
Evaluating the Impact of Long-Term Land Use Change and Age since Disturbance on Soil Faunal Diversity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
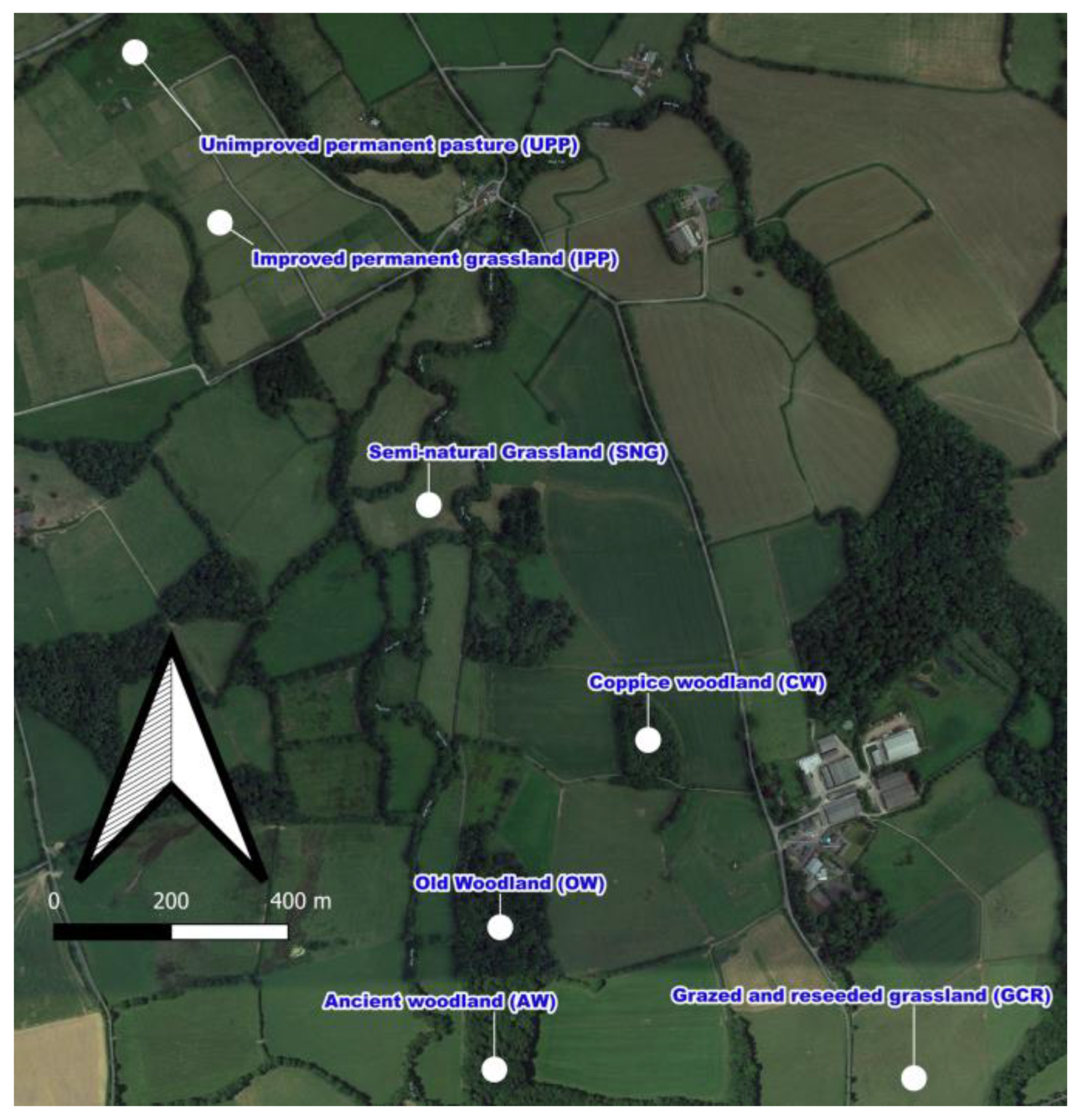
2.1. Site Description
- Ancient woodland (AW), undisturbed and uncultivated soil for more than 200 years, dominated by oak (Quercus robur L.).
- Old woodland (OW), undisturbed and uncultivated soil for less than 200 years old, dominated by ash (Fraxinus spp.).
- Semi-natural grassland (SNG), with no anthropogenic disturbance for more than 50 years, dominated by purple moor grass (Molinia caerulea L.), meadow sweet (Filipendula ulmaria L.) and tussock grass (Deschampsia cespitosa L.), and classified as old culm grassland.
- Coppice woodland (CW) with no disturbance for more than 30 years, dominated by willow (Salix spp.) and poplar (Populus spp.).
- Unimproved permanent pasture (UPP), which received no fertiliser inputs for more than 30 years but was annually grazed by cattle and sheep. The habitat is dominated by creeping bent (Agrostis stolonifera L.), Yorkshire fog (Holcus lanatus L.) and soft rush (Juncus effusus L.).
- Improved permanent grassland (IPP), less than 10 years old, on average 200 kg N ha−1 applied annually and grazed, dominated by perennial ryegrass (Lolium perenne L.) and creeping bent (A. stolonifera).
- Grazed and reseeded grassland (GR), had 40 kg N ha−1 fertilizer per annum and was grazed for 2 years. It was reseeded with high yielding perennial ryegrass (L. perenne cv AberMagic), and white clover (Trifolium repens L. cv AberHerald).
2.2. Arthropod Sampling and Extraction
2.3. Analysis of Chemical Properties of Soil and Plants
2.4. Statistical Analysis
3. Results
3.1. Soil and Litter Properties
3.2. Abundance of Invertebrates
3.3. Diversity of Invertebrate Communities
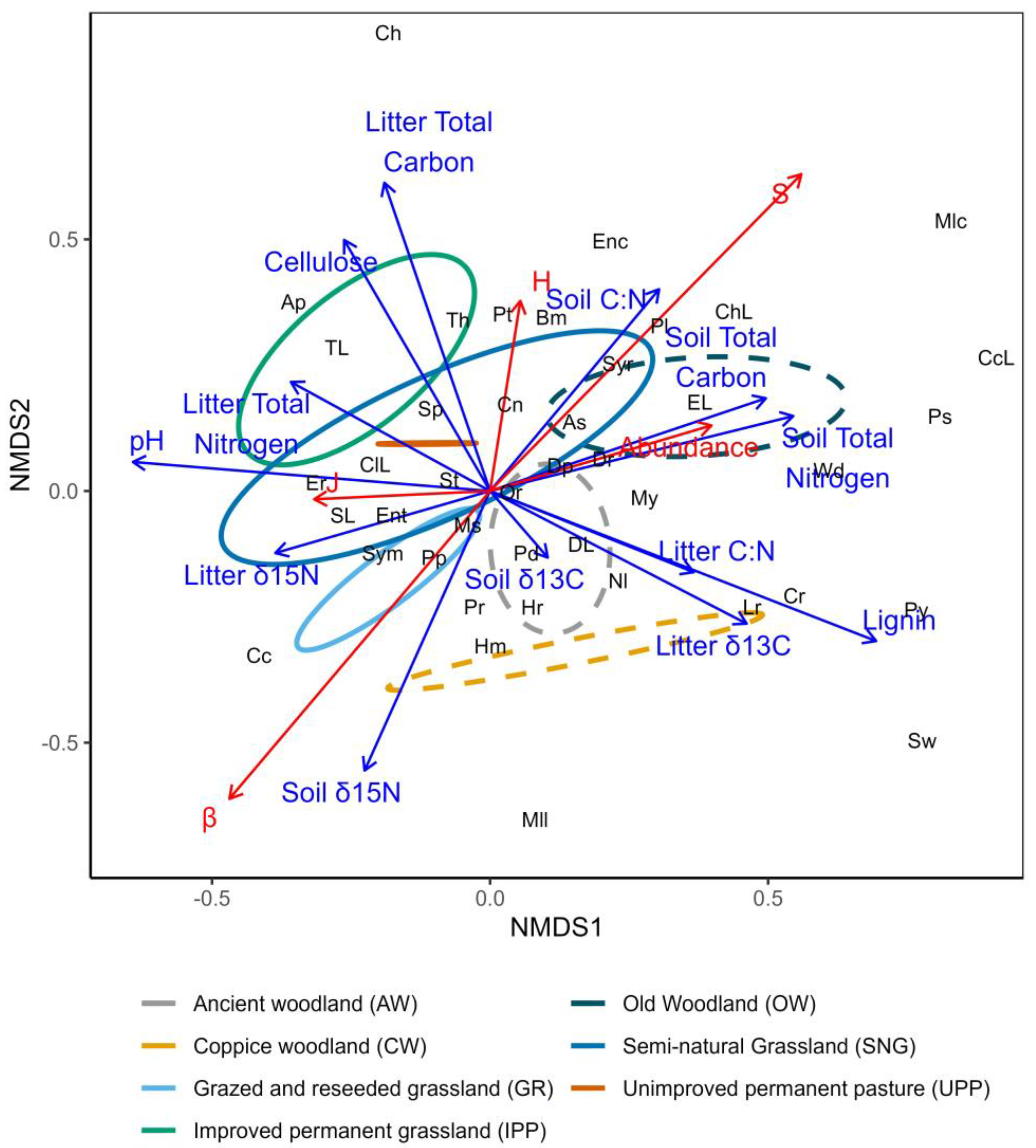
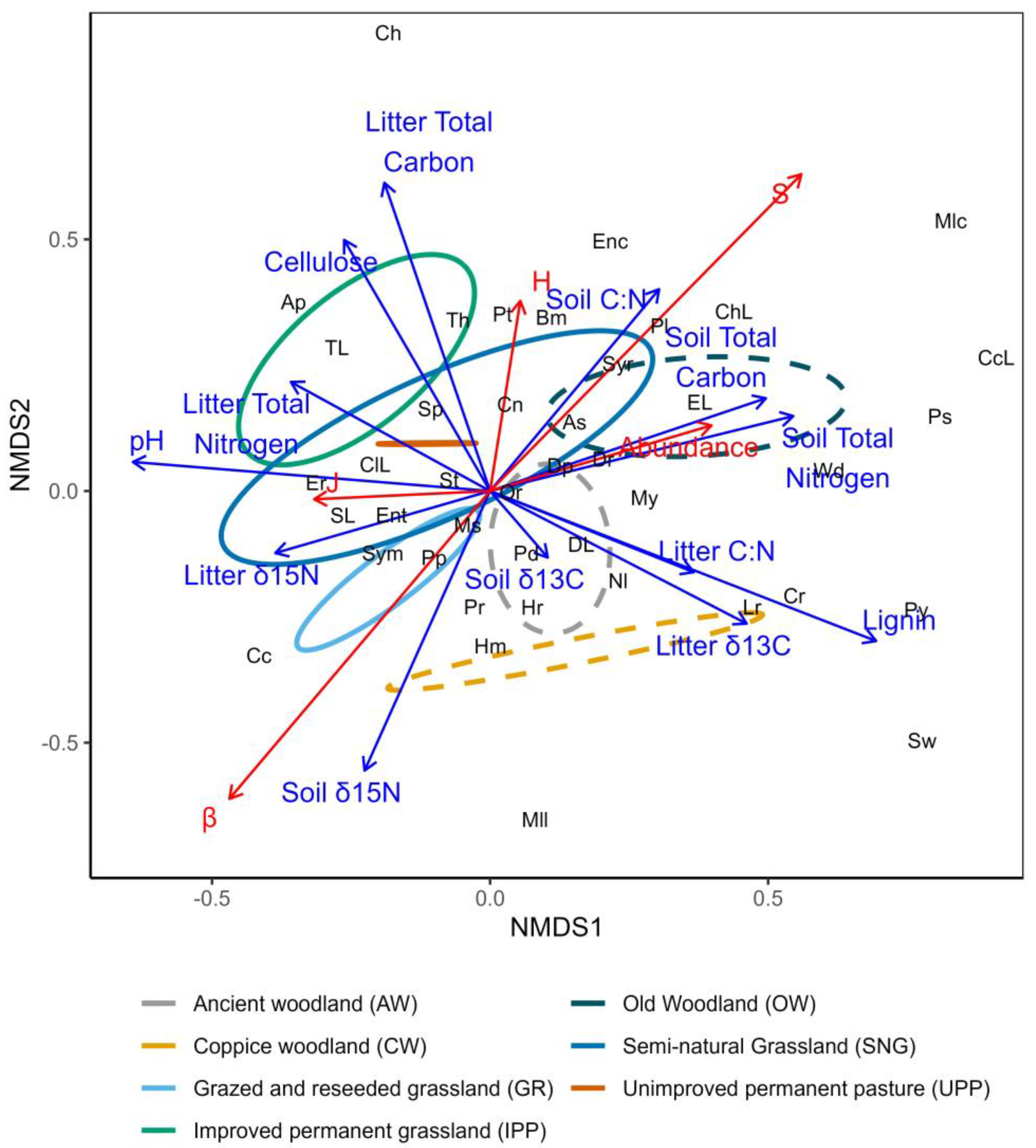
3.4. Multivariate Community Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bardgett, R.; Bowman, W.; Kaufmann, R.; Schmidt, S. A Temporal Approach to Linking Aboveground and Belowground Ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef] [PubMed]
- de Wit, R.; Bouvier, T. “Everything Is Everywhere, but, the Environment Selects”; What Did Baas Becking and Beijerinck Really Say? Environ. Microbiol. 2006, 8, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Maraun, M. The Structure of Oribatid Mite Communities (Acari, Oribatida): Patterns, Mechanisms and Implications for Future Research. Ecography 2000, 23, 374–383. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Certini, G. Effects of Fire on Properties of Forest Soils: A Review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Scheu, S.; Schulz, E. Secondary Succession, Soil Formation and Development of a Diverse Community of Oribatids and Saprophagous Soil Macro-Invertebrates. Biodivers. Conserv. 1996, 5, 235–250. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef]
- Ashford, O.S.; Foster, W.A.; Turner, B.L.; Sayer, E.J.; Sutcliffe, L.; Tanner, E.V. Litter Manipulation and the Soil Arthropod Community in a Lowland Tropical Rainforest. Soil Biol. Biochem. 2013, 62, 5–12. [Google Scholar] [CrossRef]
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L.; et al. Perennial Grasslands Enhance Biodiversity and Multiple Ecosystem Services in Bioenergy Landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef]
- Rhrig, R.; Langmaack, M.; Schrader, S.; Larink, O. Tillage Systems and Soil Compaction Their Impact on Abundance and Vertical Distribution of Enchytraeidae. Soil Tillage Res. 1998, 46, 117–127. [Google Scholar] [CrossRef]
- Moore, J.C.; Berlow, E.L.; Coleman, D.C.; Ruiter, P.C.; Dong, Q.; Hastings, A.; Johnson, N.C.; McCann, K.S.; Melville, K.; Morin, P.J.; et al. Detritus, Trophic Dynamics and Biodiversity: Detritus, Trophic Dynamics and Biodiversity. Ecol. Lett. 2004, 7, 584–600. [Google Scholar] [CrossRef]
- Hairiah, K.; Sitompul, S.M.; Van Noordwijk, M.; Palm, C. Methods for Sampling Carbon Stocks above and below Ground; ICRAF: Bogor, Indonesia, 2001. [Google Scholar]
- Rantalainen, M.-L.; Haimi, J.; Fritze, H.; Setälä, H. Effects of Small-Scale Habitat Fragmentation, Habitat Corridors and Mainland Dispersal on Soil Decomposer Organisms. Appl. Soil Ecol. 2006, 34, 152–159. [Google Scholar] [CrossRef]
- Barrios, E. Soil Biota, Ecosystem Services and Land Productivity. Ecol. Econ. 2007, 64, 269–285. [Google Scholar] [CrossRef]
- Bedano, J.C.; Cantú, M.P.; Doucet, M.E. Influence of Three Different Land Management Practices on Soil Mite (Arachnida: Acari) Densities in Relation to a Natural Soil. Appl. Soil Ecol. 2006, 32, 293–304. [Google Scholar] [CrossRef]
- Culliney, T.W. Role of Arthropods in Maintaining Soil Fertility. Agriculture 2013, 3, 629–659. [Google Scholar] [CrossRef]
- Caruso, T.; Taormina, M.; Migliorini, M. Relative Role of Deterministic and Stochastic Determinants of Soil Animal Community: A Spatially Explicit Analysis of Oribatid Mites: Deterministic and Stochastic Determinants of Soil Animal Community. J. Anim. Ecol. 2012, 81, 214–221. [Google Scholar] [CrossRef]
- Gulvik, M.E. Mites [Acari] as Indicators of Soil Biodiversity and Land Use Monitoring: A Review. Pol. J. Ecol. 2007, 3, 415–440. [Google Scholar]
- Buchs, W.; Raubuch, M.; Prescher, S.; Behr, K.; Muller, A.; Roose, K. Impact of Ostrinia-Resistant Bt-Maize on Microbial and Invertebrate Decomposer Communities in Field Soils. Mitteilungen-Biol. Bundesanst. Fur Land Und Forstwirtsch. 2007, 410, 26. [Google Scholar]
- Breure, A.M.; Mulder, C.; Römbke, J.; Ruf, A. Ecological Classification and Assessment Concepts in Soil Protection. Ecotoxicol. Environ. Saf. 2005, 62, 211–229. [Google Scholar] [CrossRef]
- Buckland, S.T.; Magurran, A.E.; Green, R.E.; Fewster, R.M. Monitoring Change in Biodiversity through Composite Indices. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 243–254. [Google Scholar] [CrossRef]
- George, P.B.L.; Keith, A.M.; Creer, S.; Barrett, G.L.; Lebron, I.; Emmett, B.A.; Robinson, D.A.; Jones, D.L. Evaluation of mesofauna communities as soil quality indicators in a national-level monitoring programme. Soil Biol. Biochem. 2017, 115, 537–546. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; de Ruiter, P.C.; van der Putten, W.H.; Birkhofer, K.; Hemerik, L.; de Vries, F.T.; Bardgett, R.D.; Brady, M.V.; et al. Intensive Agriculture Reduces Soil Biodiversity across Europe. Glob. Chang. Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Crotty, F.V.; Adl, S.M.; Blackshaw, R.P.; Murray, P.J. Protozoan Pulses Unveil Their Pivotal Position within the Soil Food Web. Microb. Ecol. 2012, 63, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Crotty, F.V.; Blackshaw, R.P.; Adl, S.M.; Inger, R.; Murray, P.J. Divergence of Feeding Channels within the Soil Food Web Determined by Ecosystem Type. Ecol. Evol. 2014, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.L.; Blackshaw, R.P.; Dunn, R.M.; Critchley, N.R.; Smith, K.E.; Williams, J.R.; Randall, N.P.; Murray, P.J. Improving above and Below-Ground Arthropod Biodiversity in Maize Cultivation Systems. Appl. Soil Ecol. 2016, 108, 25–46. [Google Scholar] [CrossRef]
- Norris, S.L. The Effect of Maize Cultivation on Invertebrate Biodiversity. Ph.D. Thesis, Harper Adams University, Newport, UK, 2015. [Google Scholar]
- Werner, M.R.; Dindal, D.L. Effects of Conversion to Organic Agricultural Practices on Soil Biota. Am. J. Altern. Agric. 1990, 5, 24–32. [Google Scholar] [CrossRef]
- Hopkin, S.P. A Key to the Collembola (Springtails) of Britain and Ireland, 1st ed.; Occasional Publication/Field Studies Council; Field Studies Council: Preston Montford, UK, 2007; ISBN 978-1-85153-220-9. [Google Scholar]
- Krantz, G.W.; Walter, D.E. (Eds.) A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; ISBN 978-0-89672-620-8. [Google Scholar]
- Tilling, S.M. A Key to the Major Groups of British Terrestrial Invertebrates; Field Studies Council: Shrewsbury, UK, 1987; ISBN 978-1-85153-187-5. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Kitcherside, M.A.; Glen, E.F.; Webster, A.J.F. FibreCap: An Improved Method for the Rapid Analysis of Fibre in Feeding Stuffs. Anim. Feed. Sci. Technol. 2000, 86, 125–132. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Racine, J.S. RStudio: A Platform-Independent IDE for R and Sweave: Software Review. J. Appl. Econ. 2012, 27, 167–172. [Google Scholar] [CrossRef]
- Gu, C. Smoothing Spline ANOVA Models: R Package Gss. J. Stat. Softw. 2014, 58, 1–25. [Google Scholar] [CrossRef]
- De Mendiburu, F.; Simon, R. Agricolae—Ten Years of an Open Source Statistical Tool for Experiments in Breeding, Agriculture and Biology. PeerJ PrePrints 2015, 3, e1404v1. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2018, 1. 15-1; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring Beta Diversity for Presence-Absence Data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef]
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. R Package Version 2.0-1; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. JOSS 2019, 4, 1686. [Google Scholar] [CrossRef]
- Culman, S.W.; Young-Mathews, A.; Hollander, A.D.; Ferris, H.; Sánchez-Moreno, S.; O’Geen, A.T.; Jackson, L.E. Biodiversity Is Associated with Indicators of Soil Ecosystem Functions over a Landscape Gradient of Agricultural Intensification. Landsc. Ecol. 2010, 25, 1333–1348. [Google Scholar] [CrossRef]
- Ramezani, L.; Mossadegh, M. Biodiversity of Oribatid Mites in Two Different Microhabitats of Khuzestan Province (Southwestern Iran). J. Crop Prot. 2014, 3, 443–448. [Google Scholar]
- Osler, G.H.R.; Sommerkorn, M. Toward a Complete Soil C and N Cycle: Incorporating the Soil Fauna. Ecology 2007, 88, 1611–1621. [Google Scholar] [CrossRef]
- Halaj, J.; Peck, R.W.; Niwa, C.G. Trophic Structure of a Macroarthropod Litter Food Web in Managed Coniferous Forest Stands: A Stable Isotope Analysis with Δ15N and Δ13C. Pedobiologia 2005, 49, 109–118. [Google Scholar] [CrossRef]
- Neher, D.A.; Barbercheck, M.E.; El-Allaf, S.M.; Anas, O. Effects of Disturbance and Ecosystem on Decomposition. Appl. Soil Ecol. 2003, 23, 165–179. [Google Scholar] [CrossRef]
- Balesdent, J.; Mariotti, A.; Boisgontier, D. Effect of Tillage on Soil Organic Carbon Mineralization Estimated from 13C Abundance in Maize Fields. J. Soil Sci. 1990, 41, 587–596. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Yuyukina, T.; Blagodatsky, S.; Kuzyakov, Y. Turnover of Soil Organic Matter and of Microbial Biomass under C3–C4 Vegetation Change: Consideration of 13C Fractionation and Preferential Substrate Utilization. Soil Biol. Biochem. 2011, 43, 159–166. [Google Scholar] [CrossRef]
- Loranger, G.; Bandyopadhyaya, I.; Razaka, B.; Ponge, J.-F. Does Soil Acidity Explain Altitudinal Sequences in Collembolan Communities? Soil Biol. Biochem. 2001, 33, 381–393. [Google Scholar] [CrossRef]
- Hines, J.; van der Putten, W.H.; De Deyn, G.B.; Wagg, C.; Voigt, W.; Mulder, C.; Weisser, W.W.; Engel, J.; Melian, C.; Scheu, S.; et al. Towards an Integration of Biodiversity–Ecosystem Functioning and Food Web Theory to Evaluate Relationships between Multiple Ecosystem Services. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 53, pp. 161–199. ISBN 978-0-12-803885-7. [Google Scholar]

{kind=link}
{kind=link}
| AW | OW | CW | SNG | UPP | IPP | GR | |
|---|---|---|---|---|---|---|---|
| pH | 4.18 (±0.08) bc | 3.82 (±0.16) c | 4.71 (±0.14) b | 4.64 (±0.12) b | 5.66 (±0.11) a | 5.36 (±0.09) a | 5.46 (±0.07) a |
| Lignin (%) | 28.31 (±2.41) a | 32.8 (±2.74) a | 26.1 (±2.84) a | 3.85 (±0.47) b | 1.2 (±0.65) bc | 1.03 (±0.13) bc | 0.64 (±0.27) c |
| Cellulose (%) | 19.3 (±1.97) ab | 17 (±3.45) ab | 13.73 (±1.07) b | 26.1 (±4.5) a | 21.96 (±0.52) ab | 21.68 (±0.67) ab | 21.49 (±1.08) ab |
| Litter TN (%) | 1.42 (±0.05) ab | 1.75 (±0.03) bc | 1.09 (±0.1) a | 1.76 (±0.18) bc | 2.62 (±0.12) c | 1.44 (±0.09) ab | 2.36 (±0.34) c |
| Litter TC (%) | 40.89 (±1.62) a | 43.07 (±0.73) a | 33.87 (±2.02) b | 40.05 (±0.74) a | 43.28 (±0.2) a | 41.91 (±0.23) a | 42.78 (±0.17) a |
| Litter C:N | 28.84 (±1.04) a | 24.56 (±0.37) ab | 31.53 (±2.02) a | 23.23 (±2.51) ab | 16.57 (±0.74) b | 29.4 (±2.08) a | 18.79 (±2.34) b |
| Litter δ13C | −29.2 (±0.33) a | −29.68 (±0.03) ab | −29.24 (±0.1) a | −29.61 (±0.31) ab | −31.14 (±0.36) c | −30.65 (±0.05) bc | −31.11 (±0.1) c |
| Litter δ15N | −2.03 (±0.26) c | −0.73 (±0.13) bc | 1.26 (±0.09) ab | 1.44 (±0.37) ab | 4.96 (±0.14) a | 0.15 (±0.82) bc | 2.97 (±1.91) ab |
| Soil TN (%) | 0.91 (±0.15) de | 0.95 (±0.01) e | 0.68 (±0) cd | 0.48 (±0) b | 0.69 (±0.01) cd | 0.61 (±0) c | 0.38 (±0) a |
| Soil TC (%) | 14.24 (±3.59) d | 13.73 (±0.09) d | 6.62 (±0.08) bc | 5.8 (±0.04) b | 7.56 (±0.06) c | 6.48 (±0.11) bc | 3.49 (±0.05) a |
| Soil C:N | 0.91 (±0.15) c | 24.42 (±4.46) a | 9.72 (±0.06) b | 12.08 (±0.04) b | 10.98 (±0.03) b | 10.67 (±0.08) b | 9.1 (±0.06) b |
| Soil δ13C | −28.09 (±0.08) e | −28.68 (±0.02) c | −29.33 (±0.03) b | −28.2 (±0.03) e | −28.73 (±0.03) c | −29.53 (±0.04) a | −28.46 (±0) d |
| Soil δ15N | 1.48 (±0.11) f | 2.1 (±0.06) e | 5.97 (±0.17) a | 4.54 (±0.2) b | 3.63 (±0.07) c | 2.83 (±0.15) d | 5.46 (±0.09) a |
| Species | AW | OW | CW | SNG | UPP | IPP | GR |
|---|---|---|---|---|---|---|---|
| Oribatida | 17,200 (±6042) | 15,281 (±6762) | 25,204 (±15,283) | 18,324 (±7296) | 7608 (±6426) | 25,072 (±13,790) | 265 (±132) |
| Mesostigmata | 3771 (±1801) | 2382 (±1301) | 3969 (±917) | 3175 (±1291) | 5292 (±1622) | 9328 (±2529) | 1588 (±992) |
| Prostigmata | 3175 (±992) | 2051 (±350) | 2646 (±1605) | 992 (±525) | 794 (±794) | 3374 (±865) | 8203 (±4532) |
| Astigmata | 265 (±175) | 6748 (±6157) | 198 (±198) | 2117 (±1720) | 66 (±66) | 926 (±288) | 198 (±115) |
| Entomobryomorpha | 2911 (±2327) | 3043 (±1236) | 7740 (±1988) | 1985 (±716) | 8269 (±1670) | 14,223 (±9964) | 3175 (±2091) |
| Symphypleona | 662 (±175) | 132 (±66) | 1588 (±525) | 926 (±652) | 5028 (±434) | 595 (±303) | 3175 (±1260) |
| Poduromorpha | 24,411 (±14,853) | 7145 (±2560) | 5094 (±529) | 2514 (±691) | 2382 (±1093) | 2183 (±115) | 1058 (±565) |
| Neelipleona | 1323 (±763) | 1786 (±751) | 2249 (±1278) | 265 (±175) | - | 926 (±926) | - |
| Staphylinidae | 132 (±66) | 66 (±66) | - | 265 (±175) | 397 (±229) | 397 (±115) | 132 (±66) |
| Diptera | 66 (±66) | 66 (±66) | - | - | - | 66 (±66) | - |
| Hemiptera | 463 (±463) | 66 (±66) | 66 (±66) | - | - | - | - |
| Curculionidae | 66 (±66) | 132 (±66) | 66 (±66) | - | - | - | - |
| Thysanoptera | 66 (±66) | 265 (±66) | - | 1588 (±1050) | 1323 (±288) | 1389 (±303) | - |
| Harpacticoida | 66 (±66) | - | - | - | - | - | - |
| Deroceras (Mollusca) | 66 (±66) | - | - | - | - | - | - |
| Mycetophagidae | 66 (±66) | - | - | - | - | - | - |
| Syrphidae | - | 66 (±66) | - | - | - | - | - |
| Bembidion | - | 66 (±66) | - | 265 (±175) | - | - | - |
| Polyxenidae | - | 66 (±66) | - | - | - | - | - |
| Ptiliidae | - | 198 (±115) | - | 66 (±66) | 66 (±66) | 198 (±115) | - |
| Malachiidae | - | 66 (±66) | - | - | - | - | - |
| Coccinellidae | - | - | 66 (±66) | 66 (±66) | - | - | - |
| Symphyta | - | - | 66 (±66) | - | - | - | - |
| Aphididae | - | - | - | - | 728 (±288) | 992 (±895) | - |
| Chalcidoidea | - | - | - | - | 66 (±66) | - | - |
| Pseudoscorpionida | - | 265 (±265) | 66 (±66) | - | - | - | - |
| Araneae | - | 198 (±198) | 66 (±66) | - | 529 (±434) | 397 (±229) | - |
| Armadillidium | - | 463 (±66) | 198 (±198) | - | - | - | - |
| Myriapoda | - | 132 (±132) | - | 66 (±66) | - | - | 66 (±66) |
| Diplopoda | - | - | 132 (±66) | - | - | - | - |
| Lumbricus | 66 (±66) | - | - | 132 (±66) | - | 198 (±115) | 66 (±66) |
| Enchytraeidae | - | 728 (±288) | - | - | 463 (±463) | - | - |
| Staphylinidae larvae | 198 (±115) | 198 (±115) | 198 (±198) | 198 (±115) | 662 (±565) | 1125 (±175) | 331 (±132) |
| Diptera larvae | 397 (±229) | 331 (±239) | 331 (±175) | - | 132 (±66) | - | 66 (±66) |
| Elateridae larvae | 66 (±66) | 132 (±66) | - | - | - | 66 (±66) | - |
| Coleoptera larvae | - | - | 265 (±175) | 265 (±175) | 132 (±66) | 198 (±115) | - |
| Tipulidae larvae | - | 198 (±198) | - | 132 (±66) | 132 (±66) | - | - |
| Coccinella larvae | - | 66 (±66) | - | - | - | - | - |
| Chironomidae larvae | - | 198 (±198) | - | 132 (±132) | - | - | - |
| Larvae | - | 265 (±265) | 66 (±66) | - | - | - | - |
| Pupa | - | - | 132 (±132) | - | 66 (±66) | - | - |
| AW | OW | CW | SNG | UPP | IPP | GR | |
|---|---|---|---|---|---|---|---|
| Richness | 12.67 (±1.86) ab | 18.67 (±0.33) a | 12.67 (±0.88) ab | 13.00 (±2.00) ab | 13.00 (±1.00) ab | 14.33 (±0.33) ab | 9.00 (±1.00) b |
| Total Abundance m−2 × 104 | 5.53 (±2.57) a | 4.27 (±1.45) a | 5.03 (±1.89) a | 3.34 (±0.96) a | 3.41 (±1.08) a | 6.15 (±1.59) a | 1.83 (±0.77) a |
| Shannon Diversity | 1.54 (±0.08) a | 2.00 (±0.14) a | 1.68 (±0.16) a | 1.62 (±0.19) a | 1.87 (±0.14) a | 1.71 (±0.23) a | 1.55 (±0.10) a |
| Evenness (J) | 0.62 (±0.06) a | 0.69 (±0.05) a | 0.66 (±0.07) a | 0.64 (±0.07) a | 0.73 (±0.03) a | 0.64 (±0.09) a | 0.71 (±0.05) a |
| β-diversity | 2.49 (±0.59) ab | 1.25 (±0.04) b | 2.35 (±0.24) ab | 2.37 (±0.45) ab | 2.27 (±0.27) ab | 1.93 (±0.07) ab | 3.80 (±0.60) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crotty, F.V.; Demirer, U.A.; Norris, S.L.; Liu, W.; Murray, P.J. Evaluating the Impact of Long-Term Land Use Change and Age since Disturbance on Soil Faunal Diversity. Forests 2023, 14, 1882. https://doi.org/10.3390/f14091882
Crotty FV, Demirer UA, Norris SL, Liu W, Murray PJ. Evaluating the Impact of Long-Term Land Use Change and Age since Disturbance on Soil Faunal Diversity. Forests. 2023; 14(9):1882. https://doi.org/10.3390/f14091882
Chicago/Turabian StyleCrotty, Felicity Victoria, Umran Akkan Demirer, Stuart Lee Norris, Wei Liu, and Philip James Murray. 2023. "Evaluating the Impact of Long-Term Land Use Change and Age since Disturbance on Soil Faunal Diversity" Forests 14, no. 9: 1882. https://doi.org/10.3390/f14091882
APA StyleCrotty, F. V., Demirer, U. A., Norris, S. L., Liu, W., & Murray, P. J. (2023). Evaluating the Impact of Long-Term Land Use Change and Age since Disturbance on Soil Faunal Diversity. Forests, 14(9), 1882. https://doi.org/10.3390/f14091882