
Physiological Responses Revealed Static Magnetic Fields Potentially Improving the Tolerance of Poplar Seedlings to Salt Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Static MF Setup
2.2. Plant Materials and Experimental Conditions
2.3. Measurement of ROS and [Ca2+]cyt
2.4. Intracellular Ion Concentrations
2.5. Root Surface Net Ion Flux
2.5.1. Steady-State Measurements of Net Na+, H+, K+ and Ca2+ Fluxes
2.5.2. Ion Channel Inhibitor Treatment
2.5.3. Transient Ion Flux Kinetics
2.6. qRT-PCR Analyses
2.7. Data Analysis
3. Results
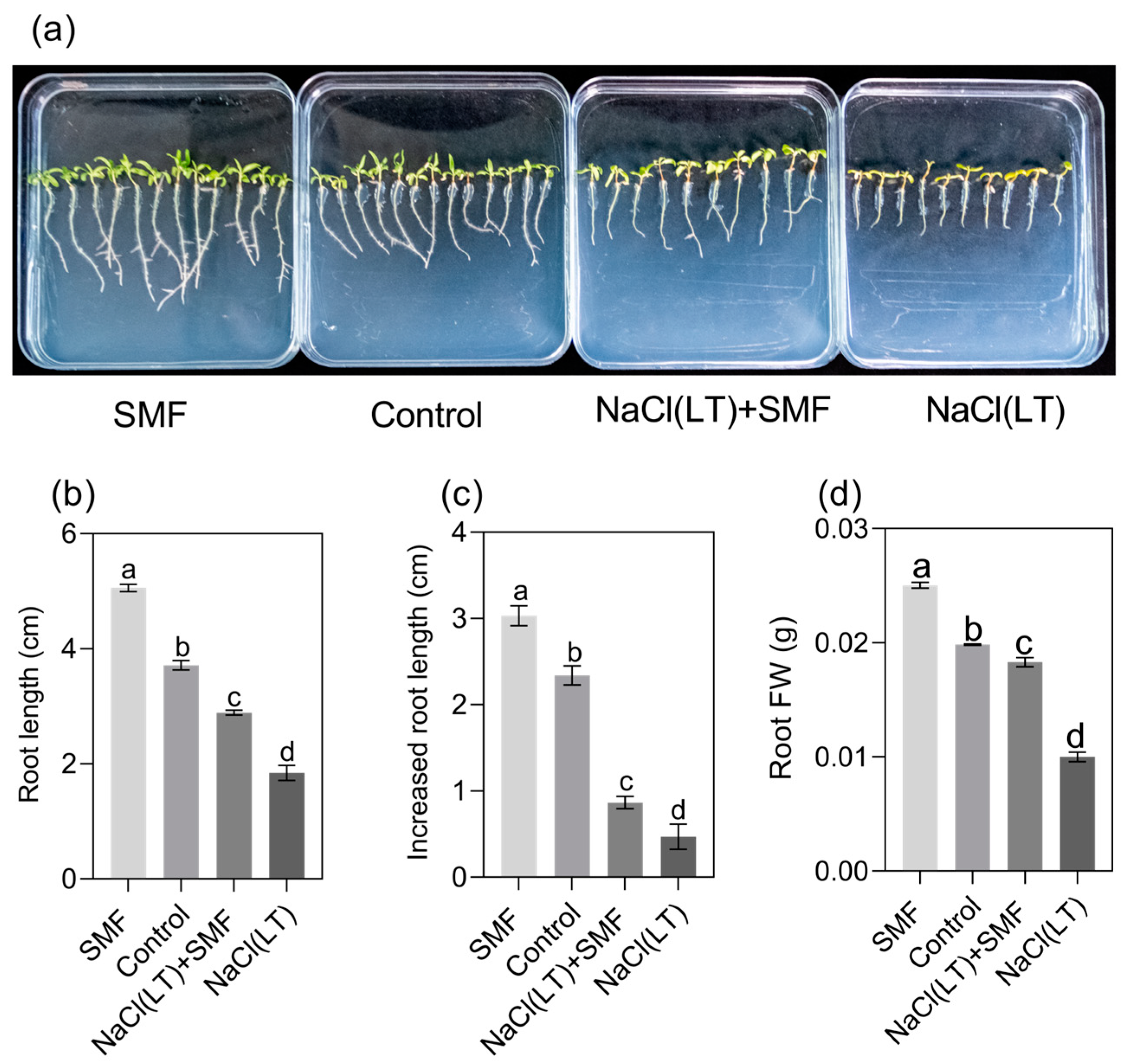
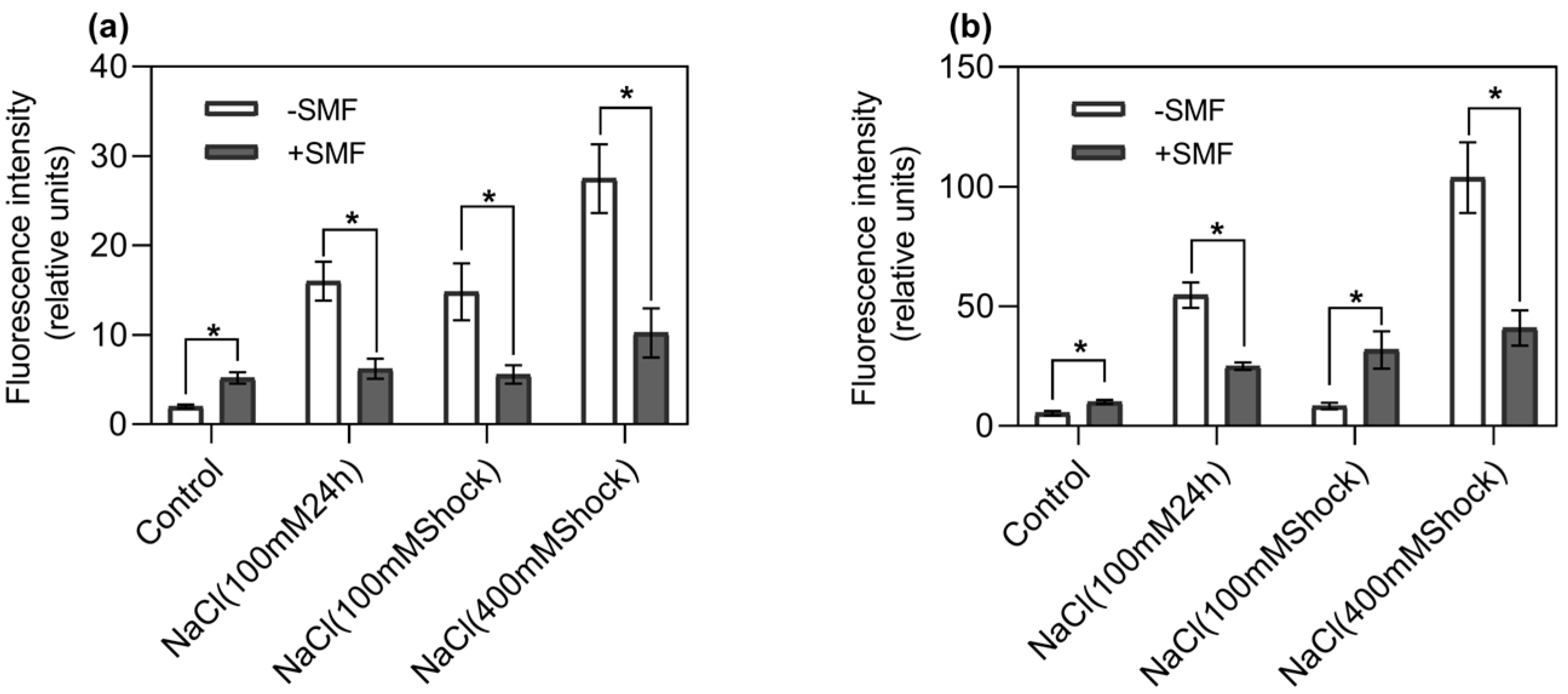
3.1. Poplar Seedling Growth and ROS and [Ca2+]cyt Levels
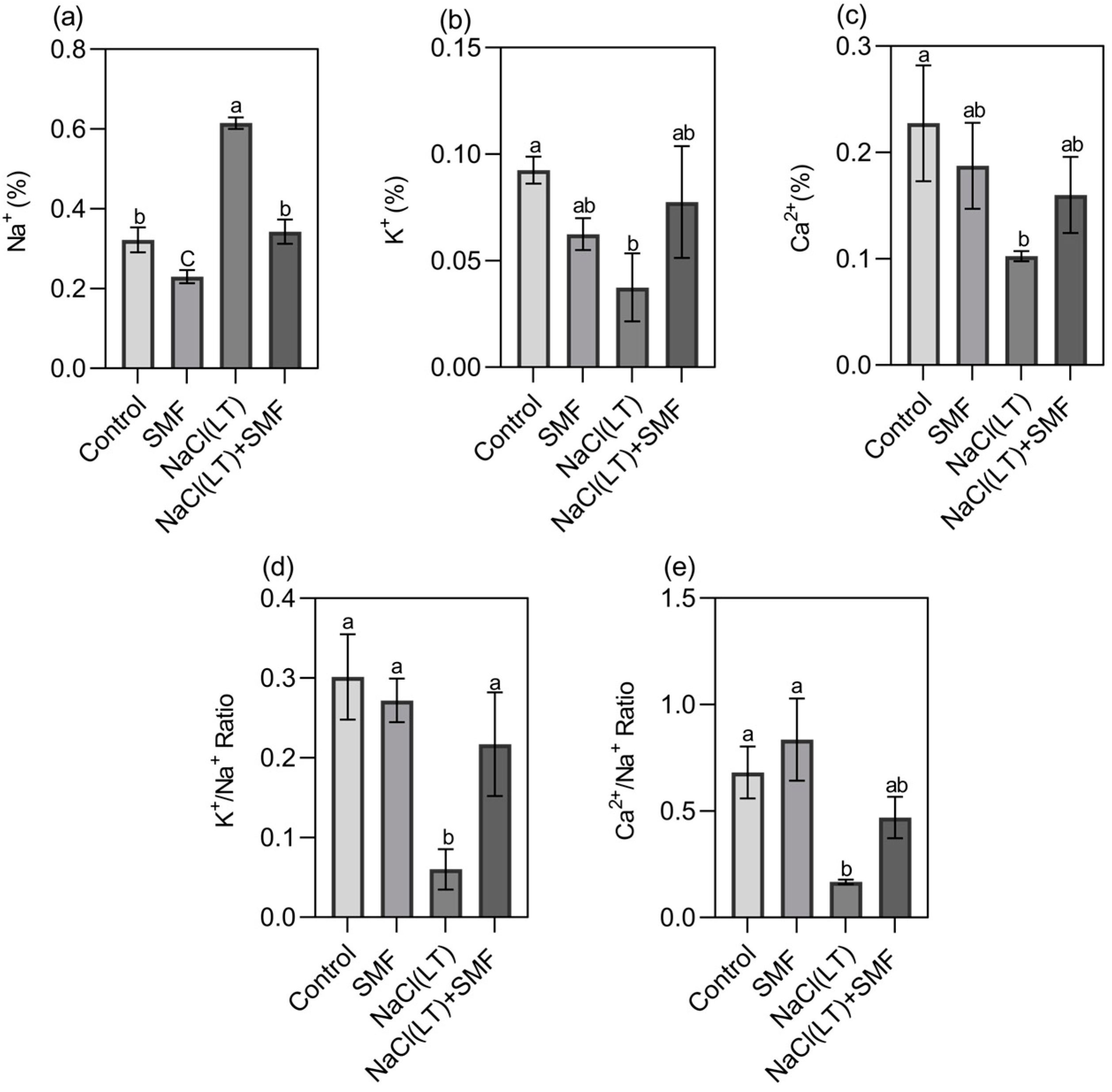
3.2. Concentrations of Na+, K+ and Ca2+ in Roots
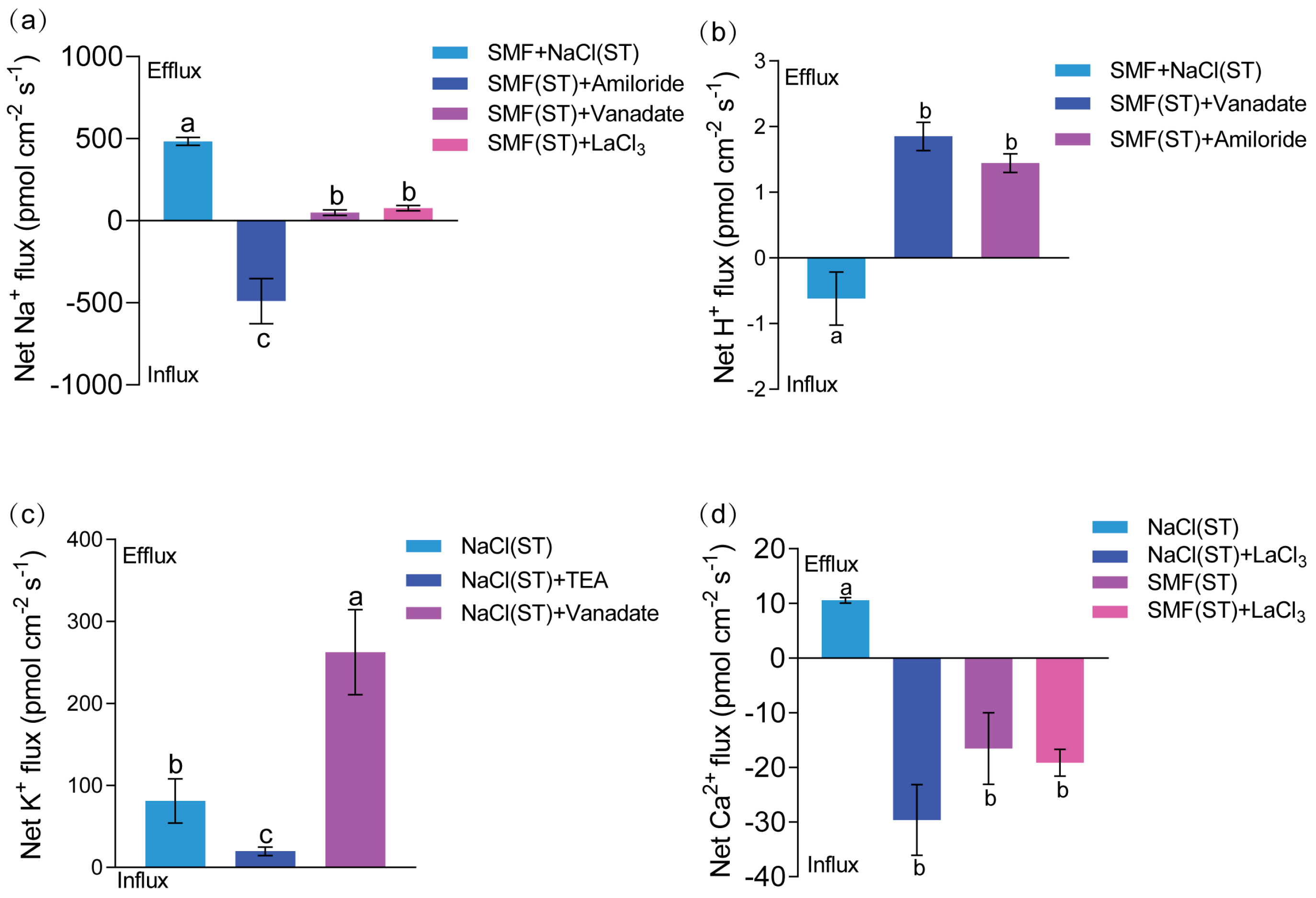
3.3. Root Surface Ion Flux Dynamics
3.3.1. Na+ Flux
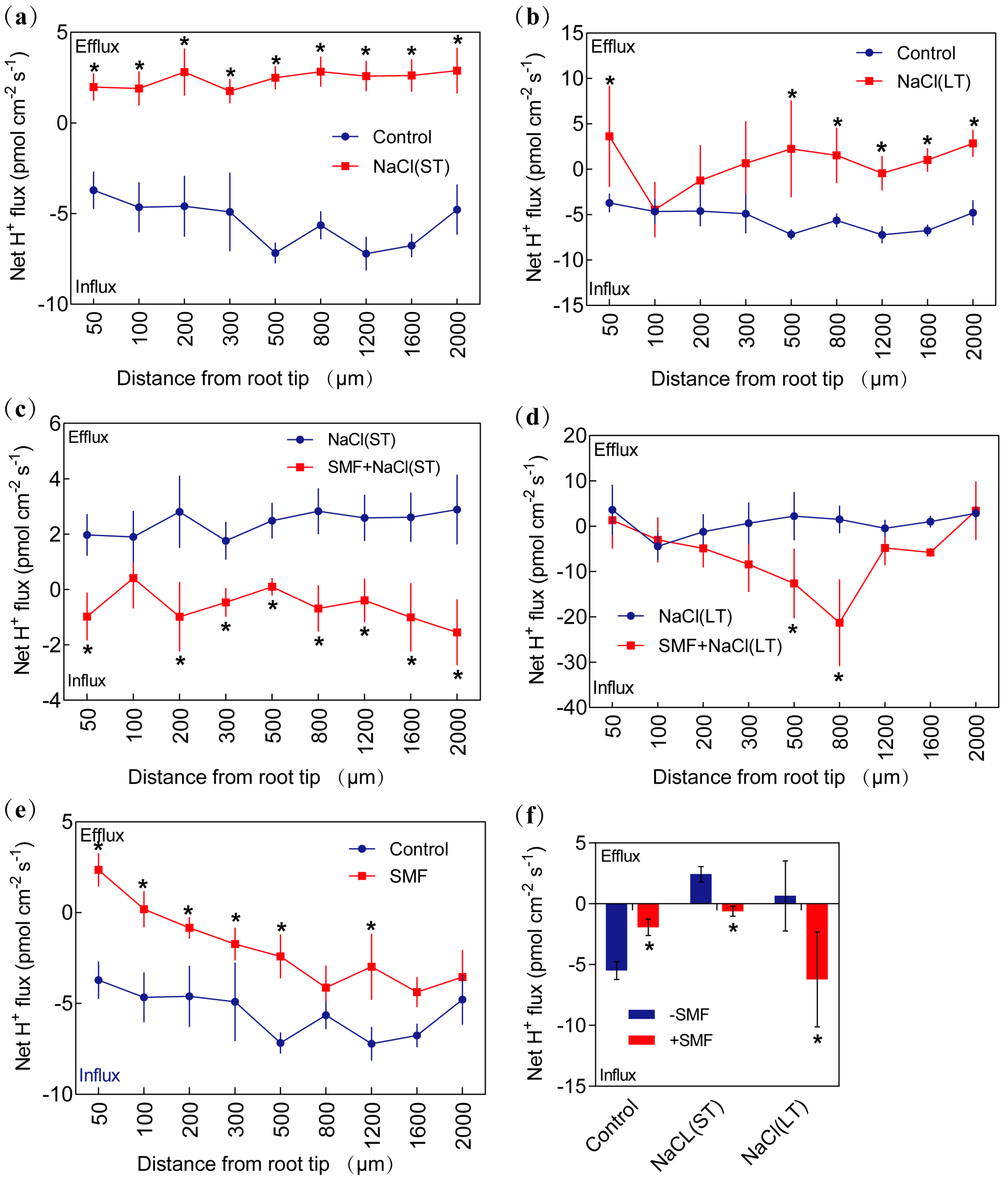
3.3.2. H+ Flux
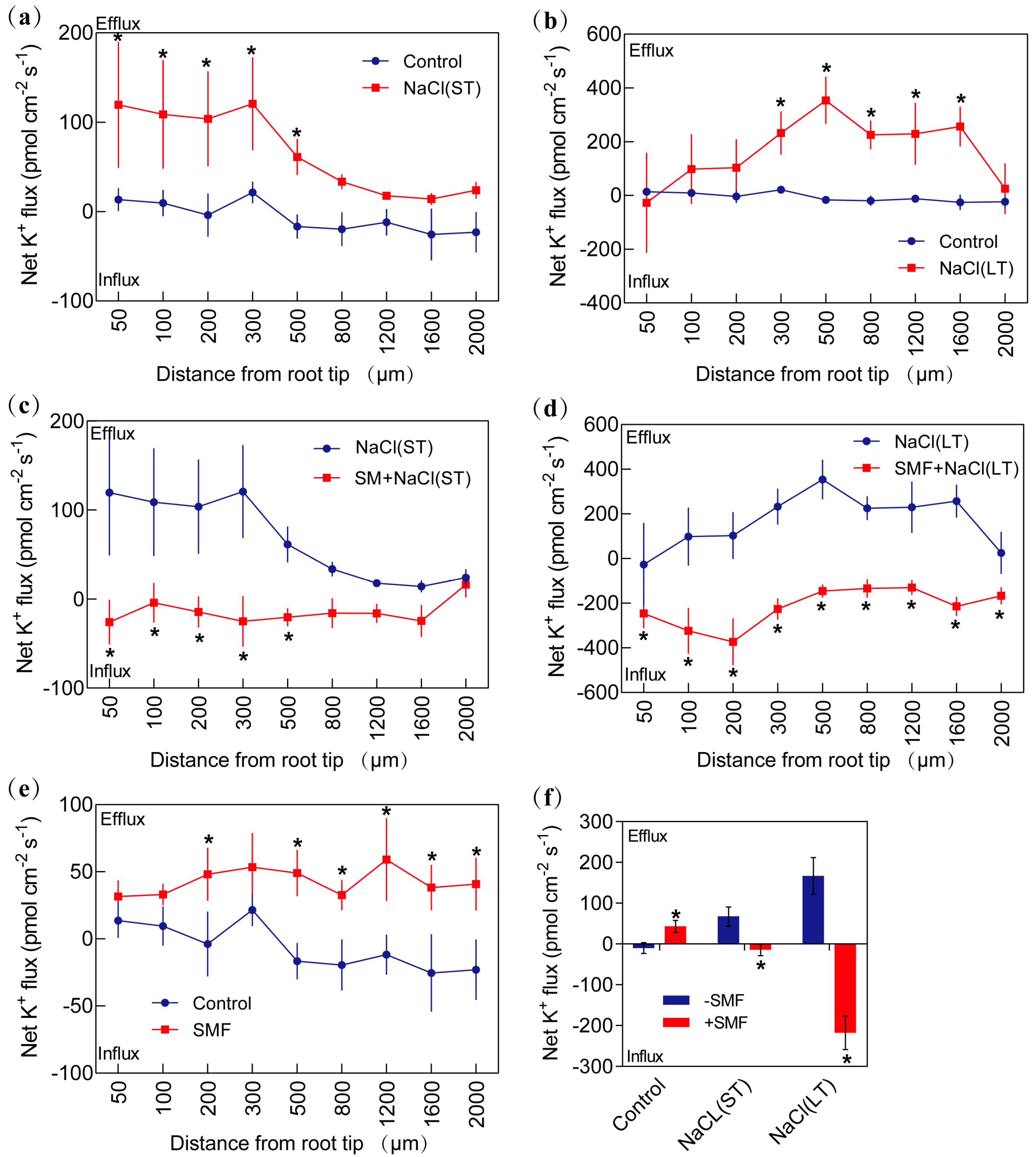
3.3.3. K+ Flux
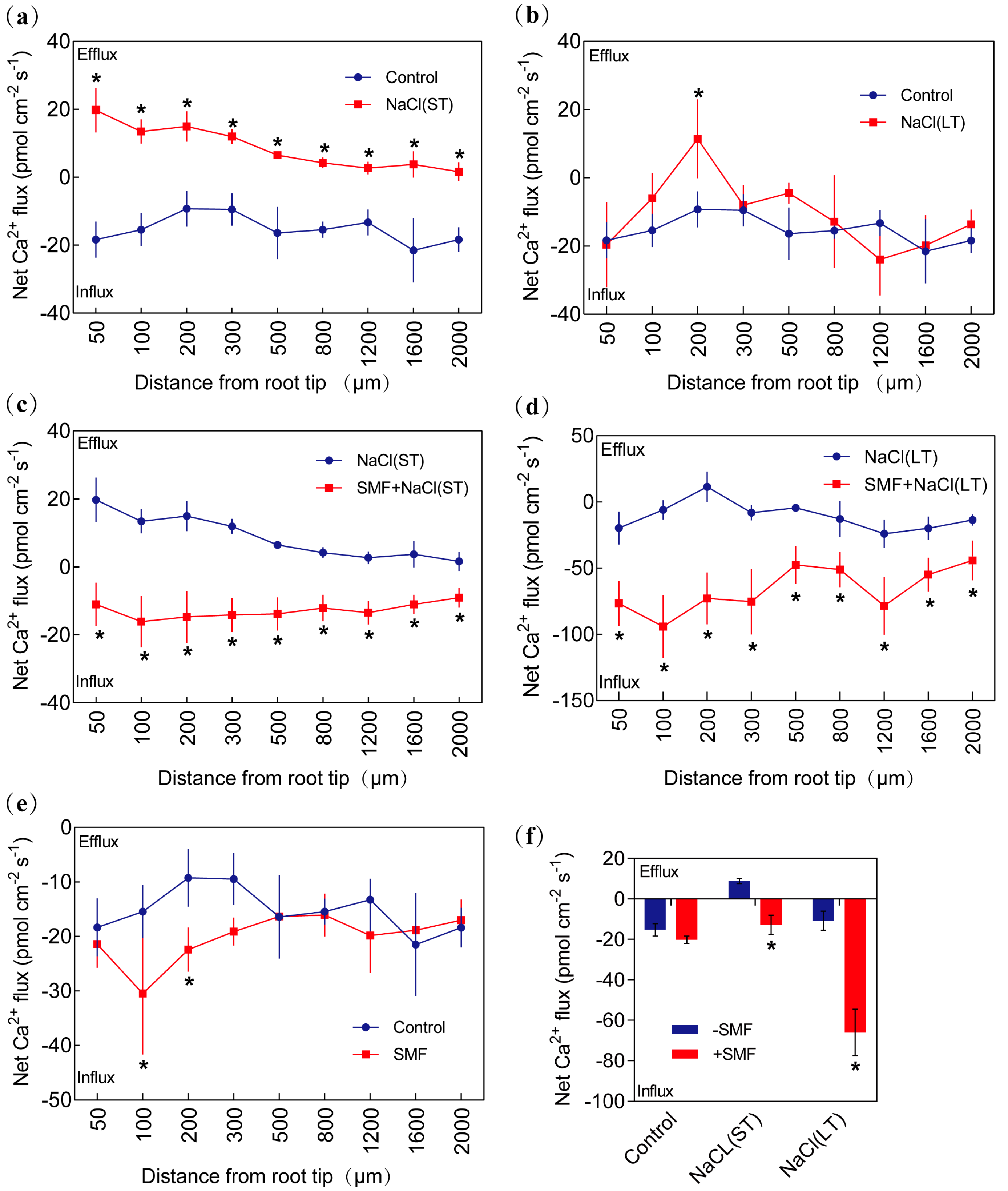
3.3.4. Ca2+ Flux
3.4. Expression of Stress-Responsive ROS Production and Ion Transporter Genes
4. Discussion
4.1. Seedling Physiological Responses to SMF and Salt Stress
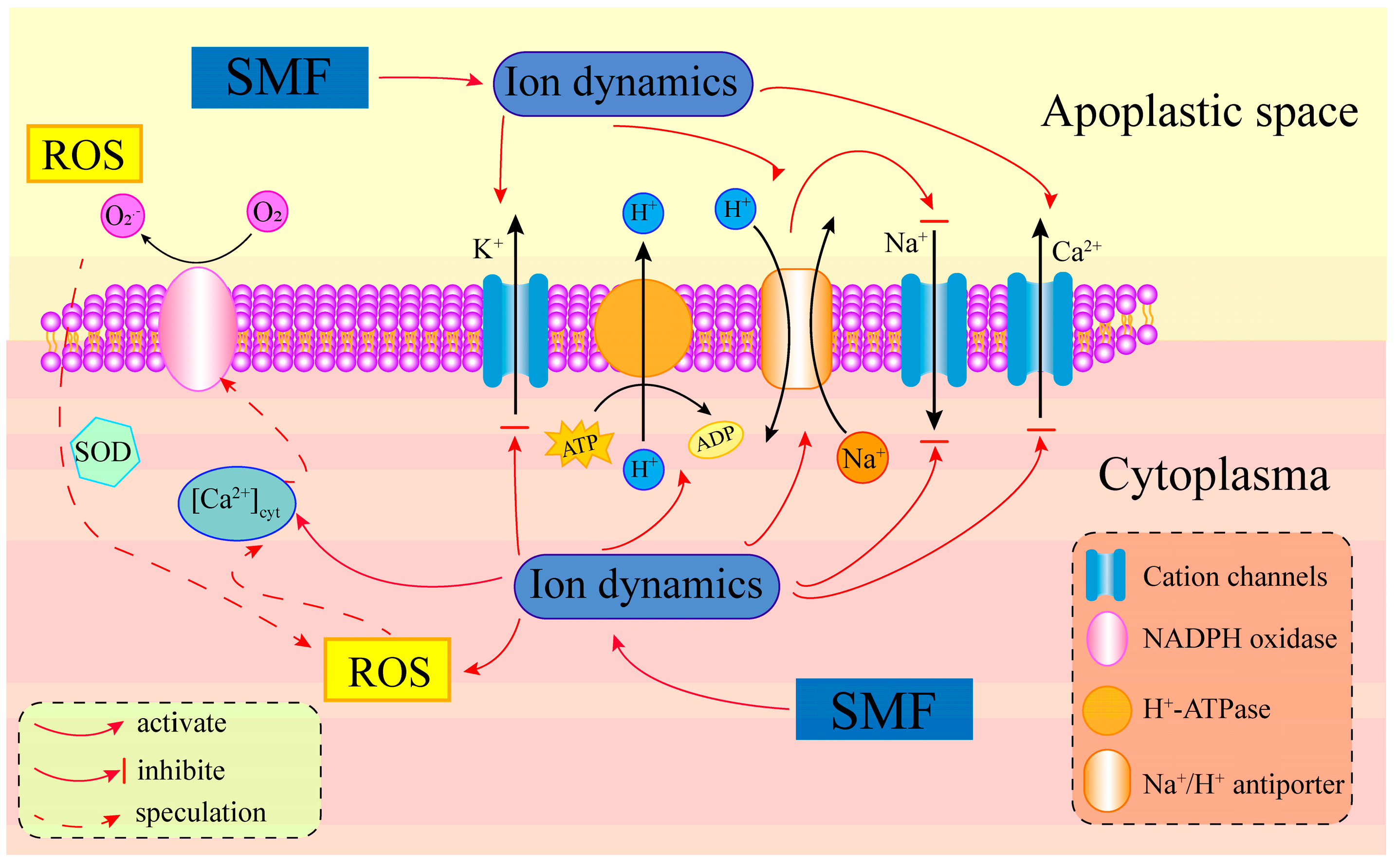
4.2. SMF Improves Poplar Salt Tolerance by Maintaining Ion Homeostasis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic Strategies for Improving Crop Yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling Salt Stress Signaling in Plants: Salt Stress Signaling. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant Abiotic Stress Response and Nutrient Use Efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, Y.; Testerink, C. Root Dynamic Growth Strategies in Response to Salinity. Plant Cell Environ. 2022, 45, 695–704. [Google Scholar] [CrossRef]
- Occhipinti, A.; De Santis, A.; Maffei, M.E. Magnetoreception: An Unavoidable Step for Plant Evolution? Trends Plant Sci. 2014, 19, 1–4. [Google Scholar] [CrossRef]
- Agliassa, C.; Maffei, M.E. Reduction of Geomagnetic Field (GMF) to near Null Magnetic Field (NNMF) Affects Some Arabidopsis Thaliana Clock Genes Amplitude in a Light Independent Manner. J. Plant Physiol. 2019, 232, 23–26. [Google Scholar] [CrossRef]
- Narayana, R.; Fliegmann, J.; Paponov, I.; Maffei, M.E. Reduction of Geomagnetic Field (GMF) to near Null Magnetic Field (NNMF) Affects Arabidopsis Thaliana Root Mineral Nutrition. Life Sci. Space Res. 2018, 19, 43–50. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic Field Effects on Plant Growth, Development, and Evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef]
- Nyakane, N.E.; Markus, E.D.; Sedibe, M.M. The Effects of Magnetic Fields on Plants Growth: A Comprehensive Review. IJFE 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Radhakrishnan, R. Magnetic Field Regulates Plant Functions, Growth and Enhances Tolerance against Environmental Stresses. Physiol. Mol. Biol. Plants 2019, 25, 1107–1119. [Google Scholar] [CrossRef]
- Dodson, C.A.; Hore, P.J.; Wallace, M.I. A Radical Sense of Direction: Signalling and Mechanism in Cryptochrome Magnetoreception. Trends Biochem. Sci. 2013, 38, 435–446. [Google Scholar] [CrossRef]
- Hore, P.J. Are Biochemical Reactions Affected by Weak Magnetic Fields? Proc. Natl. Acad. Sci. USA 2012, 109, 1357–1358. [Google Scholar] [CrossRef]
- Ikeya, N.; Woodward, J.R. Cellular Autofluorescence Is Magnetic Field Sensitive. Proc. Natl. Acad. Sci. USA 2021, 118, e2018043118. [Google Scholar] [CrossRef]
- Maeda, K.; Robinson, A.J.; Henbest, K.B.; Hogben, H.J.; Biskup, T.; Ahmad, M.; Schleicher, E.; Weber, S.; Timmel, C.R.; Hore, P.J. Magnetically Sensitive Light-Induced Reactions in Cryptochrome Are Consistent with Its Proposed Role as a Magnetoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 4774–4779. [Google Scholar] [CrossRef]
- Xu, J.; Jarocha, L.E.; Zollitsch, T.; Konowalczyk, M.; Henbest, K.B.; Richert, S.; Golesworthy, M.J.; Schmidt, J.; Déjean, V.; Sowood, D.J.C.; et al. Magnetic Sensitivity of Cryptochrome 4 from a Migratory Songbird. Nature 2021, 594, 535–540. [Google Scholar] [CrossRef]
- Afzal, I.; Saleem, S.; Skalicky, M.; Javed, T.; Bakhtavar, M.A.; ul Haq, Z.; Kamran, M.; Shahid, M.; Sohail Saddiq, M.; Afzal, A.; et al. Magnetic Field Treatments Improves Sunflower Yield by Inducing Physiological and Biochemical Modulations in Seeds. Molecules 2021, 26, 2022. [Google Scholar] [CrossRef]
- Sarraf, M.; Deamici, K.M.; Taimourya, H.; Islam, M.; Kataria, S.; Raipuria, R.K.; Abdi, G.; Brestic, M. Effect of Magnetopriming on Photosynthetic Performance of Plants. Int. J. Mol. Sci. 2021, 22, 9353. [Google Scholar] [CrossRef]
- Shabrangy, A.; Ghatak, A.; Zhang, S.; Priller, A.; Chaturvedi, P.; Weckwerth, W. Magnetic Field Induced Changes in the Shoot and Root Proteome of Barley (Hordeum vulgare L.). Front. Plant Sci. 2021, 12, 622795. [Google Scholar] [CrossRef]
- Vaezzadeh, M.; Noruzifar, E.; Faezeh, G.; Salehkotahi, M.; Mehdian, R. Excitation of Plant Growth in Dormant Temperature by Steady Magnetic Field. J. Magn. Magn. Mater. 2006, 302, 105–108. [Google Scholar] [CrossRef]
- Liu, X.; Ma, F.; Zhu, H.; Ma, X.; Guo, J.; Wan, X.; Wang, L.; Wang, H.; Wang, Y. Effects of Magnetized Water Treatment on Growth Characteristics and Ion Absorption, Transportation, and Distribution in Populus × Euramericana ‘Neva’ under NaCl Stress. Can. J. For. Res. 2017, 47, 828–838. [Google Scholar] [CrossRef]
- Sun, J.; Chen, S.; Dai, S.; Wang, R.; Li, N.; Shen, X.; Zhou, X.; Lu, C.; Zheng, X.; Hu, Z.; et al. NaCl-Induced Alternations of Cellular and Tissue Ion Fluxes in Roots of Salt-Resistant and Salt-Sensitive Poplar Species. Plant Physiol. 2009, 149, 1141–1153. [Google Scholar] [CrossRef]
- Demidchik, V.; Maathuis, F.J.M. Physiological Roles of Nonselective Cation Channels in Plants: From Salt Stress to Signalling and Development. New Phytol. 2007, 175, 387–404. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global Plant Stress Signaling: Reactive Oxygen Species at the Cross-Road. Front. Plant Sci. 2016, 7, 187. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Dhawi, F.; Al-Khayri, J.M.; Hassan, E. Static Magnetic Field Influence on Elements Composition in Date Palm (Phoenix dactylifera L.). Res. J. Agric. Biol. Sci. 2009, 5, 161–166. [Google Scholar]
- Islam, M.; Maffei, M.E.; Vigani, G. The Geomagnetic Field Is a Contributing Factor for an Efficient Iron Uptake in Arabidopsis Thaliana. Front. Plant Sci. 2020, 11, 325. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Verret, F.; Wheeler, G.; Taylor, A.R.; Farnham, G.; Brownlee, C. Calcium Channels in Photosynthetic Eukaryotes: Implications for Evolution of Calcium-based Signalling. New Phytol. 2010, 187, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Bekhite, M.M.; Figulla, H.-R.; Sauer, H.; Wartenberg, M. Static Magnetic Fields Increase Cardiomyocyte Differentiation of Flk-1+ Cells Derived from Mouse Embryonic Stem Cells via Ca2+ Influx and ROS Production. Int. J. Cardiol. 2013, 167, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Calabrò, E.; Condello, S.; Currò, M.; Ferlazzo, N.; Caccamo, D.; Magazù, S.; Ientile, R. Effects of Low Intensity Static Magnetic Field on FTIR Spectra and ROS Production in SH-SY5Y Neuronal-like Cells: SMF Effects on SH-SY5Y Neuronal-Like Cells. Bioelectromagnetics 2013, 34, 618–629. [Google Scholar] [CrossRef]
- De Nicola, M.; Cordisco, S.; Cerella, C.; Albertini, M.C.; D’Alessio, M.; Accorsi, A.; Bergamaschi, A.; Magrini, A.; Ghibelli, L. Magnetic Fields Protect from Apoptosis via Redox Alteration. Ann. N. Y. Acad. Sci. 2006, 1090, 59–68. [Google Scholar] [CrossRef]
- Solov’yov, I.A.; Chandler, D.E.; Schulten, K. Magnetic Field Effects in Arabidopsis Thaliana Cryptochrome-1. Biophys. J. 2007, 92, 2711–2726. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X. Magnetic Fields and Reactive Oxygen Species. Int. J. Mol. Sci. 2017, 18, 2175. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Stress-Triggered Redox Signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A Mutual Interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Xia, X.-J.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.-Q. Interplay between Reactive Oxygen Species and Hormones in the Control of Plant Development and Stress Tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory Burst Oxidases: The Engines of ROS Signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Steinhorst, L.; He, G.; Moore, L.K.; Schültke, S.; Schmitz-Thom, I.; Cao, Y.; Hashimoto, K.; Andrés, Z.; Piepenburg, K.; Ragel, P.; et al. A Ca2+-Sensor Switch for Tolerance to Elevated Salt Stress in Arabidopsis. Dev. Cell 2022, 57, 2081–2094.e7. [Google Scholar] [CrossRef] [PubMed]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-light Dependent Reactive Oxygen Species Formation by Arabidopsis Cryptochrome May Define a Novel Evolutionarily Conserved Signaling Mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Kattnig, D.R. Radical-Pair-Based Magnetoreception Amplified by Radical Scavenging: Resilience to Spin Relaxation. J. Phys. Chem. B 2017, 121, 10215–10227. [Google Scholar] [CrossRef]
- Mondal, P.; Huix-Rotllant, M. Theoretical Insights into the Formation and Stability of Radical Oxygen Species in Cryptochromes. Phys. Chem. Chem. Phys. 2019, 21, 8874–8882. [Google Scholar] [CrossRef] [PubMed]
- Player, T.C.; Hore, P.J. Viability of Superoxide-Containing Radical Pairs as Magnetoreceptors. J. Chem. Phys. 2019, 151, 225101. [Google Scholar] [CrossRef] [PubMed]
- Zadeh-Haghighi, H.; Simon, C. Magnetic Field Effects in Biology from the Perspective of the Radical Pair Mechanism. J. R. Soc. Interface 2022, 19, 20220325. [Google Scholar] [CrossRef] [PubMed]
- Lopresto, V.; Merla, C.; Pinto, R.; Benvenuto, E. High-Intensity Static Magnetic Field Exposure Devices for in Vitro Experiments on Biopharmaceutical Plant Factories in Aerospace Environments. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 893–896. [Google Scholar]
- McGinley, J.V.M.; Ristic, M.; Young, I.R. A Permanent MRI Magnet for Magic Angle Imaging Having Its Field Parallel to the Poles. J. Magn. Reson. 2016, 271, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Deng, C.; Deng, S.; Li, N.; Zhao, C.; Zhao, R.; Liang, S.; Chen, S. The Arabidopsis Ca2+-Dependent Protein Kinase CPK12 Is Involved in Plant Response to Salt Stress. Int. J. Mol. Sci. 2018, 19, 4062. [Google Scholar] [CrossRef]
- Sun, J.; Wang, M.-J.; Ding, M.-Q.; Deng, S.-R.; Liu, M.-Q.; Lu, C.-F.; Zhou, X.-Y.; Shen, X.; Zheng, X.-J.; Zhang, Z.-K.; et al. H2O2 and Cytosolic Ca2+ Signals Triggered by the PM H+-Coupled Transport System Mediate K+/Na+ Homeostasis in NaCl-Stressed Populus Euphratica Cells: Salt Stress Signalling in P. Euphratica. Plant Cell Environ. 2010, 33, 943–958. [Google Scholar] [CrossRef]
- Li, J.; Bao, S.; Zhang, Y.; Ma, X.; Mishra-Knyrim, M.; Sun, J.; Sa, G.; Shen, X.; Polle, A.; Chen, S. Paxillus Involutus Strains MAJ and NAU Mediate K+/Na+ Homeostasis in Ectomycorrhizal Populus × Canescens under Sodium Chloride Stress. Plant Physiol. 2012, 159, 1771–1786. [Google Scholar] [CrossRef]
- Csillag, A.; Kumar, B.V.; Szabó, K.; Szilasi, M.; Papp, Z.; Szilasi, M.E.; Pázmándi, K.; Boldogh, I.; Rajnavölgyi, É.; Bácsi, A.; et al. Exposure to Inhomogeneous Static Magnetic Field Beneficially Affects Allergic Inflammation in a Murine Model. J. R. Soc. Interface 2014, 11, 20140097. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, Y.; Li, S.; Zhang, W.; Yin, C.; Lin, Y. Regulation of Phytohormones on the Growth and Development of Plant Root Hair. Front. Plant Sci. 2022, 13, 865302. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Sahu, P.K.; Kumar, K.; Pal, G.; Gond, S.K.; Kharwar, R.N.; White, J.F. Endophyte Roles in Nutrient Acquisition, Root System Architecture Development and Oxidative Stress Tolerance. J. Appl. Microbiol. 2021, 131, 2161–2177. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Claeijs, N.; Balcerowicz, D.; Schoenaers, S. Hormonal Regulation of Root Hair Growth and Responses to the Environment in Arabidopsis. J. Exp. Bot. 2020, 71, 2412–2427. [Google Scholar] [CrossRef] [PubMed]
- Van Huizen, A.V.; Morton, J.M.; Kinsey, L.J.; Von Kannon, D.G.; Saad, M.A.; Birkholz, T.R.; Czajka, J.M.; Cyrus, J.; Barnes, F.S.; Beane, W.S. Weak Magnetic Fields Alter Stem Cell–Mediated Growth. Sci. Adv. 2019, 5, eaau7201. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of Superoxide and Hydrogen Peroxide in Arabidopsis Root and Their Influence on Root Development: Possible Interaction with Peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, L.; Zhan, A.; Wang, M.; Tian, L.; Guo, W.; Pan, Y. Long-Term Exposure to a Hypomagnetic Field Attenuates Adult Hippocampal Neurogenesis and Cognition. Nat. Commun. 2021, 12, 1174. [Google Scholar] [CrossRef]
- Demidchik, V.; Tyutereva, E.V.; Voitsekhovskaja, O.V. The Role of Ion Disequilibrium in Induction of Root Cell Death and Autophagy by Environmental Stresses. Funct. Plant Biol. 2018, 45, 28. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium Transport across Plant Membranes: Mechanisms and Functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, H.; Sun, L.; Jiao, Y.; Zhang, G.; Miao, C.; Hao, F. NADPH Oxidase AtrbohD and AtrbohF Function in ROS-Dependent Regulation of Na+/K+ Homeostasis in Arabidopsis under Salt Stress. J. Exp. Bot. 2012, 63, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Bai, T.; Wei, H.; Gardea-Torresdey, J.L.; Keller, A.; White, J.C. Nanobiotechnology-Based Strategies for Enhanced Crop Stress Resilience. Nat. Food 2022, 3, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, Y.; Yang, Y.; Wu, H.; Wang, D.; Liu, J. Involvement of Hydrogen Peroxide and Nitric Oxide in Salt Resistance in the Calluses from Populus Euphratica. Plant Cell Env. 2007, 30, 775–785. [Google Scholar] [CrossRef]
- Conrath, U. Molecular Aspects of Defence Priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Fichman, Y.; Zandalinas, S.I.; Sengupta, S.; Burks, D.; Myers, R.J.; Azad, R.K.; Mittler, R. MYB30 Orchestrates Systemic Reactive Oxygen Signaling and Plant Acclimation. Plant Physiol. 2020, 184, 666–675. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic Signaling during Abiotic Stress Combination in Plants. Proc. Natl. Acad. Sci. USA 2020, 117, 13810–13820. [Google Scholar] [CrossRef]
- Locato, V.; Cimini, S.; De Gara, L. ROS and Redox Balance as Multifaceted Players of Cross-Tolerance: Epigenetic and Retrograde Control of Gene Expression. J. Exp. Bot. 2018, 69, 3373–3391. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Zhang, H.; Han, W.; Wang, N.; Ma, S.; Ma, F.; Tian, H.; Wang, Y. Physiological Responses Revealed Static Magnetic Fields Potentially Improving the Tolerance of Poplar Seedlings to Salt Stress. Forests 2024, 15, 138. https://doi.org/10.3390/f15010138
Hu J, Zhang H, Han W, Wang N, Ma S, Ma F, Tian H, Wang Y. Physiological Responses Revealed Static Magnetic Fields Potentially Improving the Tolerance of Poplar Seedlings to Salt Stress. Forests. 2024; 15(1):138. https://doi.org/10.3390/f15010138
Chicago/Turabian StyleHu, Jihuai, Haojie Zhang, Wenhao Han, Nianzhao Wang, Shuqi Ma, Fengyun Ma, Huimei Tian, and Yanping Wang. 2024. "Physiological Responses Revealed Static Magnetic Fields Potentially Improving the Tolerance of Poplar Seedlings to Salt Stress" Forests 15, no. 1: 138. https://doi.org/10.3390/f15010138
APA StyleHu, J., Zhang, H., Han, W., Wang, N., Ma, S., Ma, F., Tian, H., & Wang, Y. (2024). Physiological Responses Revealed Static Magnetic Fields Potentially Improving the Tolerance of Poplar Seedlings to Salt Stress. Forests, 15(1), 138. https://doi.org/10.3390/f15010138