
Niche and Interspecific Association of Dominant Tree Species in Spruce–Fir Mixed Forests in Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Location Overview of Study Area
2.2. Data Sources
2.3. Data Analysis
2.3.1. Calculation of Important Values
2.3.2. Calculation of Niche Breadth and Overlap Index
2.3.3. Overall Connectivity Test
2.3.4. Test for Interspecific Correlation
2.3.5. Data Analysis
3. Results
3.1. Importance Value and Niche Breadth of Dominant Tree Species
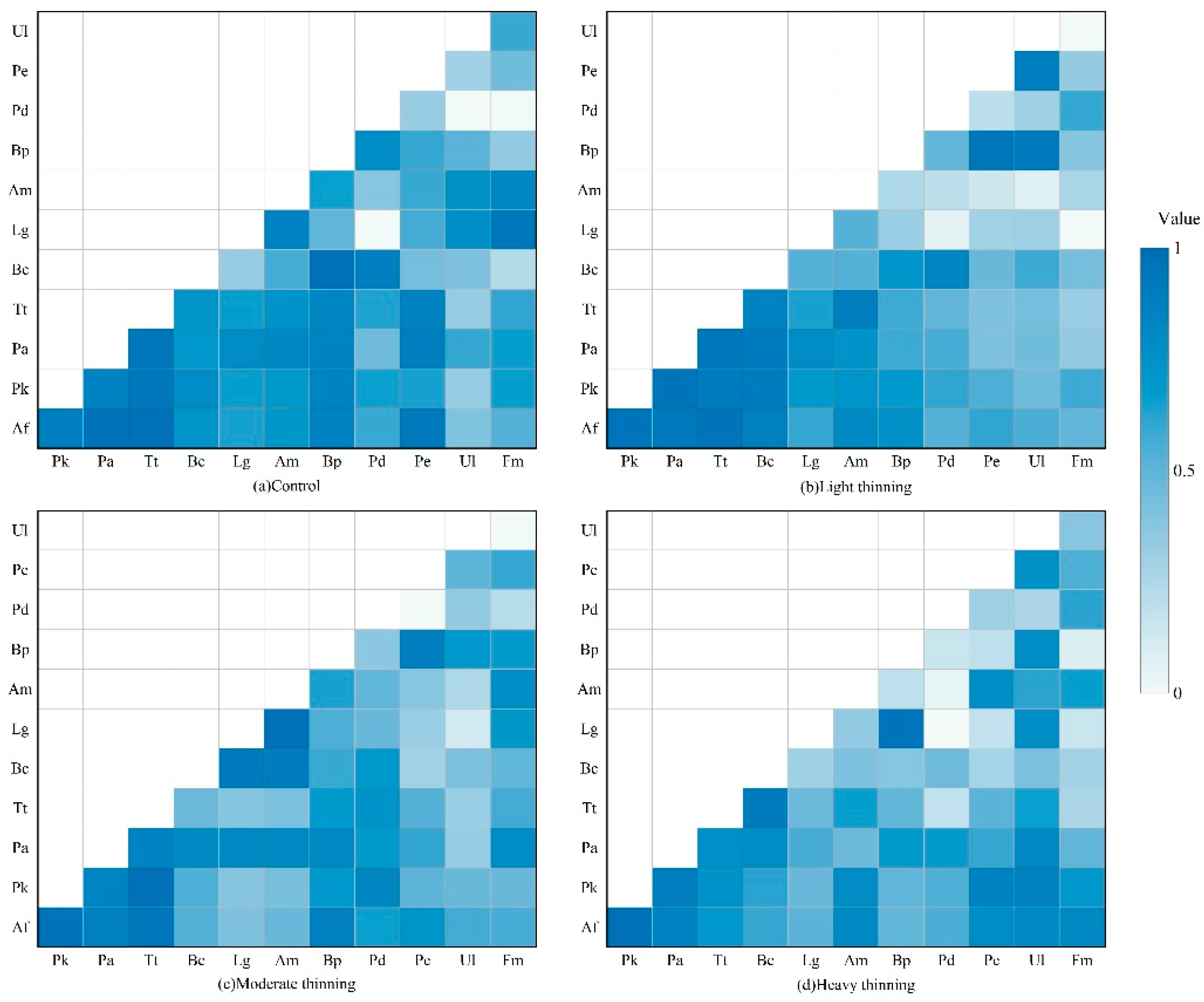
3.2. Niche Overlap of Dominant Tree Species
3.3. Overall Interspecies Association
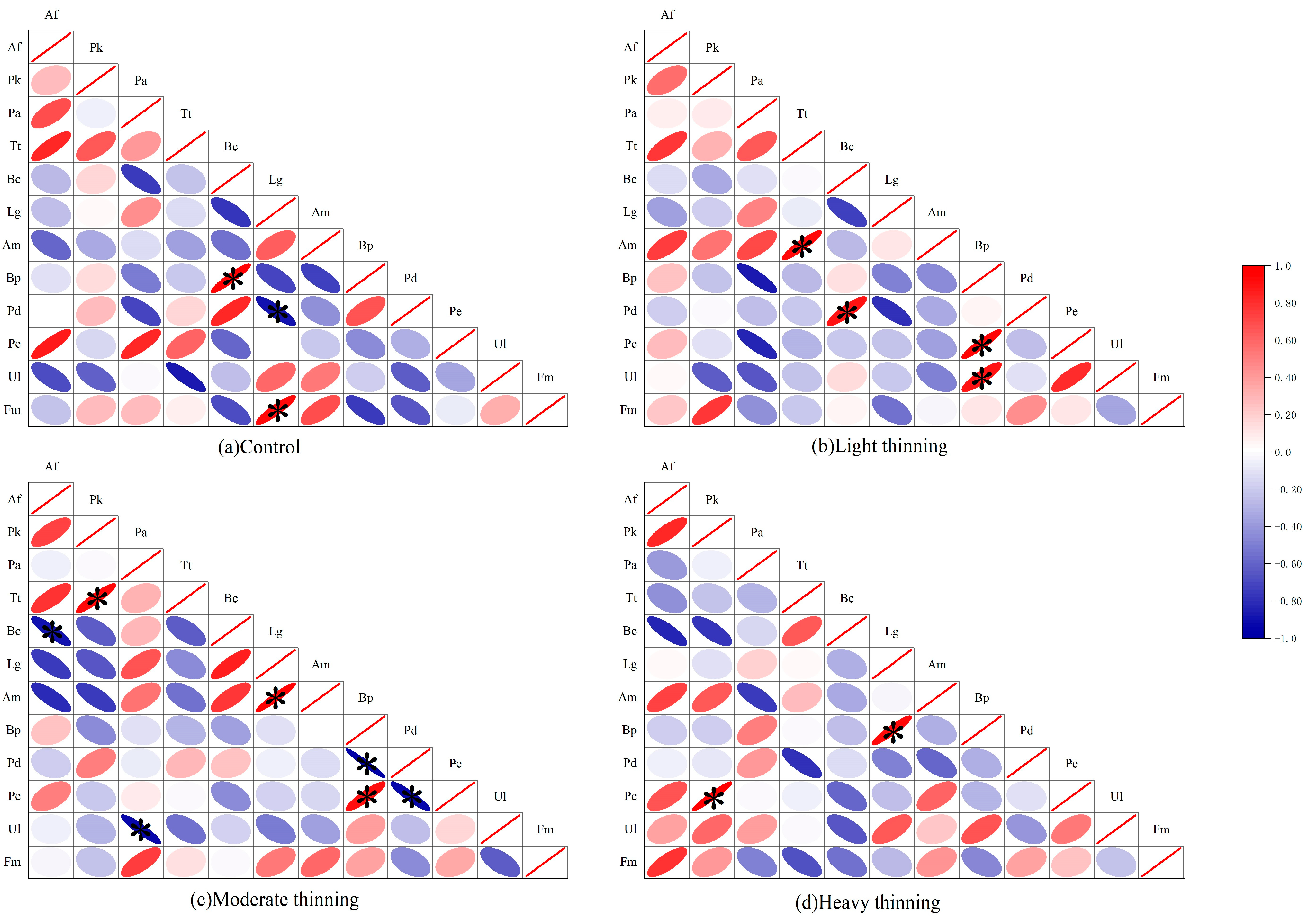
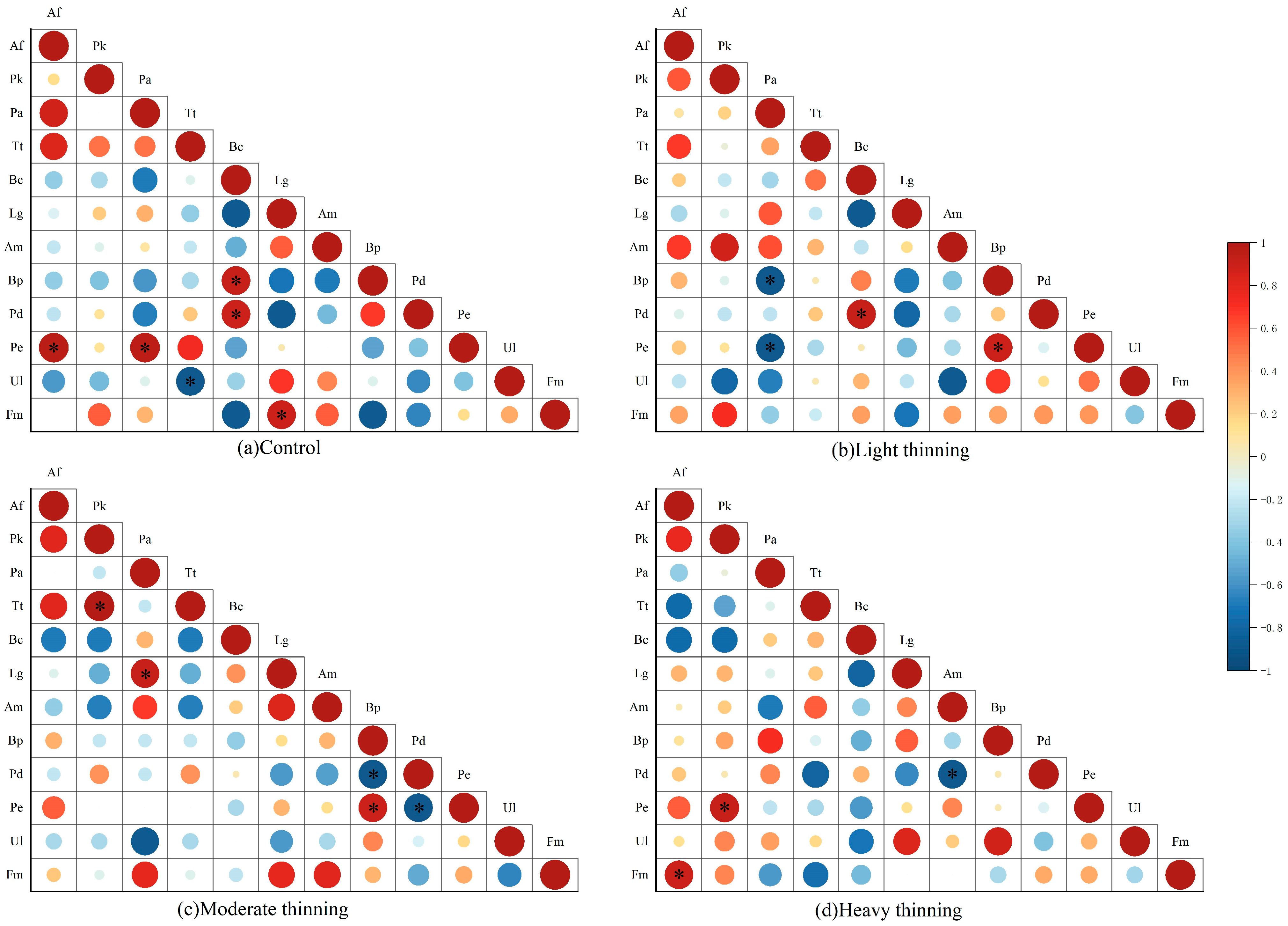
3.4. Interspecific Correlation Analysis
4. Discussion
4.1. Niche Characteristics of Dominant Tree Species
4.2. Interspecific Correlation Characteristics of Dominant Tree Species
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterson, A.T. Predicting the Geography of Species’ Invasions via Ecological Niche Modeling. Q. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The Role of Functional Traits and Trade-Offs in Structuring Phytoplankton Communities: Scaling from Cellular to Ecosystem Level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Buche, L.; Spaak, J.W.; Jarillo, J.; De Laender, F. Niche Differences, Not Fitness Differences, Explain Predicted Coexistence across Ecological Groups. J. Ecol. 2022, 110, 2785–2796. [Google Scholar] [CrossRef]
- Petraitis, P.S. Likelihood Measures of Niche Breadth and Overlap. Ecology 1979, 60, 703–710. [Google Scholar] [CrossRef]
- Ingram, T.; Costa-Pereira, R.; Araújo, M.S. The Dimensionality of Individual Niche Variation. Ecology 2018, 99, 536–549. [Google Scholar] [CrossRef]
- Su, S.; Liu, J.; He, Z.; Zheng, S.; Hong, W.; Xu, D. Ecological Species Groups and Interspecific Association of Dominant Tree Species in Daiyun Mountain National Nature Reserve. J. Mt. Sci. 2015, 12, 637–646. [Google Scholar] [CrossRef]
- Lancaster, L.T. On the Macroecological Significance of Eco-Evolutionary Dynamics: The Range Shift–Niche Breadth Hypothesis. Phil. Trans. R. Soc. B 2022, 377, 20210013. [Google Scholar] [CrossRef]
- Detto, M.; Levine, J.M.; Pacala, S.W. Maintenance of High Diversity in Mechanistic Forest Dynamics Models of Competition for Light. Ecol. Monogr. 2022, 92, e1500. [Google Scholar] [CrossRef]
- Falk, D.A.; van Mantgem, P.J.; Keeley, J.E.; Gregg, R.M.; Guiterman, C.H.; Tepley, A.J.; Young, D.J.N.; Marshall, L.A. Mechanisms of Forest Resilience. For. Ecol. Manag. 2022, 512, 120129. [Google Scholar] [CrossRef]
- Löffler, J.; Pape, R. Thermal Niche Predictors of Alpine Plant Species. Ecology 2020, 101, e02891. [Google Scholar] [CrossRef] [Green Version]
- Cassini, M.H. A Review of the Critics of Invasion Biology. Biol. Rev. 2020, 95, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, Q.; Pan, S.; Liu, C.; Han, M.; Brancelj, A. Niche and Interspecific Associations of Pseudoanabaena Limnetica—Exploring the Influencing Factors of Its Succession Stage. Ecol. Indic. 2022, 138, 108806. [Google Scholar] [CrossRef]
- Zhang, L.-N.; Zhu, G.-Q.; Yang, K.; Liu, X.-Y.; Gong, H.-D.; Zheng, L. Niche and interspecific association of main woody plants in Myrica nana shrubland in central Yunnan, China. Chin. J. Plant Ecol. 2022, 46, 1400–1410. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of Stand Age, Richness and Density on Productivity in Subtropical Forests in China. J. Ecol. 2019, 107, 2266–2277. [Google Scholar] [CrossRef]
- Lu, D.; Zhu, J.; Wang, X.; Hao, G.; Wang, G.G. A Systematic Evaluation of Gap Size and within-Gap Position Effects on Seedling Regeneration in a Temperate Secondary Forest, Northeast China. For. Ecol. Manag. 2021, 490, 119140. [Google Scholar] [CrossRef]
- Manlick, P.J.; Pauli, J.N. Human Disturbance Increases Trophic Niche Overlap in Terrestrial Carnivore Communities. Proc. Natl. Acad. Sci. USA 2020, 117, 26842–26848. [Google Scholar] [CrossRef]
- O’Neil, S.T.; Coates, P.S.; Brussee, B.E.; Ricca, M.A.; Espinosa, S.P.; Gardner, S.C.; Delehanty, D.J. Wildfire and the Ecological Niche: Diminishing Habitat Suitability for an Indicator Species within Semi-arid Ecosystems. Glob. Change Biol. 2020, 26, 6296–6312. [Google Scholar] [CrossRef]
- Koutsidi, M.; Moukas, C.; Tzanatos, E. Trait-based Life Strategies, Ecological Niches, and Niche Overlap in the Nekton of the Data-poor Mediterranean Sea. Ecol. Evol. 2020, 10, 7129–7144. [Google Scholar] [CrossRef]
- Collins, S.L.; Good, R.E. The Seedling Regeneration Niche: Habitat Structure of Tree Seedlings in an Oak-Pine Forest. Oikos 1987, 48, 89. [Google Scholar] [CrossRef] [Green Version]
- Feinsinger, P.; Spears, E.E.; Poole, R.W. A Simple Measure of Niche Breadth. Ecology 1981, 62, 27–32. [Google Scholar] [CrossRef]
- Pianka, E.R. Niche Overlap and Diffuse Competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [PubMed]
- Charles, A.; Darné, O. Variance-Ratio Tests of Random Walk: An Overview. J. Econ. Surv. 2009, 23, 503–527. [Google Scholar] [CrossRef] [Green Version]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation Coefficients: Appropriate Use and Interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Zar, J.H. Significance Testing of the Spearman Rank Correlation Coefficient. J. Am. Stat. Assoc. 1972, 67, 578–580. [Google Scholar] [CrossRef]
- Cameron, A.D. Importance of Early Selective Thinning in the Development of Long-Term Stand Stability and Improved Log Quality: A Review. Forestry 2002, 75, 25–35. [Google Scholar] [CrossRef]
- Lu, X.; Cheng, G.; Xiao, F.; Fan, J. Modeling Effects of Temperature and Precipitation on Carbon Characteristics and GHGs Emissions in Abies Fabric Forest of Subalpine. J. Environ. Sci. 2008, 20, 339–346. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ying, C.C.; Spittlehouse, D.L.; Hamilton, D.A., Jr. Genetic Responses to Climate in Pinus Contorta: Niche Breadth, Climate Change, and Reforestation. Ecol. Monogr. 1999, 69, 375–407. [Google Scholar] [CrossRef]
- Hethcoat, M.G.; Edwards, D.P.; Carreiras, J.M.B.; Bryant, R.G.; França, F.M.; Quegan, S. A Machine Learning Approach to Map Tropical Selective Logging. Remote Sens. Environ. 2019, 221, 569–582. [Google Scholar] [CrossRef]
- Rosas, Y.M.; Peri, P.L.; Lencinas, M.V.; Martínez Pastur, G. Potential Biodiversity Map of Understory Plants for Nothofagus Forests in Southern Patagonia: Analyses of Landscape, Ecological Niche and Conservation Values. Sci. Total Environ. 2019, 682, 301–309. [Google Scholar] [CrossRef]
- Mason, N.W.H.; De Bello, F.; Doležal, J.; Lepš, J. Niche Overlap Reveals the Effects of Competition, Disturbance and Contrasting Assembly Processes in Experimental Grassland Communities: Grassland Community Assembly Processes. J. Ecol. 2011, 99, 788–796. [Google Scholar] [CrossRef]
- Pandey, R.; Rawat, M.; Singh, V.; Yousefpour, R.; Reshi, Z.A. Large Scale Field-Based Evaluation of Niche Breadth, Niche Overlap and Interspecific Association of Western Himalayan Temperate Forest Tree Species. Ecol. Indic. 2023, 146, 109876. [Google Scholar] [CrossRef]
- Cazzolla Gatti, R.; Castaldi, S.; Lindsell, J.A.; Coomes, D.A.; Marchetti, M.; Maesano, M.; Di Paola, A.; Paparella, F.; Valentini, R. The Impact of Selective Logging and Clearcutting on Forest Structure, Tree Diversity and above-Ground Biomass of African Tropical Forests. Ecol. Res. 2015, 30, 119–132. [Google Scholar] [CrossRef]
- Ahmad, M.; Sharma, P.; Rathee, S.; Singh, H.P.; Batish, D.R.; Lone, G.R.; Kaur, S.; Jaryan, V.; Kohli, R.K. Niche Width Analyses Facilitate Identification of High-Risk Endemic Species at High Altitudes in Western Himalayas. Ecol. Indic. 2021, 126, 107653. [Google Scholar] [CrossRef]
- Wu, S.; Wen, L.; Dong, S.; Gao, X.; Xu, Y.; Li, S.; Dong, Q.; Wessell, K. The Plant Interspecific Association in the Revegetated Alpine Grasslands Determines the Productivity Stability of Plant Community Across Restoration Time on Qinghai–Tibetan Plateau. Front. Plant Sci. 2022, 13, 850854. [Google Scholar] [CrossRef]
- Yang, H.; Miao, N.; Li, S.; Ma, R.; Liao, Z.; Wang, W.; Sun, H. Relationship between Stand Characteristics and Soil Properties of Two Typical Forest Plantations in the Mountainous Area of Western Sichuan, China. J. Mt. Sci. 2019, 16, 1816–1832. [Google Scholar] [CrossRef]
- Mikusin, G. Strengthening the Network of High Conservation Value Forests in Boreal Landscapes. Front. Ecol. Evol. 2021, 8, 486. [Google Scholar]
- Akatov, V.V.; Akatova, T.V.; Chefranov, S.G. Degree of Dominance and Species Richness in Plant Communities with High and Low Intensity of Interspecies Competition. Biol. Bull. Rev. 2018, 8, 389–400. [Google Scholar] [CrossRef]
- Dalmaso, C.A.; Inoue, M.T.; Marcelino, V.R.; Oliveira Filho, P.C.D.; Nascimento, A.R.T.; Cysneiros, V.C. Interspecific Interactions of Trees with the Regeneration of Ocotea odorifera (vell.) Rohwer in Mixed Ombrophylous Forest. Floresta 2019, 49, 189. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grouping | Plot Number | Selective Cutting Intensity (%) | Area (m2) | Tree Species Composition | Mean DBH (cm) | Stand Density (Plant/ha) | Stand Volume (m3/ha) |
|---|---|---|---|---|---|---|---|
| Unselected thinning | 21 | 0 | 30 × 70 | Picea asperata Abies fabri Pinus koraiensis Other tree species (group) | 23.5 | 824 | 288.56 |
| 28 | 0 | 40 × 50 | 25 | 725 | 298 | ||
| 32 | 0 | 40 × 40 | 19.9 | 1142 | 261.56 | ||
| 36 | 0 | 40 × 50 | 23.9 | 520 | 178.58 | ||
| 39 | 0 | 40 × 50 | 23.2 | 956 | 298.08 | ||
| Light thinning | 22 | 14.4 | 30 × 70 | Picea asperata Abies fabri Pinus koraiensis Other tree species (group) | 24.2 | 652 | 241.4 |
| 25 | 17.7 | 40 × 50 | 24.2 | 580 | 228.5 | ||
| 30 | 18.1 | 40 × 40 | 25.3 | 638 | 275.62 | ||
| 34 | 21.6 | 40 × 50 | 26.4 | 530 | 247.08 | ||
| 38 | 20 | 40 × 40 | 23.6 | 669 | 224.61 | ||
| Moderate thinning | 24 | 30.8 | 40 × 50 | Picea asperata Abies fabri Pinus koraiensis Other tree species (group) | 25.5 | 519 | 210.83 |
| 27 | 34.3 | 40 × 50 | 25.5 | 620 | 269.91 | ||
| 31 | 31.7 | 30 × 70 | 20.8 | 728 | 235.19 | ||
| 35 | 28.6 | 40 × 50 | 26 | 605 | 262.94 | ||
| 40 | 30.3 | 40 × 50 | 22 | 764 | 343.1 | ||
| Heavy thinning | 23 | 38.1 | 30 × 70 | Picea asperata Abies fabri Pinus koraiensis Other tree species (group) | 24.9 | 581 | 231.24 |
| 26 | 40.9 | 40 × 50 | 26.7 | 480 | 232.65 | ||
| 29 | 37.7 | 30 × 70 | 21.8 | 463 | 144.91 | ||
| 33 | 34.1 | 40 × 50 | 27 | 735 | 352.78 | ||
| 37 | 40.6 | 40 × 50 | 21.2 | 644 | 169.15 |
| Species Name | Abbreviation | Selective Thinning Intensity | Average Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Light Thinning | Moderate Thinning | Heavy Thinning | ||||||||
| IV | BL | IV | BL | IV | BL | IV | BL | IV | BL | ||
| Abies fabri | Af | 28.03 | 4.41 | 29.77 | 4.39 | 31.8 | 4.34 | 31.55 | 4.32 | 30.29 | 4.37 |
| Pinus koraiensis | Pk | 14.85 | 3.92 | 14.59 | 4.77 | 12.97 | 4.06 | 13.07 | 4.35 | 13.87 | 4.28 |
| Picea asperata | Pa | 13.94 | 4.67 | 12.32 | 4.75 | 14.82 | 4.35 | 13.38 | 4.58 | 13.61 | 4.59 |
| Tilia tuan | Tt | 9.05 | 4.35 | 8.84 | 4.05 | 7.22 | 3.62 | 7.52 | 3.55 | 8.16 | 3.89 |
| Betula costata | Bc | 8.06 | 3.47 | 5.24 | 4.43 | 6.35 | 3.12 | 6.75 | 3.49 | 6.6 | 3.63 |
| Larix gmelinii | Lg | 4.1 | 2.74 | 8.45 | 2.73 | 6.17 | 2.19 | 4.63 | 1.45 | 5.83 | 2.28 |
| Acer mono | Am | 5.52 | 3.72 | 5.49 | 2.05 | 3.9 | 2.61 | 6.16 | 2.14 | 5.27 | 2.63 |
| Betula platyphylla | Bp | 5.51 | 4.23 | 5.01 | 2.71 | 4.96 | 3.9 | 5.45 | 1.72 | 5.23 | 3.14 |
| Populus davidiana | Pd | 2.74 | 1.96 | 2.97 | 1.96 | 4.95 | 2.84 | 2.96 | 1.85 | 3.41 | 2.15 |
| Phellodendron amurense | Pe | 3.62 | 2.98 | 2.96 | 1.63 | 2.41 | 2 | 2.54 | 1.98 | 2.88 | 2.15 |
| Ulmus laciniata | Ul | 2.24 | 1.92 | 3.01 | 1.69 | 2.22 | 2 | 3.55 | 2.96 | 2.76 | 2.14 |
| Fraxinus mandschurica | Fm | 2.35 | 2 | 1.35 | 1 | 2.24 | 1.88 | 2.44 | 2 | 2.1 | 1.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.; Wang, X. Niche and Interspecific Association of Dominant Tree Species in Spruce–Fir Mixed Forests in Northeast China. Forests 2023, 14, 1513. https://doi.org/10.3390/f14081513
Yuan S, Wang X. Niche and Interspecific Association of Dominant Tree Species in Spruce–Fir Mixed Forests in Northeast China. Forests. 2023; 14(8):1513. https://doi.org/10.3390/f14081513
Chicago/Turabian StyleYuan, Shuai, and Xinjie Wang. 2023. "Niche and Interspecific Association of Dominant Tree Species in Spruce–Fir Mixed Forests in Northeast China" Forests 14, no. 8: 1513. https://doi.org/10.3390/f14081513
APA StyleYuan, S., & Wang, X. (2023). Niche and Interspecific Association of Dominant Tree Species in Spruce–Fir Mixed Forests in Northeast China. Forests, 14(8), 1513. https://doi.org/10.3390/f14081513