Abstract
Mangrove ecosystems play important roles in mitigating climate change but are suffering from an area decrease. The sediment in a mangrove ecosystem is a crucial component in estuarine carbon and nutrient cycles. However, it is unclear whether the restoration of mangroves enhances their sediment carbon sequestration and nutrient storage. To investigate the restoration effects of native Aegiceras corniculatum mangrove forests, the contents and density of sediment organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) were quantified in an unvegetated mudflat (MF), an area with mature A. corniculatum (AC), and two restored A. corniculatum mangroves planted in 2002 (02AC) and 2008 (08AC) in Quanzhou, southeastern China. Compared with the MF, mangrove restoration increased the SOC and TN contents in the sediment, primarily at the 0–40 cm depth for SOC and 0–20 cm for TN, while the TP content was not impacted by restoration. The sediment under AC showed a higher SOC content only at the 0–10 cm and 20–40 cm depths compared to restored 02AC and 08AC. The restoration of A. corniculatum mangroves did not significantly increase the stock of sediment SOC, TN, and TP relative to the MF, which might be attributed to the decrease in bulk density and the relatively short restoration period. Thus, further continuous monitoring is needed to verify whether the sediment carbon pool can be promoted on a longer-term scale by the restoration of A. corniculatum.
1. Introduction
Mangroves are woody plant communities growing in subtropical regions that are among the ecosystems with the highest primary productivity in the world and play an important role in maintaining biodiversity, food-web energy supply, and nutrient circulation [1,2]. Mangroves store large amounts of CO2 in plant biomass through their high primary productivity. The average mangrove biomass in equatorial regions can reach 247 ton/hm2, which is comparable to that of rainforests [2]. Most of the organic material may be stored in mangrove sediments and dead roots, making the mangrove forest a huge blue carbon reservoir [3]. Moreover, the preservation of nutrients, such as nitrogen and phosphorus, has an important impact on the growth of mangroves themselves and the circulation of ecosystem substances [4]. Due to the effects of excessive logging and aquaculture, the mangrove area globally has decreased by 30%–50% in the past half-century [3,5]. With the understanding of the function of mangrove carbon sinks, the protection and restoration of mangroves has gradually received attention, and the impact of global changes brought about by the rise in CO2 can be alleviated through the restoration of mangrove ecosystems [1,2]. Mangrove restoration is generally slow, and the restoration of the mangrove community structure does not mean the restoration of the ecological function [6]. Research on the long-term scale of the mangrove ecological function restoration effect is limited [7], and the underground carbon stock function and other changes in the wetland restoration process are frequently neglected [8]. A large number of mangrove restoration projects have been conducted since the 1990s in China and significantly increased the area of mangrove forests [9]. However, it has been pointed out that a large gap exists between the structure and function of wetlands and pristine wetlands after long-term restoration and that the restoration of the sediment biochemical cycle is slower than the restoration of the wetland biological community structure [10]. Therefore, it is necessary to assess the restoration effects to better understand whether mangrove forests can rapidly enhance the sediment carbon and nutrient reservoirs.
The mangrove wetlands of the Luoyang River are located on the northern edge of the mangrove distribution in China, and the mangrove area is seriously damaged. Since 2002, a large-scale mangrove afforestation project has been carried out by the local government, and the cumulative afforestation area is 420 hm2, making an important contribution to the restoration of the mangrove area. In recent years, mangrove planting has brought about a series of ecological and social problems; the high-density planting and expansion of mangroves has contributed to sediment deposition and is considered a threat to the safety of the historically significant Luoyang Bridge. The large number of mangrove planting restorations promotes an increase in the mangrove forest area but also plays an important role in the coastal wetlands in the bare mudflat area [10,11]. At the same time, mangrove stasis may lead to changes in the original habitat sediment deposition process, thus affecting the physical and chemical properties of the sediment. The accumulation of mangrove biomass and the burial of litter generally promote the improvement of sediment organic carbon (SOC) reserves, and the absorption and removal of nitrogen and phosphorus in mangrove sediments are also ecologically important and an important way to ensure the growth of mangrove plants [1,4,9].
In this study, taking the typical restoration species in Quanzhou Bay as the research object, Aegiceras corniculatum with different restoration times was selected to discuss the influence of the mangrove restoration process on the ecosystem from the perspective of sediment carbon, nitrogen, and phosphorus contents and stocks to provide a scientific basis for the development of mangrove restoration projects. Due to the slow growth of mangroves, the objective of this study was to investigate whether the sediment carbon, nitrogen, and phosphorus reservoirs increased with the development of mangrove restoration compared with the bare mudflat.
2. Materials and Methods
2.1. Sampling Site
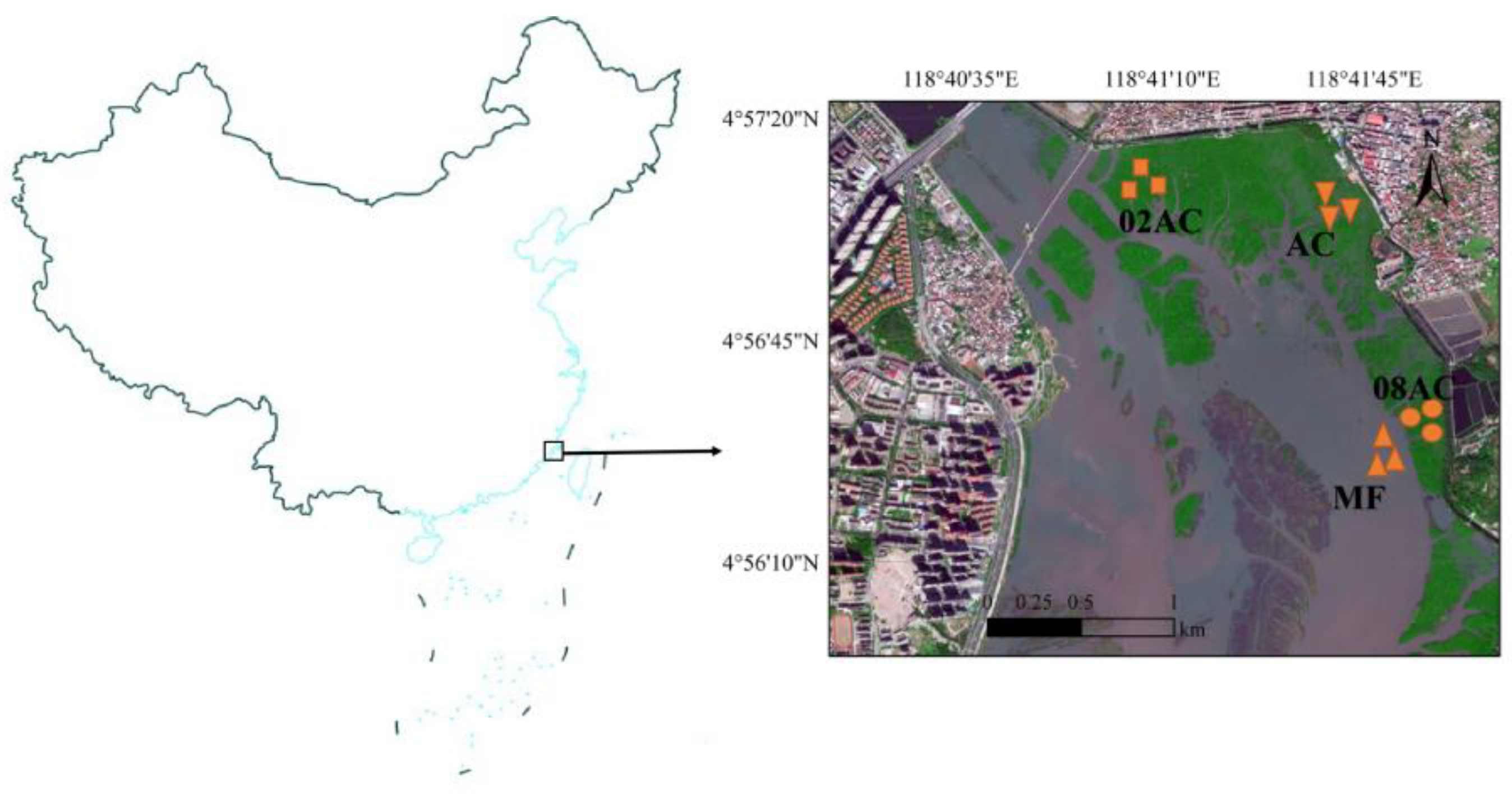
The sampling site of this study, the Luoyang River mangrove, is located at the mouth of the Luoyang River in Quanzhou (24°56′30.77″ N, 118°41′42.47″ E). It has three mangrove species: A. corniculatum, Kandelia obovata, and Aricennia marina. This area belongs to the south subtropical tropical marine monsoon climate zone, with an average temperature of 20 °C, an extreme minimum temperature of 0 °C occurring from January to February, and a maximum temperature of 38.19 °C. The annual precipitation is between l009 and l200 mm, and the annual evaporation is approximately 2000 mm. It has a semi-diurnal tide with an average range of 4.52 m, and the seawater salinity ranges from 3.5 to 28.9‰ in the estuarine area [11]. Since 2002, mangrove restoration and planting projects have been carried out, and the main species are A. corniculatum and Kandelia obovata. Compared with 1993, more than 420 ha of mangroves have been restored, and the current mangrove area has increased by nearly 10 times and reached 700 ha. The Luoyang River mangrove is the northernmost limit of the natural distribution of A. corniculatum in the western Pacific. In this study, we chose a bare mudflat area (MF), an A. corniculatum mangrove planted in 2008 (08AC), an A. corniculatum mangrove planted in 2002 (02AC), and a native mangrove area (AC) as the sampling sites to collect the sediment samples (Figure 1).
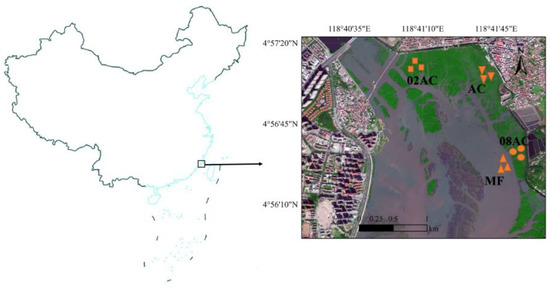
Figure 1.
Map of the sampling site in Quanzhou, southeastern China (MF: bare mudflat area; 08AC: A. corniculatum planted in 2008; 02AC: A. corniculatum planted in 2002; and native AC: A. corniculatum mangrove area).
2.2. Sample Collection and Analysis
Wetland sediment samples were collected in August 2015. At each sampling site, three quadrats were randomly chosen, with a distance of approximately 50 m between them. Two sediment cores at a depth of 0–100 cm were collected using a sediment core sampler (C040903, Eijkelkamp, Giesbeek, Holland) and then divided into the following sections: 0–10, 10–20, 20–40, 40–60, 60–80, and 80–100 cm. Sediments at the same depth from the two cores were pooled together as one replicate in each quadrat. Then, 72 samples of sediments were dried using a vacuum freeze dryer. Additionally, the physical and chemical indexes of the samples were determined after they were filtered through a 2 mm sieve, including bulk density, water content, salinity, and pH. The soil salinity and pH were measured using a multiple-parameter water analyzer (Thermal A329, Waltham, MA, USA) after samples were extracted with distilled water (water–soil ratio of 5:1 for salinity and 2.5:1 for pH). All sediment samples were then ground and filtered through a 100 mm sieve to measure the SOC and total nitrogen (TN) content using an Elementar Macrocube element analyzer. Before the organic carbon test, 1M hydrochloric acid was used to remove sediment inorganic carbon. After digestion with potassium persulfate, total phosphorus (TP) was measured using phospho-molybdenum spectrophotometry.
The total sediment carbon stock, TN stock, and TP stock in 0–100 cm sediment cores were calculated using the following equation:
where X is TOC, TN, or TP content; Xi is the corresponding source factor index content of sediment in layer i; BDi is the sediment bulk density in layer i; and Di is the depth interval of sediment in layer i.
2.3. Data Analysis
Two-factor analysis of variance (ANOVA) was used to compare the differences in the physical and chemical indexes of sediment element contents between different sampling depths and sampling sites. Principal component analysis (PCA) was used to compare the differences and correlations. ANOVA was performed using SPSS 19.0, and PCA analysis was performed using the R language Vegans software package. The figures were generated using Sigmaplot 12.0.
3. Results
3.1. Physical and Chemical Properties of Sediment
The results of two-way ANOVA (Table 1) showed that depth did not have a significant effect on salinity, and the interaction between the depth and site did not have a significant effect on the sample content, water content, bulk density, or C:N value. However, the other sediment indicators were all significantly affected by the site and depth and their interaction. This indicates that the restoration process leads to changes in the physicochemical properties of the sediment.

Table 1.
Two-way ANOVA results of site and depth effects on sediment properties in the Luoyang River estuary (df: Degree of Freedom, F: Equality of Variances, P: Probability value).
When comparing sediments at the same depth between different sites, the differences in bulk density and pH were significant (Table 2). Restoration led to a decrease in bulk density; this change was most significant in the upper sediment, at 0–10 and 10–20 cm. The MF had the highest average bulk density at 0–10 cm, but a significant difference was only found between the MF and AC. At a depth of 10–20 cm, the highest bulk density was in the MF and the lowest was in AC, while the difference between 08AC and 02AC was not significant. The sediment bulk density increased with depth at all sites.
Table 2.
Environmental parameters of sediment for each site (mean ± standard deviation; letters indicate significant differences in the same sediment layer between different sites at p < 0.05; MF: bare mudflat area; 08AC: A. corniculatum planted in 2008; 02AC: A. corniculatum planted in 2002; and native AC: A. corniculatum mangrove area).
Sediment pH values showed significant differences among sites. The pH showed a downward trend with increased restoration time. The MF had the highest sediment pH value at all depths. There was no significant difference in pH between 02AC and 08AC at 0–10 cm, but 02AC had a lower pH than 08AC in the deeper layers, except for at 80–100 cm. Compared with AC, the pH of 02AC was significantly lower, except at 0–10 cm. However, the sediment pH was significantly higher in AC than in 08AC at a depth of 0–60 cm, while there was no significant difference at a depth of 60–100 cm. The sediment pH at all sites tended to increase gradually with depth.
3.2. Carbon, Nitrogen, and Phosphorus Contents and Their Vertical Changes
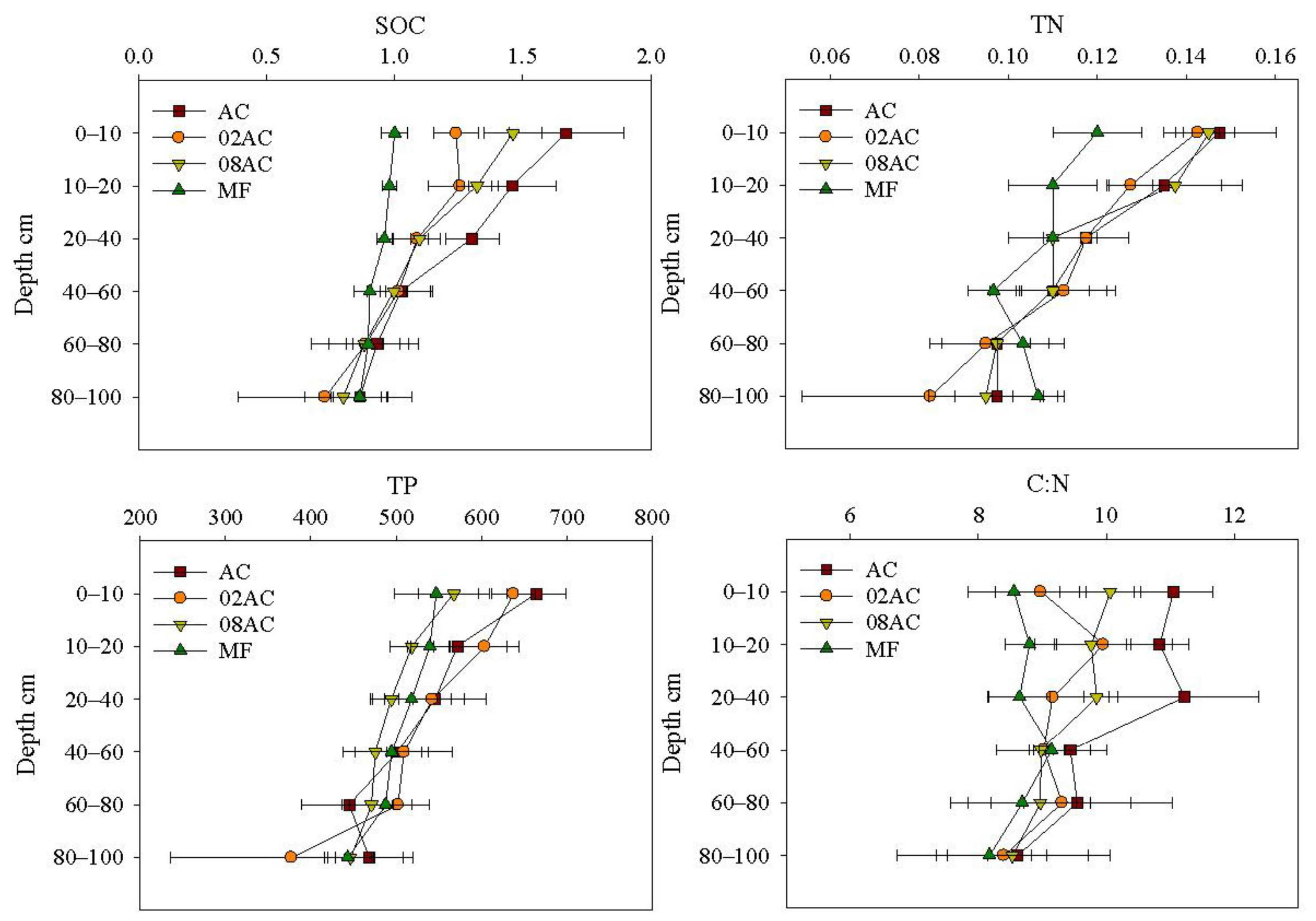
For the SOC content (Figure 2), AC had the highest mean at a depth of 0–100 cm. At a depth of 0–10 cm, AC had a significantly higher SOC content than the other sites. The SOC content in 08AC and 02AC was also significantly higher than that in the MF. At a depth of 10–20 cm, the mean SOC content of AC was higher than that of 08AC and 02AC, but the difference between the three sites was not significant, and all three had significantly higher SOC content than the MF. At the 20–40 cm depth, the SOC content at AC was significantly higher than at the other sites. However, the differences between 02AC, 08AC, and the MF were not significant. At a depth of 40–100 cm, the SOC content of all sites was not insignificantly different. The SOC content of all sites tended to decrease with depth, indicating that the effects of restoration or vegetation growth on the sediment SOC content were mainly concentrated at a depth range of 0–40 cm.
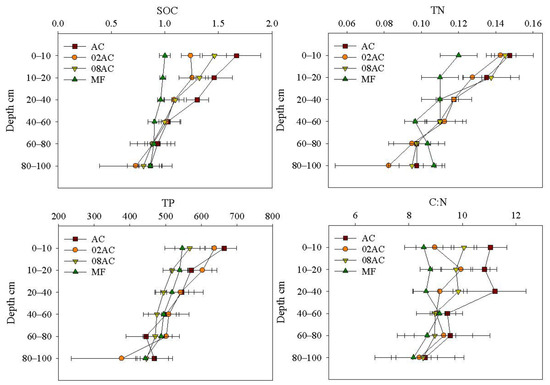
Figure 2.
Vertical distribution of sediment organic carbon, total nitrogen, and total phosphorus contents and SOC:TN ratios from different sampling sites in the Luoyang River estuary (mean ± standard deviation; MF: bare mudflat area; 08AC: A. corniculatum planted in 2008; 02AC: A. corniculatum planted in 2002; and native AC: A. corniculatum mangrove area).
For the TN content of the sediment, the difference between the AC of the two restoration sites and natural sites was not significant, and the difference between the MF of the mangrove forest occurred only at a depth of 0–20 cm. As the depth increased, the TN content of each mangrove forest gradually decreased, but in the MF, it first decreased and then increased. This suggests that vegetation growth affected the TN content in surface sediments and did not increase with restoration time.
For sediment TP content, differences were only found in the surface sediment (0–10 cm). It was significantly higher in AC than in 08AC and the MF, but AC was not significantly different from 02AC, and 02AC was only significantly higher than the MF. The TP content of all sampling sites showed a trend of decreasing with depth. This indicates that the growth of A. corniculatum vegetation has little effect on TP content and a limited cumulative effect.
The C:N value reflects the source of sediment organic matter. AC had the highest sediment C:N at different depths. At 0–10 cm, the sediment C:N values were significantly higher at AC than at 02AC and the MF, and they were significantly higher at 08AC than at the MF. However, the difference between 02AC and MF was not significant. At 10–20 cm, significant differences occurred only between AC and the MF. At a 20–40 cm depth, they were significantly higher at AC than at 02AC and the MF, while no significant differences were observed between 08AC and AC or between 02AC and the MF. At a depth range of 40–100 cm, the differences in sediment C:N values were not significant for any of the sample sites. This shows that the contribution of vegetation to the composition of sediment organic matter has increased; the sediment organic matter at all sites is a mixture of marine microalgae and land-derived organic matter.
3.3. Changes in Carbon, Nitrogen, and Phosphorus Density in the Sediment
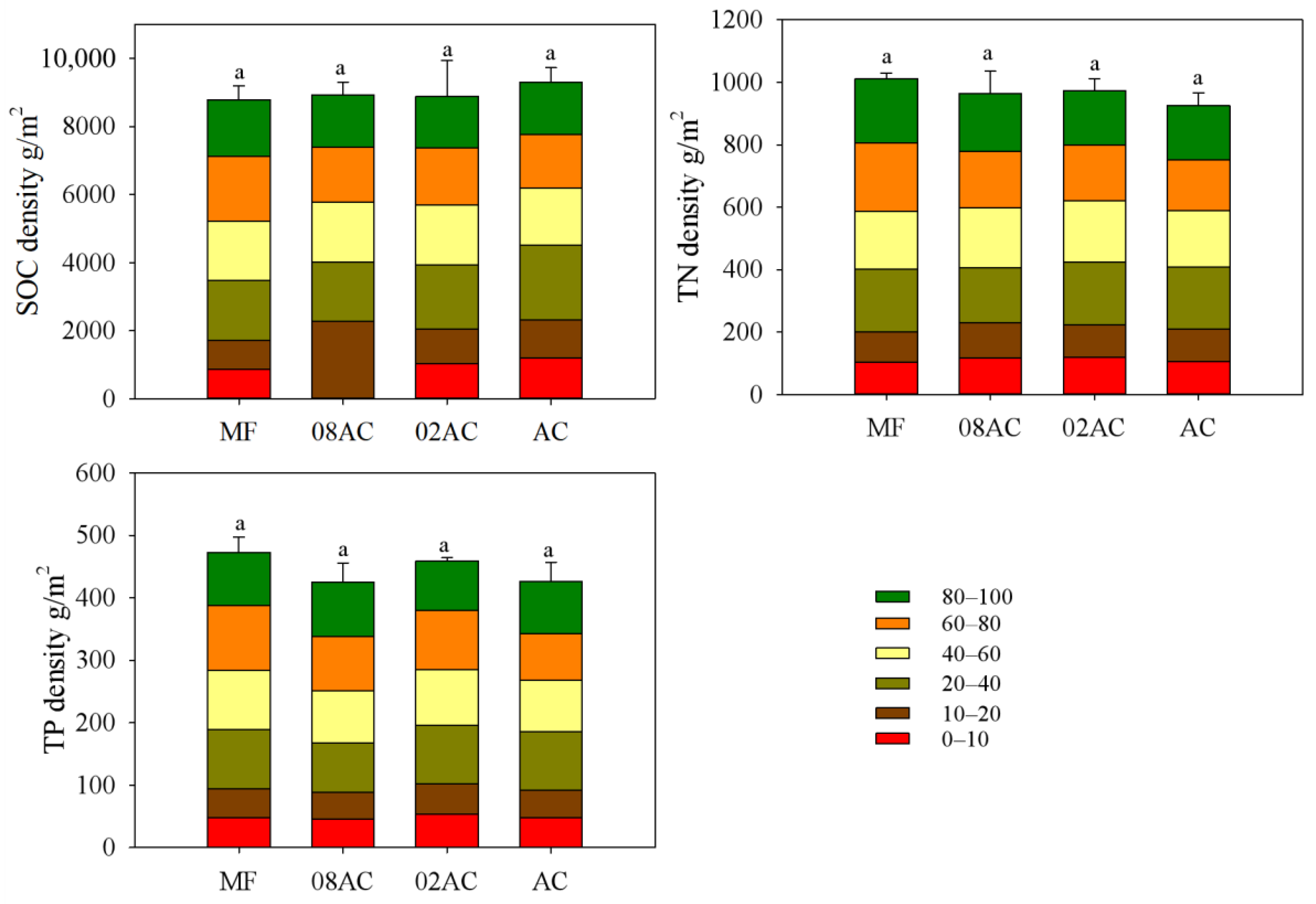
By calculating the SOC density, TN density, and TP density at depths from 0 to 100 cm (Figure 3), it was found that the restoration and growth of A. corniculatum vegetation did not significantly improve the sediment carbon, nitrogen, or phosphorus stock. As shown in Figure 4, the differences between sites mainly manifested at a depth of 0–10 cm, where the vegetation sample sites showed a higher organic carbon density than the mudflats. The presence of vegetation decreased the sediment TN and TP density to some extent.
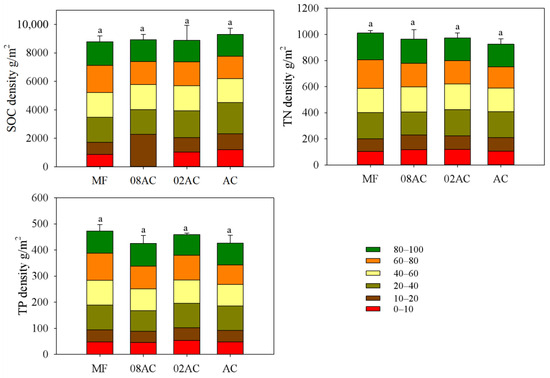
Figure 3.
Comparison of the pools of organic carbon, total nitrogen, and total phosphorus in the 100 cm sediment at each sampling site in the Luoyang River estuary (error bar represents standard deviation, and letters indicate significant differences at p < 0.05; MF: bare mudflat area; 08AC: A. corniculatum planted in 2008; 02AC: A. corniculatum planted in 2002; and native AC: A. corniculatum mangrove area).
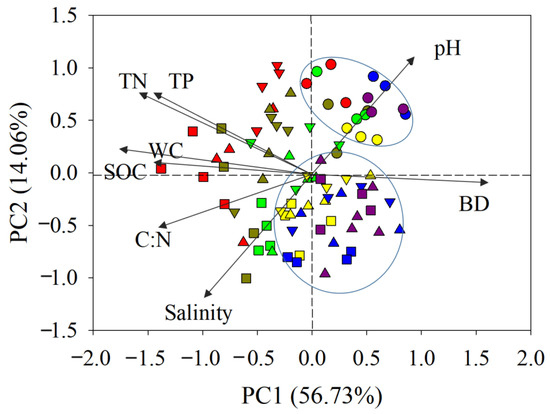
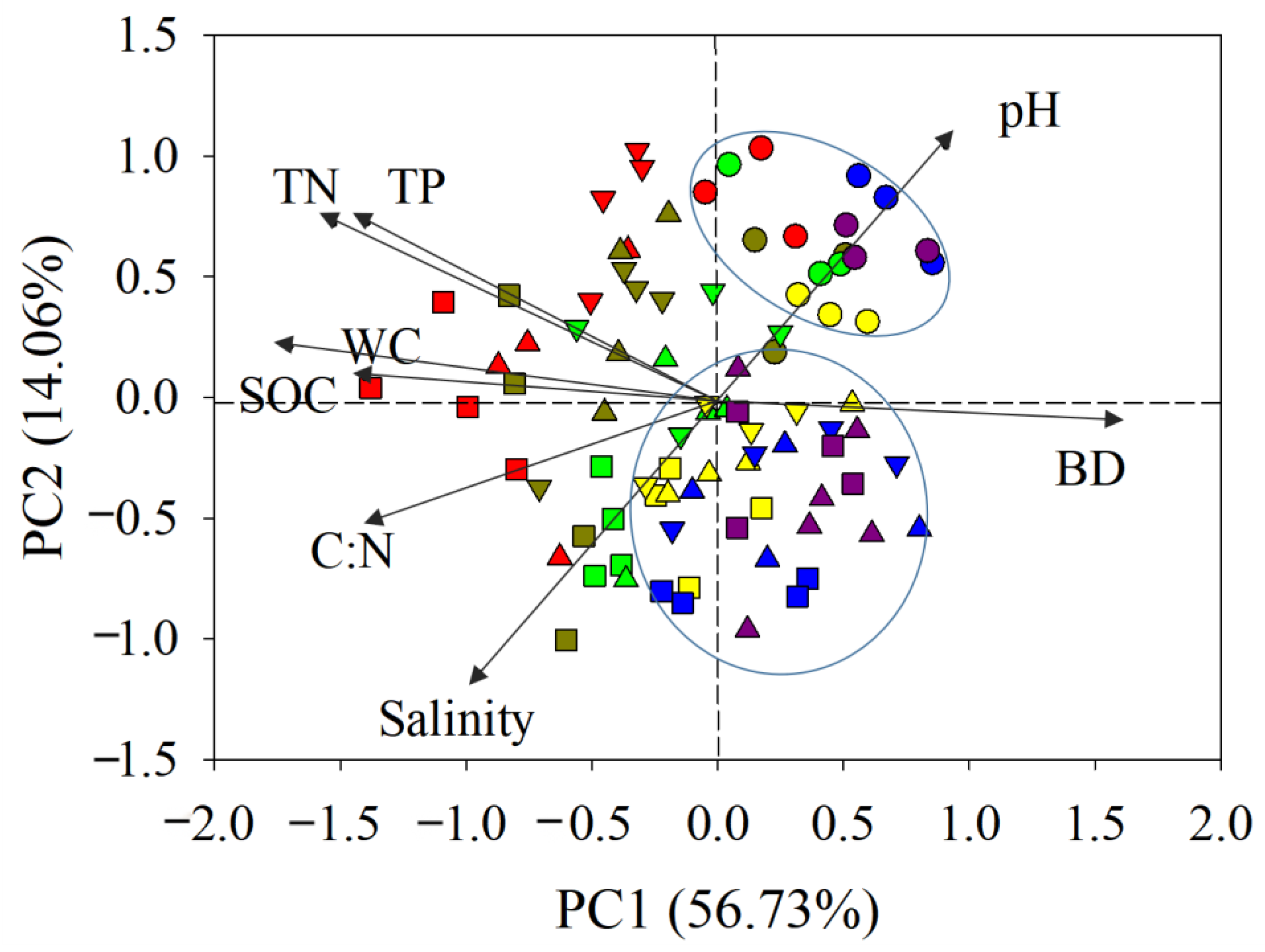
Figure 4.
PCA analysis showing the relationship between sediment biogenic contents and environmental parameters in the Luoyang River estuary (square: AC; inverted triangle: 02AC; triangle: 08AC; circle: MF; red: 0–10 cm; dark green: 10–20 cm; bright green: 20–40 cm; yellow: 40–60 cm; blue: 60–80 cm; purple: 80–100 cm; MF: bare mudflat area; 08AC: A. corniculatum planted in 2008; 02AC: A. corniculatum planted in 2002; and native AC: A. corniculatum mangrove area).
3.4. PCA Analysis of Sediment Carbon
A PCA analysis of SOC showed that the MF plots were clearly distinguished (Figure 4). In other sites, sediments from the 40–60, 60–80, and 80–100 cm layers clustered together, showing high similarity, and also clustered in the right half of the first principal component with the MF. Although the other sample sites were relatively scattered, all of them were mainly distributed in the left half of the first principal component, indicating their high similarity. PC1 was negatively correlated with SOC content and TN content, but positively correlated with sediment bulk density, which mainly reflects the vegetation and depth. PC2 was positively associated with pH and showed a negative relationship with salinity. SOC showed a high positive correlation with TN and TP but a negative correlation with the water content and bulk density, respectively, which was mainly caused by vegetation factors.
4. Discussion
4.1. Effect of Mangrove Restoration on Carbon, Nitrogen, and Phosphorus Contents
Coastal wetlands are located at the interface between land and sea, where terrestrial and marine materials intermix, making coastal wetlands important storage sites for carbon, nitrogen, and phosphorus. The sediment plays important roles in element accumulation, transformation, and transportation, impacting wetlands’ primary productivity and biogeochemical cycle processes [9,12], which are closely related to their vegetation communities [13,14] that can increase the contents of important nutrients around their roots through their root exudates [13]. The sediment texture, salinity, organic material, and nutrient availability constrain the mangrove restoration process and change with mangrove growth and development [15,16].
Generally, carbon and nitrogen elements in sediments gradually accumulate with the input and burial of mangrove litter, including leaves and roots [1,17]. In this study, the SOC content of sediments at a depth of 0–40 cm increased significantly with mangrove restoration due to the fact that the shallow sediment was easily affected by the input of litter from exogenous substances, including marine sources and mangrove vegetation. Although the SOC content of the surface sediment was significantly higher than that of the mudflat, the SOC content of all mangrove sites was less than 2%, which is far from the high carbon content of mangroves (8.54%) estimated by Durate et al. [18] and significantly lower than the global average carbon content of mangrove sediment (2.2%) summarized by Kristensen et al. [19]. Yu et al. found that the SOC content of the surface sediments of mangroves was between 1.3 and 1.6% in the Jiulongjiang estuary in China, which was significantly higher than that of S. alterniflora (1%–1.2%) and mudflats (<0.9%) [20] and did not fluctuate significantly seasonally [21]. Hainan Province has the largest and oldest mangrove forest in China, and the A. corniculatum forest in Haikou and Wenchang showed the highest average organic carbon content (larger than 5%), showing that forest age played an important role in the accumulation of organic carbon [22]. However, the SOC content in the same mangrove species showed high variation, reflecting that the SOC content was determined not only by mangrove vegetation, litter yield, and forest age but also by the sediment deposition rate, hydrologic characteristics, and other factors, such as pollutant input.
4.2. Effect of Mangrove Restoration on Carbon, Nitrogen, and Phosphorus Stocks
Mangrove afforestation is considered an effective way to increase ecosystem carbon stocks [1,9,23]. The average carbon density of the 8–9 kg/m2 A. corniculatum community in this study was far lower compared with the 70-year-old K. obovata forest (15.2 kg/m2) in Shenzhen [24], indicating that the restoration time might influence the sediment carbon stock and that the old mature K. obovata forest may have more litter accumulated and buried in the sediment. Compared to the mudflats, sediment carbon storage did not show any advantages, reflecting that short-term mangrove restoration increases mangrove vegetation biomass, but the enhancement of sediment carbon content and storage is limited. For example, in this study, 14-year-old 02AC only increased the surface SOC content by 24% compared with the mudflat, and the content decreased rapidly with the increase in depth. Additionally, the growth of vegetation leads to a change in the sediment texture and structure, and the sediment bulk density declines significantly, resulting in a lack of increase in the overall carbon stock compared with the mudflat, which has a higher bulk density. Although the carbon density of restored mangroves in this study is lower than that of the 70-year-old K. obovata forest in Shenzhen, it is similar to the 60-year-old A. corniculatum forest in Zhanjiang [24,25]. To some extent, this indicates that A. corniculatum, a rapidly growing shrub mangrove, may not have an advantage in carbon fixation [11]. The results also remind researchers and policymakers to fully consider differences in the mangrove tree species, forest age, and geographical area in estimating the mangrove carbon sequestration capacity and in conducting mangrove restoration projects.
The restoration process of A. corniculatum forests in Quanzhou Bay showed some promoting effects on the increase in TN content in the surface sediment. Compared to the non-rhizosphere sediment, the mangrove rhizosphere sediment generally has a lower C:N value and higher organic nitrogen content due to the high activity of rhizosphere microorganisms and the mangrove root retention of essential nutrients [19,26,27]. This can meet the nutrient requirements for the growth of mangroves under the conditions of N limitation or N export in intertidal areas [13]. Additionally, the anaerobic environment of mangrove wetland sediments caused by periodic flooding is not conducive to the mineralization of organic nitrogen and can lead to the accumulation of nitrogen in the sediments [10]. In this study, although the growth of A. corniculatum had a certain effect on the TN content of the sediment relative to the mudflat, the differences among the communities with different ages were not significant. It is possible that the accumulation of sediment nutrients is mainly used for plant growth, leading to a balance between storage and utilization.
The decomposition and burial of phosphorus from mangrove litter in the sediment can also increase the sediment phosphorus content. However, the phosphorus content in the sediment usually shows a negative correlation with pH [9,12], and the existence of OH− in the sediment promotes the release of phosphorus. With the growth of A. corniculatum trees, sediment pH decreases gradually, leading to a higher sediment phosphorus content in mangroves than in mudflats. The TP content showed a positive relationship with the water content and a negative relationship with the sediment bulk density. The restoration of mangroves leads to a decrease in sediment bulk density, promoting its permeability and storage capacity and thus promoting the accumulation of phosphorus elements in the sediment [12]. As the depth increases, the effect of vegetation on the phosphorus disappears due to the young forest age of A. corniculatum in Quanzhou, where the amount and depth of litter buried in sediment are limited. However, the phosphorus element mainly comes from exogenous material settlement and lacks direct exchange with the atmosphere; thus, the circulation process is conservative [9] and is not easily influenced by wetland destruction and vegetation restoration during the ecological process.
There is a significant positive correlation among carbon, nitrogen, and phosphorus contents in the mangrove sediments of Quanzhou Bay, indicating a relatively consistent source of sediment organic matter. C:N values can provide some evidence of the source of sediment organic matter. The marine C:N value is usually between 4 and 10, while the C:N value of terrigenous plants is above 20 [19,28], and the C:N value of mangrove plants is even above 80 [29]. The C:N values of all sediment deposits in the study area were between 8 and 11, indicating that marine source materials might be the main contributor to sediment organic matter in the ecosystem. The growth of mangrove plants promotes the value of the sediment C:N ratio, indicating the accumulation of mangrove source material in the sediment. However, the limited increase in the C:N value reflects the limited contribution of mangrove plant restoration to the carbon and nitrogen reservoirs of the sediments in Quanzhou Bay. Additionally, the sediment C:N value in 02AC was lower than or similar to that in the mudflat, which may be due to the fact that its location is closer to the river channel compared to the mudflat.
4.3. Implication of A. corniculatum Restoration in Quanzhou Bay
The planting of A. corniculatum mangroves has a positive effect on the area of mangrove recovery, but the development of mangroves can also effectively slow the flow of water and promote sedimentation. The hydrodynamics of the Luoyang River are weak due to the upper river dam; thus, together with mangrove restoration, the river deposition and siltation effect is more effective. This has gradually endangered the thousand-year-old ancient Wan’an Bridge and has also attracted public attention. Shih et al. studied mangrove overplanting in Taiwan and found that dense mangrove planting may lead to an increase in the probability of flooding; they proposed that 20% mangrove thinning could reduce the frequency of flooding and would not significantly affect the mangrove carbon reservoir [30]. This provides inspiration for the subsequent development of a mangrove planting project in Quanzhou Bay and in other areas in China. Mangrove restoration is not just about expanding the area but also about restoring the mangrove ecosystem service function. A detailed study of the silting effect and sedimentation rate of mangrove planting is necessary to explore the relationship between mangrove thinning and siltation effects to achieve a more scientific mode of mangrove restoration. Additionally, mangrove planting and growth decrease the area of mudflats, which is an important habitat for the bivalve benthic animal community that feeds coastal birds [31]. Therefore, the disappearance of mudflats will further impact the survival of mangrove birds. The selection of suitable sites, mangrove species, and density and the reasonable thinning of existing mangrove plants are of great significance for the maintenance of the ecosystem functions of mangrove ecosystems.
5. Conclusions
According to the results of this study, A. corniculatum mangrove planting increased the surface sediment carbon and nitrogen contents, but it did not promote the improvement of ecosystem carbon storage or nitrogen and phosphorus nutrient pools quickly or significantly in Quanzhou, China. When estimating the carbon fixation capacity of mangroves, the restoration species, restoration age, latitude position, and hydrological conditions of the restoration site should be fully considered to ensure the accuracy and validity of the data. Additionally, the results from this study area showed that in the short term, the planting of A. corniculatum only increased biomass and promoted CO2. However, the fixation and accumulation of mangrove plants did not significantly change the sediment carbon, nitrogen, or phosphorus pools. Further continuous monitoring is needed to verify whether the sediment carbon pool can be promoted on a longer-term scale.
Author Contributions
Conceptualization, J.F. and L.W.; methodology, J.F. and L.W.; investigation, J.F.; writing—original draft preparation, N.H.; writing—review and editing, N.H., L.W., Y.Z., M.W. and J.F.; funding acquisition, J.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Guangdong Provincial Forestry Science and Technology Innovation Project (2023KYXM09), the Southern Marine Science and Engineering Guangdong Laboratory(Zhuhai) (SML2023SP220) and the National Natural Science Foundation of China (41976160). It was also partly funded by the State Key Laboratory of Tropical Oceanography, South China Sea Institute of Oceanology, Chinese Academy of Sciences (LTO2205), and the Research Fund Program of Guangdong Provincial Key Laboratory of Marine Resources and Coastal Engineering.
Data Availability Statement
MDPI Research Data Policies at https://www.mdpi.com/ethics (accessed on 15 August 2015).
Acknowledgments
We thank Jinjiang Chen for the field sampling work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alongi, D.M. Impacts of climate change on blue carbon stocks and fluxes in mangrove forests. Forests 2022, 13, 149. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling and storage in mangrove forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Suwa, R.; Hagihara, A. Carbon and nitrogen pools in a mangrove stand of Kandelia obovata (S., L.) Yong: Vertical distribution in the soil–vegetation system. Wetl. Ecol. Manag. 2007, 15, 141–153. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- McKee, K.L.; Faulkner, P.L. Restoration of biogeochemical function in mangrove forests. Restor. Ecol. 2000, 8, 247–259. [Google Scholar] [CrossRef]
- Osland, M.J.; Spivak, A.C.; Nestlerode, J.A.; Lessmann, J.M.; Almario, A.E.; Heitmuller, P.T.; Russell, M.J.; Krauss, K.W.; Alvarez, F.; Dantin, D.D. Ecosystem development after mangrove wetland creation: Plant–soil change across a 20-year chronosequence. Ecosystems 2012, 15, 848–866. [Google Scholar] [CrossRef]
- Irving, A.D.; Connell, S.D.; Russell, B.D. Restoring coastal plants to improve global carbon storage: Reaping what we sow. PLoS ONE 2011, 6, e18311. [Google Scholar] [CrossRef]
- Feng, J.; Cui, X.; Zhou, J.; Wang, L.; Zhu, X.; Lin, G. Effects of exotic and native mangrove forests plantation on soil organic carbon, nitrogen, and phosphorus contents and pools in Leizhou, China. Catena 2019, 180, 1–7. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Power, M.E.; Comín, F.A.; Yockteng, R. Structural and functional loss in restored wetland ecosystems. PLoS Biol. 2012, 10, e1001247. [Google Scholar] [CrossRef]
- Chen, S.; Chen, B.; Chen, G.; Ji, J.; Yu, W.; Liao, J.; Chen, G. Higher soil organic carbon sequestration potential at a rehabilitated mangrove comprised of Aegiceras corniculatum compared to Kandelia obovata. Sci. Total Environ. 2021, 752, 142279. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhou, J.; Wang, L.; Cui, X.; Ning, C.; Wu, H.; Zhu, X.; Lin, G. Effects of short-term invasion of Spartina alterniflora and the subsequent restoration of native mangroves on the soil organic carbon, nitrogen and phosphorus stock. Chemosphere 2017, 184, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Nohara, S.; Matsumoto, K.; Anzai, Y. What happens to soil chemical properties after mangrove plants colonize? Plant Soil 2011, 346, 259–273. [Google Scholar] [CrossRef]
- Yu, J.; Zhan, C.; Li, Y.; Zhou, D.; Fu, Y.; Chu, X.; Xing, Q.; Han, G.; Wang, G.; Guan, B. Distribution of carbon, nitrogen and phosphorus in coastal wetland soil related land use in the Modern Yellow River Delta. Sci. Rep. 2016, 6, 37940. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; He, Y.; Lu, J.; Chen, H.; Feng, J. Seasonal variations in soil physicochemical properties and microbial community structure influenced by Spartina alterniflora invasion and Kandelia obovata restoration. Sci. Total Environ. 2021, 797, 149213. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.X.; Jiang, Z.Y.; Wu, P.; Wang, Y.F.; Cheng, H.; Wang, Y.S.; Gu, J.D. Effect of mangrove restoration on sediment properties and bacterial community. Ecotoxicology 2021, 30, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Deng, M.; Yang, S.; Liu, W.; Wang, X.; Wang, J.; Liu, L. The coordination between leaf and fine root litter decomposition and the difference in their controlling factors. Glob. Ecol. Biogeogr. 2021, 30, 2286–2296. [Google Scholar] [CrossRef]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef]
- Kristensen, E.; Bouillon, S.; Dittmar, T.; Marchand, C. Organic carbon dynamics in mangrove ecosystems: A review. Aquat. Bot. 2008, 89, 201–219. [Google Scholar] [CrossRef]
- Yu, X.; Yang, J.; Liu, L.; Tian, Y.; Yu, Z. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland. Environ. Sci. Pollut. Res. 2015, 22, 3107–3115. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, G.; Ye, Y. Coastal vegetation invasion increases greenhouse gas emission from wetland soils but also increases soil carbon accumulation. Sci. Total Environ. 2015, 526, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Xin, K.; Yan, K.; Gao, C.; Li, Z. Carbon storage and its influencing factors in Hainan Dongzhangang mangrove wetlands. Mar. Freshw. Res. 2018, 69, 771–779. [Google Scholar] [CrossRef]
- Yu, C.; Feng, J.; Liu, K.; Wang, G.; Zhu, Y.; Chen, H.; Guan, D. Changes of ecosystem carbon stock following the plantation of exotic mangrove Sonneratia apetala in Qi’ao Island, China. Sci. Total Environ. 2020, 717, 137142. [Google Scholar] [CrossRef] [PubMed]
- Lunstrum, A.; Chen, L. Soil carbon stocks and accumulation in young mangrove forests. Soil Biol. Biochem. 2014, 75, 223–232. [Google Scholar] [CrossRef]
- Lu, W.; Yang, S.; Chen, L.; Wang, W.; Du, X.; Wang, C.; Ma, Y.; Lin, G.; Lin, G. Changes in carbon pool and stand structure of a native subtropical mangrove forest after inter-planting with exotic species Sonneratia apetala. PLoS ONE 2014, 9, e91238. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, X.; Huo, S.; Chen, X.; Wu, L.; Chen, M.; Zhou, K.; Li, Q.; Peng, L. Properties of root exudates and rhizosphere sediment of Bruguiera gymnorrhiza (L.). J. Soils Sediments 2017, 17, 266–276. [Google Scholar] [CrossRef]
- Lu, K.; Yang, Q.; Jiang, Y.; Liu, W. Changes in Temporal Dynamics and Factors Influencing the Environment of the Bacterial Community in Mangrove Rhizosphere Sediments in Hainan. Sustainability 2022, 14, 7415. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, Y.; Kang, Q.; Zhang, J. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China. Estuar. Coast. Shelf Sci. 2007, 71, 47–59. [Google Scholar] [CrossRef]
- Scharler, U.; Ulanowicz, R.E.; Fogel, M.; Wooller, M.; Jacobson-Meyers, M.; Lovelock, C.; Feller, I.; Frischer, M.; Lee, R.; McKee, K. Variable nutrient stoichiometry (carbon: Nitrogen: Phosphorus) across trophic levels determines community and ecosystem properties in an oligotrophic mangrove system. Oecologia 2015, 179, 863–876. [Google Scholar] [CrossRef]
- Shih, S.-S.; Hsieh, H.-L.; Chen, P.-H.; Chen, C.-P.; Lin, H.-J. Tradeoffs between reducing flood risks and storing carbon stocks in mangroves. Ocean Coast. Manag. 2015, 105, 116–126. [Google Scholar] [CrossRef]
- Chen, Z.; Guo, L.; Jin, B.; Wu, J.; Zheng, G. Effect of the exotic plant Spartina alterniflora on macrobenthos communities in salt marshes of the Yangtze River Estuary, China. Estuar. Coast. Shelf Sci. 2009, 82, 265–272. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).