
Rethinking Iconic Species Reforestation in West Africa: Seed Shape Harnessing Is Strategic for Enhanced Germination and Vigorous Growth in Khaya senegalensis and Parkia biglobosa

 ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Methods
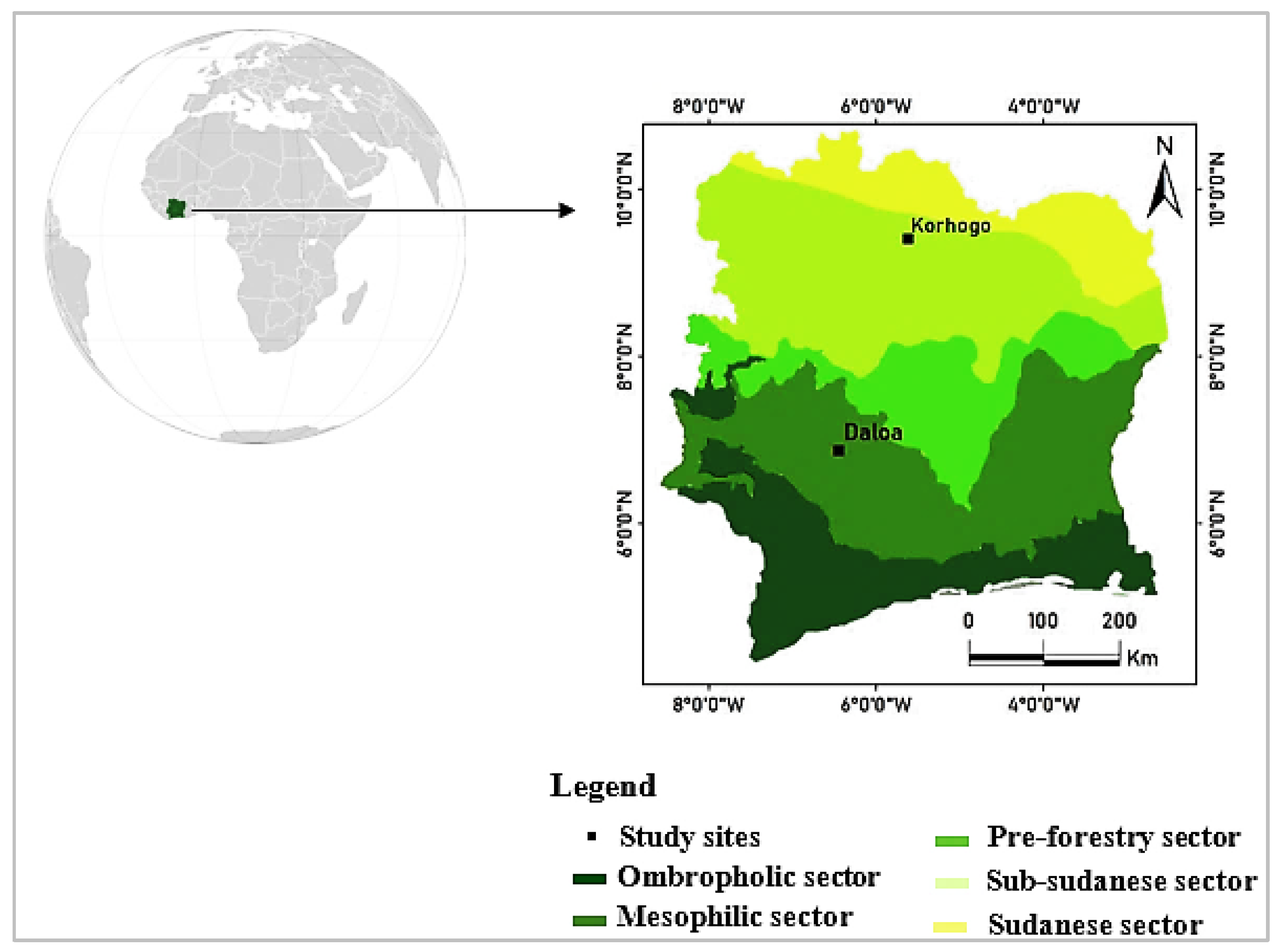
2.2.1. Experimental Sites
2.2.2. Collecting and Grading Seeds
2.2.3. Setting up Trials and Sowing
2.2.4. Measurements
- -
- Waiting, or latency, time: this is the time in days needed for the first seed to germinate in a batch set (number of days needed for the first germination);
- -
- Germination delay: the time in days between sowing and germination for a given seed in a set of lots (number of days needed for germination of each seed sown);
- -
- Germination speed: the average time in days until 50% of the seeds in a batch germinate (number of days needed for half the seeds sown to germinate);
- -
- Germination duration or spread: the time in days between the first and last germination of a batch (number of days between the first and last germination);
- -
- Germination rate: the proportion of germinated seeds versus the total number of seeds sown. This is expressed as a percentage (ratio of the number of germinated seeds over the number of sown seeds, as a percentage).
2.2.5. Seedling Growth and Mortality
2.2.6. Statistical Analyses
3. Results
3.1. Seed Germination or Emergence
3.1.1. Germination in Khaya Senegalensis
3.1.2. Germination in Parkia Biglobosa
3.2. Seedling Growth
3.2.1. Effect of Seed Size and Site on Seedling Growth in Khaya Senegalensis
3.2.2. Effect of Seed Size and Site on Seedling Growth in Parkia Biglobosa
3.2.3. Assessment of Mortality in 12-Month-Old Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Amani, B.H.K.; N’Guessan, A.E.; Meersch, V.V.d.; Derroire, G.; Piponiot, C.; Elogne, A.G.M.; Traoré, K.; N’Dja, J.K.; Hérault, B. Lessons from a regional analysis of forest recovery trajectories in West Africa. Environ. Res. Lett. 2022, 17, 115005. [Google Scholar] [CrossRef]
- Bonn, C. Forest Landscape Restoration. 2016. Available online: http://www.bonnchallenge.org/content/c%C3%B4te-d-ivoire (accessed on 17 May 2022).
- IUCN. World Conservation Monitoring Centre. Khaya senegalensis. The IUCN Red List of Threatened Species. 1998, e.T32171A9684583. Available online: https://www.iucnredlist.org/species/32171/9684583 (accessed on 17 May 2022).
- IUCN. The IUCN Red List of Threatened Species. Version 2020-2. 2020. Available online: https://www.iucnredlist.org (accessed on 17 May 2022).
- Adji, B.I.; Akaffou, D.S.; De Reffye, P.; Sabatier, S. Maternal environment and seed size are important for successful germination and seedling establishment of Pterocarpus erinaceus (Fabaceae). J. For. Res. 2021, 33, 977–990. [Google Scholar] [CrossRef]
- Hérault, B.; Anatole, K.N.; N’klo, O.; Assandé, A.; Fabrice, B.; Brahima, C.; Doua-Bi, Y.; Koffi, Y.; Koffi, K.J.C.; Konaté, I.; et al. The Long-Term Performance of 35 Tree Species of Sudanian West Africa in Pure and Mixed Plantings. For. Ecol. Manag. 2020, 468, 118171. [Google Scholar] [CrossRef]
- Adji, B.I.; Akaffou, S.D.; Kouassi, K.H.; Houphouet, Y.P.; Duminil, J.; Sabatier, S. Influence of different environments on germination parameters and seedling morphology in Khaya senegalensis (Desr.) A. Juss (Meliaceae). Am. J. Plant Sci. 2020, 11, 1579–1600. [Google Scholar] [CrossRef]
- Adji, B.I.; Akaffou, S.D.; Kouassi, K.H.; Houphouet, Y.P.; De Reffye, P.; Duminil, J.; Jaeger, M.; Sabatier, S. Allometric models for non-destructive estimation of dry biomass and leaf area in Khaya senegalensis (Desr.) A. Juss., 1830 (Meliaceae), Pterocarpus erinaceus Poir., 1804 (Fabaceae) and Parkia biglobosa (Jacq.) R. Br., 1830 (Fabaceae). Trees 2021, 35, 1905–1920. [Google Scholar] [CrossRef]
- Assane, M.; Baba, R.M.; Bassene, E.; Sere, A. Antihypertensive action of Parkia biglobosa (Jacq) Benth seeds in the rat. Dakar Med. 1993, 38, 49–54. [Google Scholar]
- Chacón, P.; Bustamante, R.O.; Henriquez, C. The effect of seed size on germination and seedling growth of Cryptocarya alba (Lauraceae) in Chile. Rev. Chil. Hist. Nat. 2001, 71, 189–197. [Google Scholar] [CrossRef]
- Mao, P.; Guo, L.; Gao, Y.; Qi, L.; Cao, B. Effects of Seed Size and Sand Burial on Germination and Early Growth of Seedlings for Coastal Pinus thunbergii Parl. in the Northern Shandong Peninsula, China. Forests 2019, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Agboola, D.A. The effect of seed size on germination and seedling growth of three tropical tree species. J. Trop. For. Sci. 1996, 9, 44–51. [Google Scholar]
- Mtambalika, K.; Munthali, C.; Gondwe, D.; Missanjo, E. Effect of Seed Size of Afzelia quanzensis on Germination and Seedling Growth. Int. J. For. Res. 2014, 2014, 384565. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.L. Effects of seed mass on seedling success in Artocarpus heterophyllus 381 L., a tropical tree species of north-east India. Acta Oecol. 2004, 25, 103–110. [Google Scholar] [CrossRef]
- Fornah, R.Y.; Mattia, S.M.; Otesile, A.A.; Ernest, G.; Kamara, E.G. Effects of Provenance and Seed Size on Germination, Seedling Growth and Physiological Traits of Gmelina arborea. Int. J. Agric. For. 2017, 7, 28–34. [Google Scholar]
- Guillaumet, J.L.; Adjanohoun, E.; Avenard, J.M.; Eldin, M.; Girard, G.; Circoulin, J.; Toucheboeuf, P.; Perraud, A. Le milieu naturel de Côte d’Ivoire. Mém. ORSTOM no50. ORSTOM, Paris. 1971, pp. 156–264. Available online: https://www.documentation.ird.fr/hor/fdi:16372 (accessed on 17 May 2022).
- Louppe, D.; Ouattara, N. Les arboretums d’espèces locales en Nord Côte d’Ivoire. IDEFOR. 1996, p. 14. Available online: http://agritrop.cirad.fr/581418/ (accessed on 17 May 2022).
- Akaffou, S.D.; Kouamé, K.A.; Gore, B.B.N.; Abessika, Y.G.; Kouassi, K.H.; Hamon, P.; Sabatier, S.; Duminil, J. Effect of the seeds provenance and treatment on the germination rate and plants growth of four forest trees species of Côte d’Ivoire. J. For. Res. 2019, 32, 161–169. [Google Scholar] [CrossRef]
- Toledo-Aceves, T. Germination rate of endangered cloud forest trees in Mexico: Potential for ex situ propagation. J. For. Res. 2017, 22, 61–64. [Google Scholar] [CrossRef]
- Iralu, V.; Upadhaya, K. Seed dormancy, germination and seedling characteristics of Elaeocarpus prunifolius Wall. ex Müll. Berol.: A threatened tree species of north-eastern India. N. Z. J. For. Sci. 2018, 48, 16. [Google Scholar] [CrossRef]
- Burslem, D.F.R.P.; Miller, J. Seed size, germination and seedling relative growth rates in three tropical tree species. J. Trop. For. Sci. 2001, 13, 148–161. [Google Scholar]
- Deb, P.; Sundriyal, R.C. Effect of seeds size on germination and seedling fitness on four tropical rainforest tress species. Ind. J. For. 2014, 40, 313–322. [Google Scholar]
- Kuniyal, C.P.; Purohit, V.; Butola, J.S.; Sundriyal, R.C. Seed size correlates seedlings emergence in Terminalia bellerica. S. Afr. J. Bot. 2013, 87, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.M.; Chakraborty, D.; Gupta, K. Seeds size variation: Influence on germination and subsequent seedlings performance in Hyptis suaveolens (Lamiaceae). Res. J. Seed Sci. 2008, 322, 26–33. [Google Scholar]
- Souza, M.L.; Fagundes, M. Seed size as key factor in germination and seedling development of copment of Copaifera langsdorffii (Fabaceae). Am. J. Plant Sci. 2014, 5, 2566–2573. [Google Scholar] [CrossRef] [Green Version]
- Simão, E.; Takaki, M. Effect of light and temperature on seed germination in Tibouchina mutabilis (Vell.) Cogn. (Melastomataceae). Biota Neotrop. 2008, 8, 63–68. Available online: http://www.biotaneotropica.org.br/v8n2/pt/abstract?article+bn00908022008 (accessed on 17 May 2022). [CrossRef]
- Vargas-Figueroa, J.A.; Torres-González, A.M. Germination and seed conservation of a pioneer species, Tecoma stans (Bignoniaceae), from tropical dry forest of Colombia. Rev. Biol. Trop. Int. J. Trop. Biol. Conserv. 2018, 66, 918–936. [Google Scholar] [CrossRef] [Green Version]
- Nakar, R.N.; Jadeja, B.A. Seed pattern, germination and viability of some tree species seeds from Girnar Reserve Forest of Gujarat. Ind. J. Plant Physiol. 2014, 19, 57–64. [Google Scholar] [CrossRef]
- Ellis, R.H. Seed and seedling vigor in relation to crop growth and yield. J. Plant Growth Regul. 1992, 11, 249–255. [Google Scholar] [CrossRef]
- Milberg, P.; Lamont, B.B. Seed/cotyledon size and nutrient content play a major role in early performance of species on nutrient-poor soils. Wiley Online Libr. N. Phytol. 1997, 137, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Fredrick, C.; Muthuri, C.; Ngamau, K.; Sinclair, F. Provenance and pretreatment effect on seed germination of six provenances of Faidherbia albida (Delile) A. Chev. Agrofor. Syst. 2017, 91, 1007–1017. [Google Scholar] [CrossRef]
- Adji, B.I.; Akaffou, D.S.; Sabatier, S. Variation in growth unit morphology in Khaya senegalensis (Desr.) A. Juss. (Meliaceae) and Pterocarpus erinaceus Poir. (Fabaceae) according to habitat and climate. Bois Et. Des. Trop. 2022, 354, 41–54. [Google Scholar] [CrossRef]
- Wang, H.; Wang, X.; Li, Y.; Gao, R.; Rao, M.P.N.; Song, J.; Li, Q. Effect of environmental factors on seed germination and seedling emergence of Viola prionantha, a cleistogamous plant. J. Plant Res. 2023. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Provenances | GPS Coordinates | Number of Seed Trees | DBH (cm) | Height (m) | Number of Healthy Seeds | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Longitude | Latitude | Min | Max | Mean | Min | Max | Mean | ||||
| Khaya senegalensis | Katiola | 5°7′35.814″ W | 8°13′53.94″ N | 7 | 43.12 | 87.6 | 63.6 ± 22.1 b | 13.5 | 35 | 22.64 ± 6.8 b | 426.61 ± 58.3 b |
| Niakara | 5°18′40.735″ W | 8°40′47.97″ N | 10 | 57.34 | 108.65 | 72.34 ± 19.3 ab | 16.3 | 28.5 | 18.31 ± 7.42 b | 793.4 ± 78.6 a | |
| Korhogo | 5°36′12.316″ W | 9°33′24.69″ N | 10 | 55.93 | 98.22 | 69.91 ± 20.5 ab | 24.5 | 39.5 | 28.94 ± 8.78 a | 804 ± 104.53 a | |
| Ferké | 5°23′43.396″ W | 9°36′1.87″ N | 9 | 47.69 | 126.77 | 88.3 ± 48.4 a | 21.6 | 42.5 | 31.52 ± 11.2 a | 856.11 ± 153.5 a | |
| Pr > F | 0.0131 | 0.021 | 0.001 | ||||||||
| Parkia biglobosa | Katiola | 5°7′35.814″ W | 8°13′53.94″ N | 15 | 18.4 | 47.23 | 21.84 ± 5.1 b | 6.5 | 23.6 | 12.54 ± 3.3 b | 434 ± 87.5 b |
| Niakara | 5°18′40.735″ W | 8°40′47.97″ N | 14 | 20.6 | 55.81 | 26.7 ± 7.34 ab | 8.5 | 27.5 | 20.1 ± 6.5 a | 1026 ± 209.6 a | |
| Korhogo | 5°36′12.316″ W | 9°33′24.69″ N | 20 | 19.7 | 63.33 | 33.4 ± 11.3 a | 11.5 | 26.5 | 18.77 ± 6.2 a | 1009 ± 303.7 a | |
| Ferké | 5°23′43.396″ W | 9°36′1.87″ N | 12 | 23.56 | 38.11 | 28.7 ± 8.6 a | 13.7 | 21.6 | 17.64 ± 8.32 ab | 579.34 ± 179.82 b | |
| Pr > F | 0.016 | 0.037 | 0.001 | ||||||||
| Study Sites | Coordinates | Vegetation | Climate | Temperature (°C) | Rainfall (mm/y) | RH (%) | Soil Type |
|---|---|---|---|---|---|---|---|
| Korhogo | 9°570′80556″ N; 5°542′88889″ W | Clear open forest (wooded and grassy savannah) | Tropical dry (Sudano-dry zone) | 26.6–35.7 | 817–1216 | 20–80 | Ferruginous (80%) and ferralitic (20%); superficial gravelly acidic soil, deep gravel with a heavy texture (sandy loamy clay), low in organic matter, highly desaturated, low in calcium, magnesium, potassium and phosphorus |
| Daloa | 6°909′6363″ N 6°438′1157″ W | Dense rain forest (Dense humid vegetation) | Wet tropical (subequatorial; Guinean-humid zone) | 21.0–27.2 | 1000–1900 | 42–95 | Ferralitic, deep, acidic and desaturated in exchangeable bases, rich in organic matter (calcium, magnesium, potassium and phosphorus) |
| Species | Seed Size | Mass (g) | Length (mm) | Width (mm) | Thickness (mm) |
|---|---|---|---|---|---|
| Khaya senegalensis | Large Min-Maxi | 0.31 ± 0.03 a (0.25–0.37) | 30.7 ± 3.2 a (23–36) | 21.2 ± 2.3 a (17–25) | 2.6 ± 0.38 a (1.79–3.2) |
| Medium Min-Maxi | 0.21 ± 0.02 b (0.17–0.24) | 23.7 ± 3.6 b (18–32) | 17.3 ± 2.4 b (14–22) | 2.37 ± 0.43 a (1.61–3.59) | |
| Small Min-Maxi | 0.1 ± 0.03 c (0.05–0.16) | 19.4 ± 2.4 c (14–24) | 14.6 ± 3.1 c (11–22) | 1.95 ± 0.55 a (0.7–2.97) | |
| Pr > F | 0.001 | 0.001 | 0.02 | 0.194 | |
| Parkia biglobosa | Large Min-Maxi | 0.27 ± 0.03 a (0.2–0.32) | 11.1 ± 0.9 a (10–13) | 7.9 ± 0.9 a (6–11) | 5.54 ± 0.42 a (4.88–6.4) |
| Medium Min-Maxi | 0.17 ± 0.01 b (0.16–0.19) | 9 ± 0.8 a b (8–11) | 6.7 ± 0.7 a b (5–8) | 5.13 ± 0.34 a (4.61–5.99) | |
| Small Min-Maxi | 0.12 ± 0.05 c (0.04–0.13) | 7.2 ± 0.8 c (5–9) | 5.5 ± 0.9 c (4–7) | 4.71 ± 0.62 a (3.42–5.89) | |
| Pr > F | 0.002 | 0.031 | 0.004 | 0.182 |
| Development Parameters | Height (cm) | Diameter (mm) | Number of Single Leaves | Number of Compound Leaves | Root Length (cm) | Root Diameter (mm) |
|---|---|---|---|---|---|---|
| Large | 17.77 ± 3.92 a | 3.07 ± 2.6 a | 7.43 ± 1.66 a | 0.4 ± 1.1 a | 13.90 ± 2.87 a | 2.97 ± 0.52 a |
| Medium | 12.36 ± 2.08 b | 2.07 ± 0.33 b | 5.67 ± 1.17 b | 0.11 ± 0.46 b | 11.6 ± 2.38 b | 2.23 ± 0.39 b |
| Small | 8.28 ± 2.11 c | 1.6 ± 0.35 c | 4.43 ± 0.85 c | 0 ± 0 c | 8.78 ± 2.43 c | 1.83 ± 0.44 b |
| Pr > F | 0.0001 | 0.0001 | 0.0041 | 0.0007 | 0.0001 | 0.0063 |
| Dry Biomass Parameters | Total Dry Mass of Seedlings (g) | Total Dry Mass of Leaves (g) | Stem Dry Mass (g) | Root Dry Mass (g) | ||
| Large | 0.71 ± 0.25 a | 0.32 ± 0.11 a | 0.48 ± 0.19 a | 0.14 ± 0.05 a | ||
| Medium | 0.41 ± 0.12 b | 0.20 ± 0.06 b | 0.32 ± 1.52 b | 0.09 ± 0.04 b | ||
| Small | 0.22 ± 0.11 c | 0.1 ± 0.05 c | 0.04 ± 0.02 c | 0.05 ± 0.02 c | ||
| Pr > F | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||
| Development Parameters | Seedling Height (cm) | Stem Diameter (mm) | Number of Single Leaves | Number of Compound Leaves | Root Length (cm) | Root Diameter (mm) |
|---|---|---|---|---|---|---|
| Daloa | 15.85 ± 5.07 a | 2.76 ± 2.39 a | 6.88 ± 1.87 a | 0.35 ± 1.02 a | 12.78 ± 3.71 a | 2.74 ± 0.64 a |
| Korhogo | 10.93 ± 3.12 b | 1.91 ± 0.44 b | 5.18 ± 1.25 b | 0.04 ± 0.26 b | 10.7 ± 2.48 b | 2.09 ± 0.51 a |
| Pr > F | 0.0001 | 0.0002 | 0.0075 | 0.0001 | 0.0069 | 0.076 |
| Dry Biomass Parameters | Total Dry Mass of Seedlings (g) | Total Dry Mass of Le aves (g) | Stem Dry Mass (g) | Root Dry Mass (g) | ||
| Daloa | 0.61 ± 0.29 a | 0.26 ± 0.13 a | 0.29 ± 1.25 a | 0.12 ± 0.06 a | ||
| Korhogo | 0.24 ± 0.16 b | 0.18 ± 0.09 b | 0.08 ± 0.04 b | 0.08 ± 0.04 b | ||
| Pr > F | 0.0001 | 0.0001 | 0.0001 | 0.0001 | ||
| Species | Seed Size Sources | Height (cm) | Collar Diameter (cm) | Death Rate (%) |
|---|---|---|---|---|
| Khaya senegalensis | Larges | 167.11 ± 18.3 a | 3.3 ± 0.2 a | 0 |
| Mediums | 142.7 ± 79.1 a | 2.07 ± 0.53 a | 47.2 | |
| Small | 89.52 ± 56.66 a | 2.02 ± 1.37 a | 72 | |
| Pr > F | 0.603 | 0.83 | - | |
| Parkia biglobosa | Larges | 61.25 ± 23.8 a | 0.7 ± 0.41 a | 32.5 |
| Mediums | 56.57 ± 17.1 a | 0.68 ± 0.39 a | 53.33 | |
| Small | 54.1 ± 33.3 a | 0.66 ± 0.51 a | 64.8 | |
| Pr > F | 0.874 | 0.711 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adji, B.I.; Letort, V.; Wang, X.; Kang, M.; De Reffye, P.; Jaeger, M.; Cilas, C.; Kouassi, K.H.; Duminil, J.; Sabatier, S.; et al. Rethinking Iconic Species Reforestation in West Africa: Seed Shape Harnessing Is Strategic for Enhanced Germination and Vigorous Growth in Khaya senegalensis and Parkia biglobosa. Forests 2023, 14, 1311. https://doi.org/10.3390/f14071311
Adji BI, Letort V, Wang X, Kang M, De Reffye P, Jaeger M, Cilas C, Kouassi KH, Duminil J, Sabatier S, et al. Rethinking Iconic Species Reforestation in West Africa: Seed Shape Harnessing Is Strategic for Enhanced Germination and Vigorous Growth in Khaya senegalensis and Parkia biglobosa. Forests. 2023; 14(7):1311. https://doi.org/10.3390/f14071311
Chicago/Turabian StyleAdji, Beda Innocent, Véronique Letort, Xiujuan Wang, Mengzhen Kang, Philippe De Reffye, Marc Jaeger, Christian Cilas, Kouadio Henri Kouassi, Jérôme Duminil, Sylvie Sabatier, and et al. 2023. "Rethinking Iconic Species Reforestation in West Africa: Seed Shape Harnessing Is Strategic for Enhanced Germination and Vigorous Growth in Khaya senegalensis and Parkia biglobosa" Forests 14, no. 7: 1311. https://doi.org/10.3390/f14071311