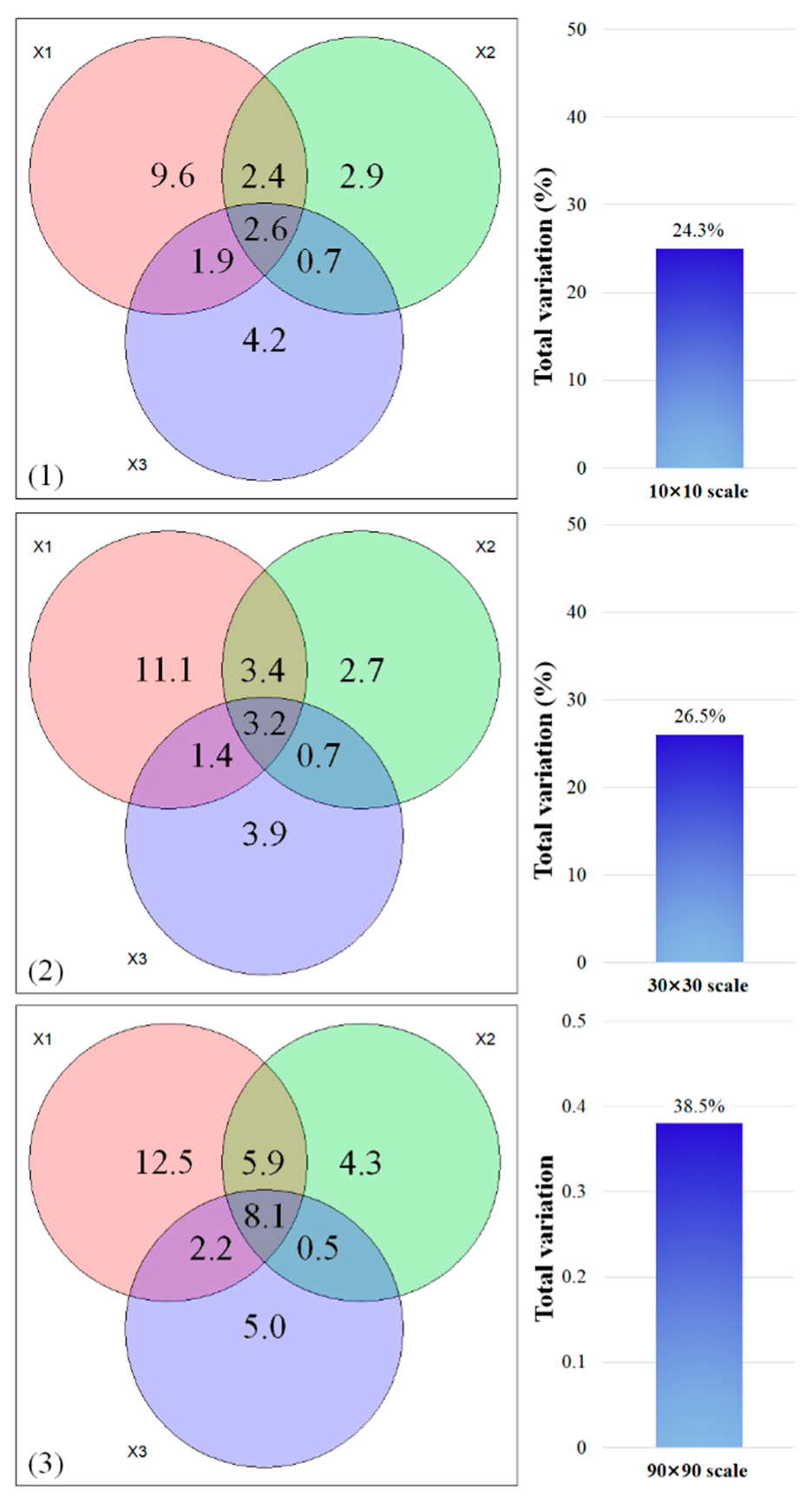
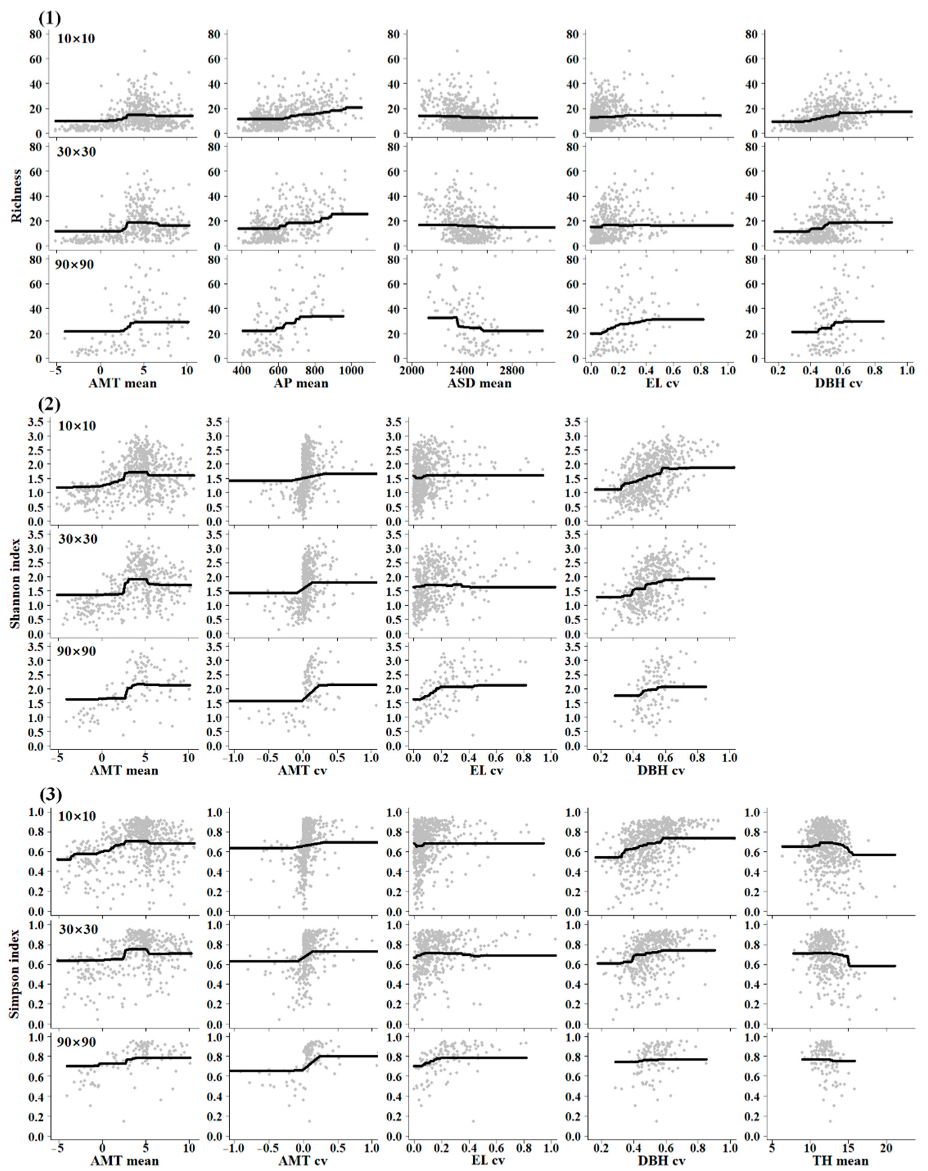
3.2. Ecological Drivers of Tree Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawley, M.J.; Harral, J.E. Scale dependence in plant biodiversity. Science 2001, 291, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Belmaker, J.; Jetz, W. Cross-scale variation in species richness-environment associations. Global Ecol. Biogeogr. 2011, 20, 464–474. [Google Scholar] [CrossRef]
- Malanson, G.P.; Fagre, D.B.; Zimmerman, D.L. Scale dependence of diversity in alpine tundra, Rocky Mountains, USA. Plant Ecol. 2018, 219, 999–1008. [Google Scholar] [CrossRef]
- Keil, P.; Chase, J.M. Global patterns and drivers of tree diversity integrated across a continuum of spatial grains. Nat. Ecol. Evol. 2019, 3, 390–399. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- McGill, B.J. Matters of Scale. Science 2010, 328, 575–576. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224. [Google Scholar] [CrossRef]
- Chase, J.M.; Knight, T.M. Scale-dependent effect sizes of ecological drivers on biodiversity: Why standardised sampling is not enough. Ecol. Lett. 2013, 16, 17–26. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Brooker, R.W.; Butterfield, B.J.; Cook, B.J.; Kikvidze, Z.; Lortie, C.J.; Callaway, R.M. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 2014, 17, 193–202. [Google Scholar] [CrossRef]
- Gavilan, R.G.; Callaway, R.M. Effects of foundation species above and below tree line. Plant Biosyst. 2017, 151, 665–672. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Vormisto, J.; Duque, A.; Sánchez, M.; Paredes, V.V.; Lähteenoja, O. Effect of sampling grain on patterns of species richness and turnover in Amazonian forests. Ecography 2017, 40, 840–852. [Google Scholar] [CrossRef]
- Hillebrand, H.; Blenckner, T. Regional and local impact on species diversity—From pattern to processes. Oecologia 2002, 132, 479–491. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fang, J.; Sanders, N.J.; White, P.S.; Tang, Z. Relative importance of climate vs local factors in shaping the regional patterns of forest plant richness across northeast China. Ecography 2009, 32, 133–142. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Wang, R.; Wang, Y.; Liu, J. Relationships between plant species richness and environmental factors in nature reserves at different spatial scales. Pol. J. Environ. Stud. 2017, 26, 2375–2384. [Google Scholar] [CrossRef]
- Harrison, S.; Spasojevic, M.J.; Li, D. Climate and plant community diversity in space and time. Proc. Natl. Acad. Sci. USA 2020, 117, 4464–4470. [Google Scholar] [CrossRef]
- Naud, L.; Msviken, J.; Freire, S.; Angerbjorn, A.; Dalen, L.; Dalerum, L. Altitude effects on spatial components of vascular plant diversity in a subarctic mountain tundra. Ecol. Evol. 2019, 9, 4783–4795. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [Green Version]
- Sibly, R.M.; Brown, J.H.; Kodric-Brown, A. Metabolic Ecology: A Scaling Approach; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar] [CrossRef]
- Gaston, K.J.; Chown, S.L. Why Rapoport’s Rule Does Not Generalise. Oikos 1999, 84, 309. [Google Scholar] [CrossRef]
- O’Brien, E.; Field, R.; Whittaker, R. Climatic gradients in woody plant (tree and shrub) diversity: Water-energy dynamics, residual variation, and topography. Oikos 2000, 89, 588–600. [Google Scholar] [CrossRef]
- Wang, Z.; Fang, J.; Lin, T.X. Patterns, determinants and models of woody plant diversity in China. Proc. Biol. Sci. 2011, 278, 2122–2132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A. Multiple drivers of plant diversity in forest ecosystems. Global Ecol. Biogeogr. 2014, 23, 885–893. [Google Scholar] [CrossRef]
- Cowles, J.; Boldgiv, B.; Liancourt, P.; Petraitis, P.S.; Casper, B.B. Effects of increased temperature on plant communities depend on landscape location and precipitation. Ecol. Evol. 2018, 8, 5267–5278. [Google Scholar] [CrossRef]
- Gu, Y.; Han, S.J.; Zhang, J.H.; Chen, Z.J.; Wang, W.J.; Feng, Y.; Jiang, Y.G.; Geng, S.C. Temperature-Dominated Driving Mechanisms of the Plant Diversity in Temperate Forests, Northeast China. Forests 2020, 11, 227. [Google Scholar] [CrossRef] [Green Version]
- Paudel, S.K.; Waeber, P.O.; Simard, S.W.; Innes, J.L.; Nitschke, C.R. Multiple factors influence plant richness and diversity in the cold and dry boreal forest of southwest Yukon, Canada. Plant Ecol. 2016, 217, 505–519. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Wipf, S. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P. Competitive exclusion in herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Chu, C.; Lutz, J.A.; Král, K.; Vrška, T.; Yin, X.; Myers, J.A.; Burslem, D.F.R.P. Direct and indirect effects of climate on richness drive the latitudinal diversity gradient in forest trees. Ecol. Lett. 2019, 22, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.W.; Field, R.; Guegan, J.F.; Hawkins, B.A.; Turner, J.R.G. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Storch, D.; Bohdalková, E.; Okie, J. The more-individuals hypothesis revisited: The role of community abundance in species richness regulation and the productivity-diversity relationship. Ecol. Lett. 2018, 21, 920–937. [Google Scholar] [CrossRef]
- O’Brien, E.M. Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; Pelt, R.V.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Chen, J. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. Forest Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Coomes, D.A.; Lines, E.R.; Allen, R.B. Moving on from Metabolic Scaling Theory: Hierarchical models of tree growth and asymmetric competition for light. J. Ecol. 2011, 99, 748–756. [Google Scholar] [CrossRef]
- Lutz, J.A.; Larson, A.J.; Furniss, T.J.; Donato, D.C.; Freund, J.A.; Swanson, M.E. Spatially nonrandom tree mortality and ingrowth maintain equilibrium pattern in an old-growth Pseudotsuga-Tsuga Forest. Ecology 2014, 95, 2047–2054. [Google Scholar] [CrossRef] [Green Version]
- Farrior, C.E.; Bohlman, S.A.; Hubbell, S.; Pacala, S.W. Dominance of the suppressed: Power-law size structure in tropical forests. Science 2016, 351, 155–157. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Han, H.; You, H.; Cheng, X.; Wang, T. Effects of local characteristics and landscape patterns on plant richness: A multi-scale investigation of multiple dispersal traits. Ecol. Indic. 2020, 117, 106584. [Google Scholar] [CrossRef]
- Scheiner, S.M. Six types of species-area curves. Global Ecol. Biogeogr. 2003, 12, 441–447. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: iNterpolation and EXTrapolation for Species Diversity. R Package, Version 3.0.0. 2022. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 13 October 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Wagner, H. Vegan: Community Ecology Package. R Package, Version 2.6–4. 2022. Available online: http://CRAN.R-project.org/package=vegan (accessed on 11 October 2022).
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Elith, J.; Francis, M.P.; Hastie, T.; Taylor, P. Variation in demersal fish species richness in the oceans surrounding New Zealand: An analysis using boosted regression trees. Mar. Ecol. Prog. Ser. 2006, 321, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Zimmermann, N.E. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in tropical rain forests and coral reefs—High diversity of trees and corals is maintained only in a nonequilibrium state. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; He, F.; Zhang, Z.; Zhao, X.; Gadow, K.V. Latitudinal gradients and ecological drivers of beta-diversity vary across spatial scales in a temperate forest region. Global Ecol. Biogeogr. 2020, 29, 1257–1264. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. Large-scale processes and the Asian bias in species diversity of temperate plants. Nature 2000, 407, 180–182. [Google Scholar] [CrossRef]
- Gao, D.; Perry, G. Detecting the small island effect and nestedness of herpetofauna of the West Indies. Ecol. Evol. 2016, 6, 5390–5403. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Kissling, W.D. Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China. Ecology 2010, 91, 1172–1183. [Google Scholar] [CrossRef]
- Schweiger, A.H.; Beierkuhnlein, C. Scale dependence of temperature as an abiotic driver of species’ distributions. Global Ecol. Biogeogr. 2016, 25, 1013–1021. [Google Scholar] [CrossRef]
- Silvertown, J. Plant coexistence and the niche. Trends Ecol. Evol. 2004, 19, 605–611. [Google Scholar] [CrossRef]
- Chen, S.B.; Jiang, G.M.; Ouyang, Z.Y.; Xu, W.H.; Xiao, Y. Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China. J. Syst. Evol. 2011, 49, 95–107. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and Large-Scale Patterns of Animal- and Plant-Species Richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Pfeifer-Meister, L.; Bridgham, S.D.; Reynolds, L.L.; Goklany, M.E.; Wilson, H.E.; Little, C.J.; Ferguson, A.; Johnson, B.R. Climate change alters plant biogeography in Mediterranean prairies along the West Coast, USA. Global Chang. Biol. 2016, 22, 845–855. [Google Scholar] [CrossRef]
- Kikvidze, Z.; Michalet, R.; Brooker, R.W.; Cavieres, L.A.; Lortie, C.J.; Pugnaire, F.I.; Callaway, R.M. Climatic drivers of plant-plant interactions and diversity in alpine communities. Alp. Bot. 2011, 121, 63–70. [Google Scholar] [CrossRef]
- Peyre, G.; Balslev, H.; Font, X.; Tello, J.S. Fine-Scale Plant Richness Mapping of the Andean Paramo According to Macroclimate. Front. Ecol. Evol. 2019, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Geng, Y.; Ma, K. Multi-scale comparison of topographic complexity indices in relation to plant species richness. Ecol. Complex. 2015, 22, 93–101. [Google Scholar] [CrossRef]
- Rahbek, C.; Graves, G.R. Multiscale assessment of patterns of avian species richness. Proc. Natl. Acad. Sci. USA 2001, 98, 4534–4539. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-going, B.M.; Harrison, S.P.; Anacker, B.L.; Safford, H.D. Climate interacts with soil to produce beta diversity in Californian plant communities. Ecology 2013, 94, 2007–2018. [Google Scholar] [CrossRef]
- Harrison, S.; Cornell, H. Toward a better understanding of the regional causes of local community richness. Ecol. Lett. 2008, 11, 969–979. [Google Scholar] [CrossRef]
- Kallimanis, A.S.; Mazaris, A.D.; Tzanopoulos, J.; Halley, J.M.; Pantis, J.D.; Sgardelis, S.P. How does habitat diversity affect the species-area relationship? Global Ecol. Biogeogr. 2008, 17, 532–538. [Google Scholar] [CrossRef]
- Quintero, I.; Jetz, W. Global elevational diversity and diversification of birds. Nature 2018, 555, 246–250. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Richness | Shannon | Simpson | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10 × 10 | 30 × 30 | 90 × 90 | 10 × 10 | 30 × 30 | 90 × 90 | 10 × 10 | 30 × 30 | 90 × 90 | ||
| Climate | AMTmean | 14.0 | 20.0 | 11.7 | 23.7 | 30.2 | 28.1 | 20.9 | 22.2 | 19.7 |
| AMTcv | 13.3 | 10.1 | 10.6 | 9.3 | 14.3 | 28.7 | 8.1 | 16.9 | 46.7 | |
| APmean | 24.5 | 21.6 | 23.6 | 8.2 | 7.4 | 4.3 | 6.4 | 5.1 | 3.3 | |
| ASDmean | 4.8 | 4.4 | 18.7 | 2.6 | 2.2 | 3.3 | 2.6 | 2.2 | 1.0 | |
| ASDcv | 0.9 | 7.9 | 2.0 | 0.3 | 2.9 | 2.5 | 0.2 | 1.2 | 1.1 | |
| Topography | ELmean | 2.5 | 2.9 | 2.5 | 3.8 | 2.4 | 0.7 | 2.6 | 3.8 | 0.1 |
| ELcv | 2.4 | 2.6 | 13.4 | 3.1 | 2.1 | 16.4 | 4.8 | 4.4 | 15.2 | |
| SLmean | 0.5 | 2.9 | 0.3 | 1.6 | 1.8 | 0.5 | 2.7 | 3.9 | 1.5 | |
| SLcv | 3.3 | 1.2 | 3.0 | 1.4 | 1.6 | 1.9 | 3.5 | 1.4 | 0.6 | |
| SMImean | 0.5 | 1.3 | 0.4 | 1.3 | 2.5 | 0.4 | 2.4 | 4.4 | 0.8 | |
| Forest structure | TDmean | 2.8 | 3.0 | 2.9 | 2.5 | 5.5 | 3.3 | 3.4 | 6.5 | 3.1 |
| TDcv | 2.7 | 3.1 | 0.9 | 5.6 | 4.3 | 0.9 | 8.8 | 6.6 | 2.9 | |
| DBHcv | 26.6 | 18.3 | 9.6 | 31.6 | 20.3 | 7.8 | 23.6 | 13.1 | 2.4 | |
| THmean | 1.2 | 0.7 | 0.6 | 4.9 | 2.5 | 1.3 | 10.1 | 8.3 | 1.6 | |
| Model parameters | Distribution | Poisson | Poisson | Poisson | Gaussian | Gaussian | Gaussian | Gaussian | Gaussian | Gaussian |
| TC | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| LR | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | |
| R2 | 0.65 | 0.77 | 0.85 | 0.69 | 0.80 | 0.86 | 0.62 | 0.73 | 0.69 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, Y.; Zhang, J.; Ma, W.; Feng, Y.; Yang, L.; Li, Z.; Guo, Y.; Shi, G.; Han, S. Ecological Factors Driving Tree Diversity across Spatial Scales in Temperate Forests, Northeast China. Forests 2023, 14, 1241. https://doi.org/10.3390/f14061241
Gu Y, Zhang J, Ma W, Feng Y, Yang L, Li Z, Guo Y, Shi G, Han S. Ecological Factors Driving Tree Diversity across Spatial Scales in Temperate Forests, Northeast China. Forests. 2023; 14(6):1241. https://doi.org/10.3390/f14061241
Chicago/Turabian StyleGu, Yue, Junhui Zhang, Wang Ma, Yue Feng, Leilei Yang, Zhuo Li, Yanshuang Guo, Guoqiang Shi, and Shijie Han. 2023. "Ecological Factors Driving Tree Diversity across Spatial Scales in Temperate Forests, Northeast China" Forests 14, no. 6: 1241. https://doi.org/10.3390/f14061241