
Optimizing China’s Afforestation Strategy: Biophysical Impacts of Afforestation with Five Locally Adapted Forest Types

Abstract
1. Introduction
2. Materials and Methods
2.1. Extracting Stable Forest and Grassland Cover
2.2. Spatial Sampling of Land Cover Data
2.3. Temperature Data and Quality Control
2.4. The Potential Impact of Forest Cover Change
3. Results
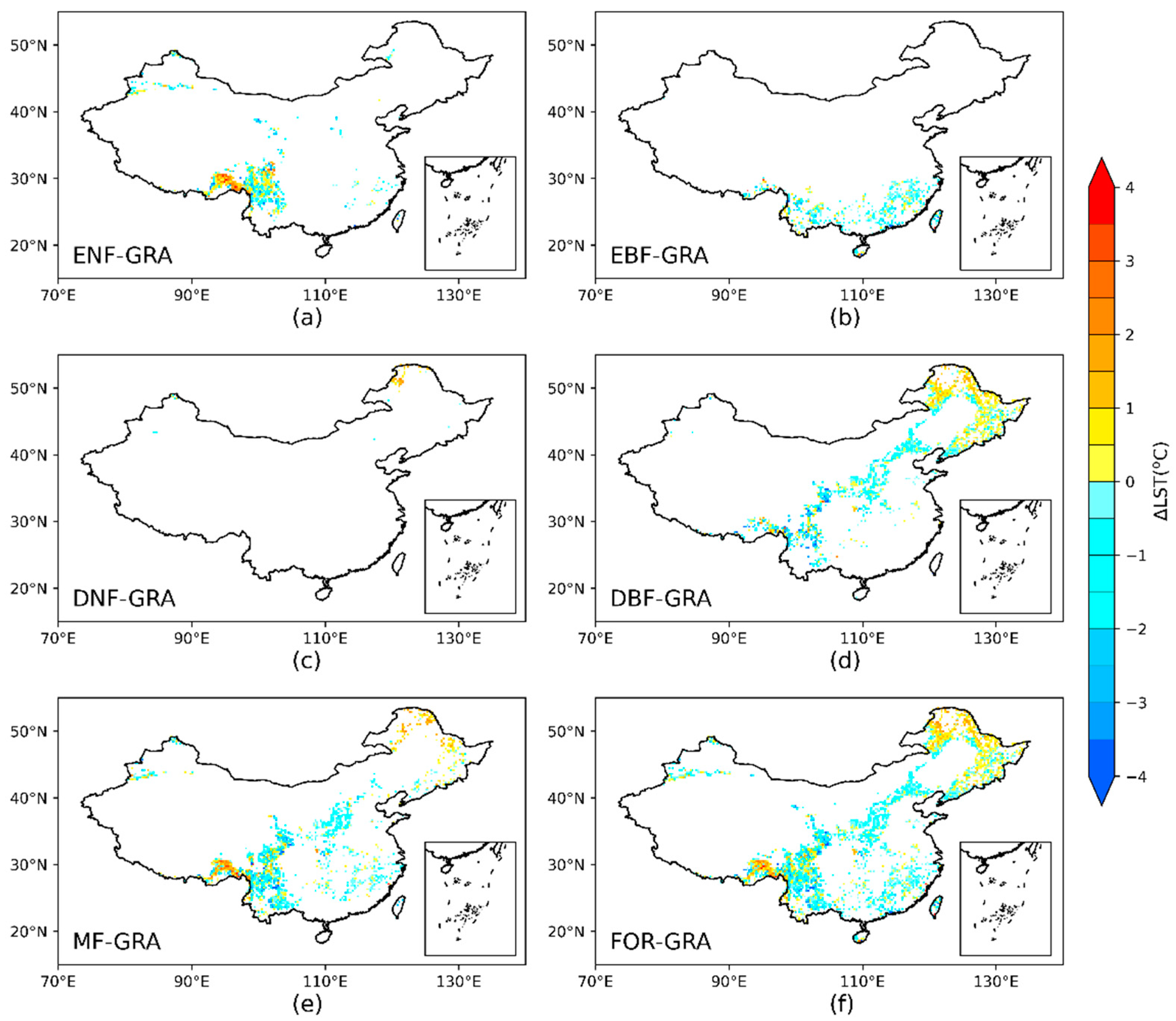
3.1. The Distribution of Potential Forest Conversion
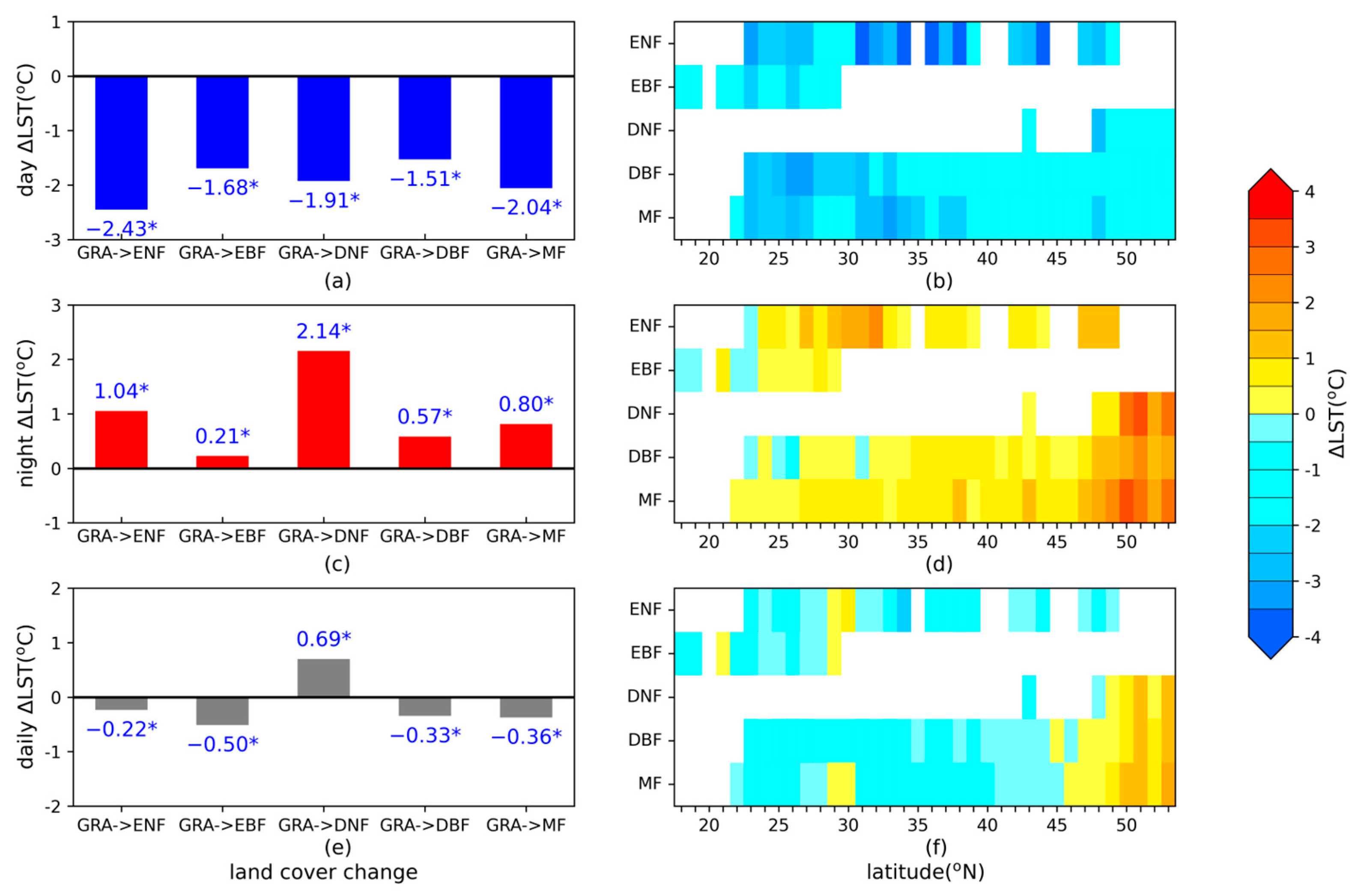
3.2. Potential Impact of Forest Conversion on ΔLST
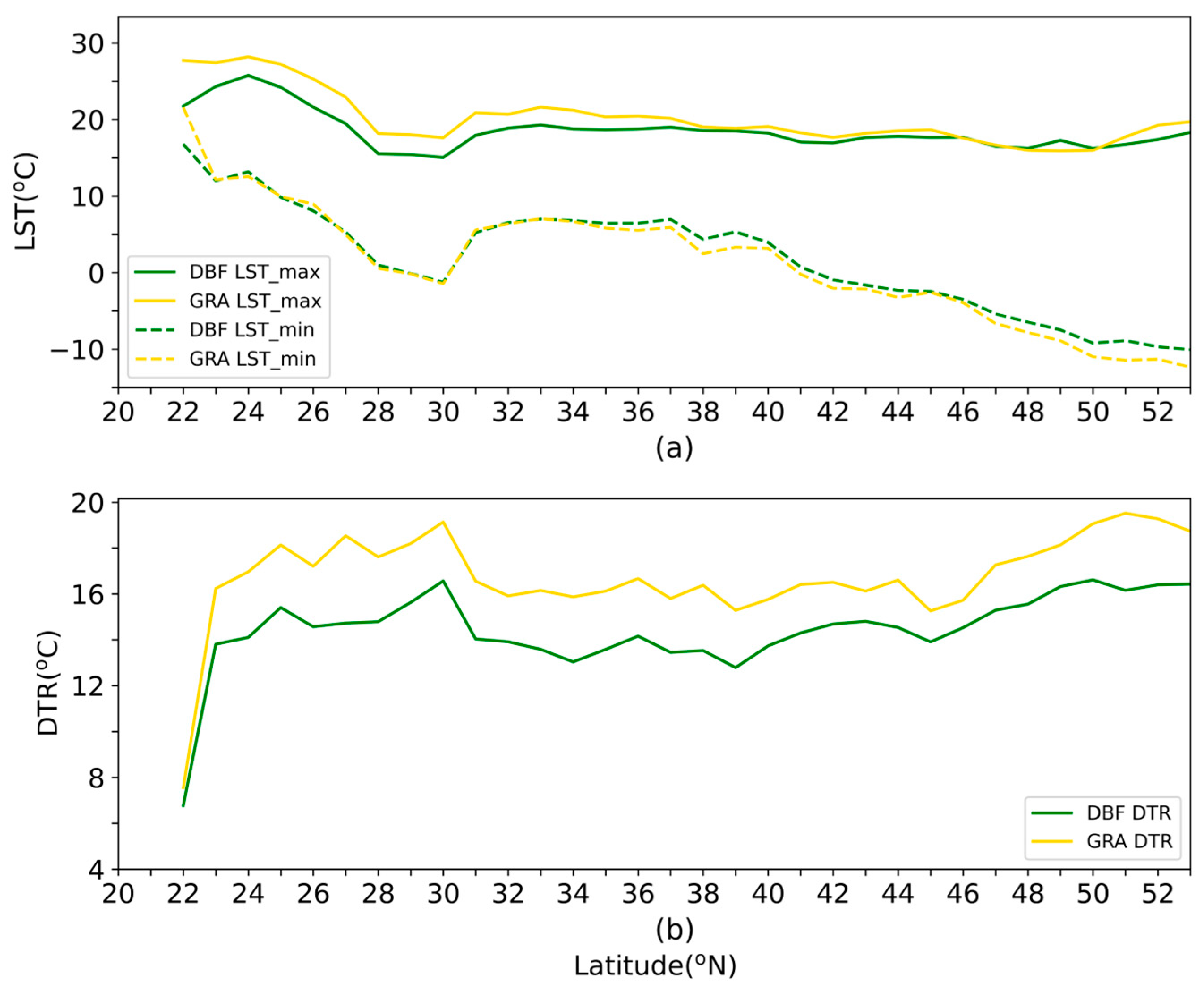
3.3. LST Patterns along Latitudinal Gradients: Perspective from DTR
4. Discussion
4.1. The Impact of Forest Types on Local Climate
4.2. Robustness of Potential Forest Change Impacts on LST
4.3. Implications and Future Work
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Deng, Z.; He, G.; Wang, H.; Zhang, X.; Lin, J.; Qi, Y.; Liang, X. Challenges and Opportunities for Carbon Neutrality in China. Nat. Rev. Earth Environ. 2022, 3, 141–155. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Canadell, J.G.; Raupach, M.R. Managing Forests for Climate Change Mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Yu, Z.; Ciais, P.; Piao, S.; Houghton, R.A.; Lu, C.; Tian, H.; Agathokleous, E.; Kattel, G.R.; Sitch, S.; Goll, D.; et al. Forest Expansion Dominates China’s Land Carbon Sink since 1980. Nat. Commun. 2022, 13, 5374. [Google Scholar] [CrossRef]
- National Development and Reform Commission; Ministry of Natural Resources of China. The Master Plan for Major Projects of National Important Ecosystem Protection and Restoration (2021–2035); National Development and Reform Commission and Ministry of Natural Resources of China: Beijing, China, 2020; pp. 10–11. Available online: https://www.ndrc.gov.cn/xxgk/zcfb/tz/202006/P020200611354032680531.pdf (accessed on 10 January 2024).
- Bright, R.M.; Davin, E.; O’Halloran, T.; Pongratz, J.; Zhao, K.; Cescatti, A. Local Temperature Response to Land Cover and Management Change Driven by Non-Radiative Processes. Nat. Clim. Chang. 2017, 7, 296–302. [Google Scholar] [CrossRef]
- Alkama, R.; Cescatti, A. Biophysical Climate Impacts of Recent Changes in Global Forest Cover. Science 2016, 351, 600–604. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Motesharrei, S.; Mu, Q.; Kalnay, E.; Li, S. Local Cooling and Warming Effects of Forests Based on Satellite Observations. Nat. Commun. 2015, 6, 6603. [Google Scholar] [CrossRef]
- Alkama, R.; Forzieri, G.; Duveiller, G.; Grassi, G.; Liang, S.; Cescatti, A. Vegetation-Based Climate Mitigation in a Warmer and Greener World. Nat. Commun. 2022, 13, 606. [Google Scholar] [CrossRef]
- Li, Y.; De Noblet-Ducoudré, N.; Davin, E.L.; Motesharrei, S.; Zeng, N.; Li, S.; Kalnay, E. The Role of Spatial Scale and Background Climate in the Latitudinal Temperature Response to Deforestation. Earth Syst. Dynam. 2016, 7, 167–181. [Google Scholar] [CrossRef]
- Lee, X.; Goulden, M.L.; Hollinger, D.Y.; Barr, A.; Black, T.A.; Bohrer, G.; Bracho, R.; Drake, B.; Goldstein, A.; Gu, L.; et al. Observed Increase in Local Cooling Effect of Deforestation at Higher Latitudes. Nature 2011, 479, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yue, C.; Zhang, Y.; Liu, D.; Piao, S. Forestation at the Right Time with the Right Species Can Generate Persistent Carbon Benefits in China. Proc. Natl. Acad. Sci. USA 2023, 120, e2304988120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Jackson, R.B. Biophysical Forcings of Land-Use Changes from Potential Forestry Activities in North America. Ecol. Monogr. 2014, 84, 329–353. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Mildrexler, D.J.; Motesharrei, S.; Mu, Q.; Kalnay, E.; Zhao, F.; Li, S.; Wang, K. Potential and Actual Impacts of Deforestation and Afforestation on Land Surface Temperature. J. Geophys. Res. Atmos. 2016, 121, 14372–14386. [Google Scholar] [CrossRef]
- Peng, S.-S.; Piao, S.; Zeng, Z.; Ciais, P.; Zhou, L.; Li, L.Z.X.; Myneni, R.B.; Yin, Y.; Zeng, H. Afforestation in China Cools Local Land Surface Temperature. Proc. Natl. Acad. Sci. USA 2014, 111, 2915–2919. [Google Scholar] [CrossRef]
- Duveiller, G.; Hooker, J.; Cescatti, A. The Mark of Vegetation Change on Earth’s Surface Energy Balance. Nat. Commun. 2018, 9, 679. [Google Scholar] [CrossRef]
- Lian, X.; Jeong, S.; Park, C.-E.; Xu, H.; Li, L.Z.X.; Wang, T.; Gentine, P.; Peñuelas, J.; Piao, S. Biophysical Impacts of Northern Vegetation Changes on Seasonal Warming Patterns. Nat. Commun. 2022, 13, 3925. [Google Scholar] [CrossRef]
- Yang, J.; Wu, Q.; Dakhil, M.A.; Halmy, M.W.A.; Bedair, H.; Fouad, M.S. Towards Forest Conservation Planning: How Temperature Fluctuations Determine the Potential Distribution and Extinction Risk of Cupressus funebris Conifer Trees in China. Forests 2023, 14, 2234. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Xu, L.; Kiang, N. How Plant Functional-Type, Weather, Seasonal Drought, and Soil Physical Properties Alter Water and Energy Fluxes of an Oak–Grass Savanna and an Annual Grassland. Agric. For. Meteorol. 2004, 123, 13–39. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P.M. Water and Energy Balance. In Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer: New York, NY, USA, 2012; pp. 94–100. [Google Scholar]
- Mahmood, R.; Pielke, R.A.; Hubbard, K.G.; Niyogi, D.; Bonan, G.; Lawrence, P.; McNider, R.; McAlpine, C.; Etter, A.; Gameda, S.; et al. Impacts of Land Use/Land Cover Change on Climate and Future Research Priorities. Bull. Am. Meteorol. Soc. 2010, 91, 37–46. [Google Scholar] [CrossRef]
- Pielke, R.A., Sr.; Pitman, A.; Niyogi, D.; Mahmood, R.; McAlpine, C.; Hossain, F.; Goldewijk, K.K.; Nair, U.; Betts, R.; Fall, S.; et al. Land Use/Land Cover Changes and Climate: Modeling Analysis and Observational Evidence. WIREs Clim. Change 2011, 2, 828–850. [Google Scholar] [CrossRef]
- Chen, C.; Li, D.; Li, Y.; Piao, S.; Wang, X.; Huang, M.; Gentine, P.; Nemani, R.R.; Myneni, R.B. Biophysical Impacts of Earth Greening Largely Controlled by Aerodynamic Resistance. Sci. Adv. 2020, 6, eabb1981. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.-L.; Wu, H.; Zhou, C.; Liu, X.; Leng, P.; Yang, P.; Wu, W.; Tang, R.; Shang, G.-F.; et al. Biophysical Impacts of Earth Greening Can Substantially Mitigate Regional Land Surface Temperature Warming. Nat. Commun. 2023, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Sulla-Menashe, D.; Friedl, M. MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V061 [MCD12Q1]. NASA EOSDIS Land Processes Distributed Active Archive Center. 2020. Available online: https://lpdaac.usgs.gov/products/mcd12q1v061/ (accessed on 10 January 2024).
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 Global Land Cover: Algorithm Refinements and Characterization of New Datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Wan, Z.M. MODIS/Terra(Aqua) Land Surface Temperature/Emissivity 8-Day L3 Global 500m SIN Grid V061 [MO(Y)D11A2]. NASA EOSDIS Land Processes Distributed Active Archive Center. 2019. Available online: https://lpdaac.usgs.gov/products/mod11a2v061/ (accessed on 10 January 2024).
- The Shuttle Radar Topography Mission (SRTM) Collection. SRTM Global 30 Arc Second Elevation [SRTMGL30v021]. NASA EOSDIS Land Processes Distributed Active Archive Center. 2019. Available online: https://lpdaac.usgs.gov/products/srtmgl30v021/ (accessed on 10 January 2024).
- Bonan, G.B. Surface Energy Fluxes. In Ecological Climatology: Principles and Applications, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 193–205. [Google Scholar]
- Shen, M.; Piao, S.; Jeong, S.-J.; Zhou, L.; Zeng, Z.; Ciais, P.; Chen, D.; Huang, M.; Jin, C.-S.; Li, L.Z.X.; et al. Evaporative Cooling over the Tibetan Plateau Induced by Vegetation Growth. Proc. Natl. Acad. Sci. USA 2015, 112, 9299–9304. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Knyazikhin, Y.; Wang, W.; Deering, D.W.; Stenberg, P.; Shabanov, N.; Tan, B.; Myneni, R.B. Stochastic Transport Theory for Investigating the Three-Dimensional Canopy Structure from Space Measurements. Remote Sens. Environ. 2008, 112, 35–50. [Google Scholar] [CrossRef]
- Waring, R.H.; Running, S.W. Water Cycles. In Forest Ecosystems: Analysis at Multiple Scales, 3rd ed.; Academic Press: San Diego, CA, USA, 2007; pp. 50–52. [Google Scholar]
- Bellasio, R.; Maffeis, G.; Scire, J.S.; Longoni, M.G.; Bianconi, R.; Quaranta, N. Algorithms to Account for Topographic Shading Effects and Surface Temperature Dependence on Terrain Elevation in Diagnostic Meteorological Models. Bound. Layer Meteorol. 2005, 114, 595–614. [Google Scholar] [CrossRef]
- Chen, X.; Su, Z.; Ma, Y.; Yang, K.; Wang, B. Estimation of Surface Energy Fluxes under Complex Terrain of Mt. Qomolangma over the Tibetan Plateau. Hydrol. Earth Syst. Sci. 2013, 17, 1607–1618. [Google Scholar] [CrossRef]
- Zhang, M.; Lee, X.; Yu, G.; Han, S.; Wang, H.; Yan, J.; Zhang, Y.; Li, Y.; Ohta, T.; Hirano, T.; et al. Response of Surface Air Temperature to Small-Scale Land Clearing across Latitudes. Environ. Res. Lett. 2014, 9, 034002. [Google Scholar] [CrossRef]
- Essery, R. Large-Scale Simulations of Snow Albedo Masking by Forests. Geophys. Res. Lett. 2013, 40, 5521–5525. [Google Scholar] [CrossRef]
- Eugster, W.; Rouse, W.; Pielke, R.A., Sr.; McFadden, J.P.; Baldocchi, D.D.; Kittel, T.G.F.; Chapin, F.S.; Liston, G.E.; Vidale, P.L.; Vaganov, E.A.; et al. Land–Atmosphere Energy Exchange in Arctic Tundra and Boreal Forest: Available Data and Feedbacks to Climate. Glob. Chang. Biol. 2000, 6, 84–115. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, D.Y.; Ollinger, S.V.; Richardson, A.D.; Meyers, T.P.; Dail, D.B.; Martin, M.E.; Scott, N.A.; Arkebauer, T.J.; Baldocchi, D.D.; Clark, K.L.; et al. Albedo Estimates for Land Surface Models and Support for a New Paradigm Based on Foliage Nitrogen Concentration. Glob. Chang. Biol. 2010, 16, 696–710. [Google Scholar] [CrossRef]
- Jarvis, P.G.; McNaughton, K.G. Stomatal Control of Transpiration: Scaling Up from Leaf to Region. In Advances in Ecological Research; MacFadyen, A., Ford, E.D., Eds.; Academic Press: Cambridge, MA, USA, 1986; Volume 15, pp. 1–49. [Google Scholar]
- Kelliher, F.M.; Jackson, R. The Physical Environment: A New Zealand Perspective. In Evaporation and the Water Balance; Sturman, A., Spronken-Smith, R., Eds.; Oxford University Press: Melbourne, Australia, 2001; pp. 206–217. [Google Scholar]
- Wilson, K.B.; Baldocchi, D.D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Dolman, H.; Falge, E.; Field, C.; Goldstein, A.; Granier, A.; et al. Energy Partitioning between Latent and Sensible Heat Flux during the Warm Season at FLUXNET Sites. Water Resour. Res. 2002, 38, 30–31. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.B.; Mooney, H.A.; Sala, O.E.; Schulze, E.-D. Maximum Rooting Depth of Vegetation Types at the Global Scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Ma, W.; Jia, G.; Zhang, A. Multiple Satellite-Based Analysis Reveals Complex Climate Effects of Temperate Forests and Related Energy Budget. J. Geophys. Res. Atmos. 2017, 122, 3806–3820. [Google Scholar] [CrossRef]
- Wickham, J.D.; Wade, T.G.; Riitters, K.H. Empirical Analysis of the Influence of Forest Extent on Annual and Seasonal Surface Temperatures for the Continental United States. Glob. Ecol. Biogeogr. 2013, 22, 620–629. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ΔLST (°C) | |||||||
| Type\Pixel | 1 | 9 | 18 | 27 | 36 | 45 | 54 |
| ENF vs. GRA | −0.22 | 0.13 | 0.21 | 0.21 | 0.28 | 0.29 | 0.29 |
| EBF vs. GRA | −0.50 | −0.46 | −0.83 | −0.72 | −0.78 | −0.78 | |
| DNF vs. GRA | 0.69 | 0.62 | 0.67 | 1.38 | |||
| DBF vs. GRA | −0.33 | −0.47 | −0.60 | −0.63 | −0.54 | −0.49 | −0.50 |
| MF vs. GRA | −0.36 | −0.44 | −0.38 | −0.42 | −0.47 | −0.47 | −0.38 |
| Selected Sample Numbers | |||||||
| Type | 1 | 9 | 18 | 27 | 36 | 45 | 54 |
| ENF vs. GRA | 696 | 336 | 244 | 190 | 150 | 111 | 85 |
| EBF vs. GRA | 563 | 35 | 12 | 6 | 2 | 2 | 0 |
| DNF vs. GRA | 67 | 5 | 3 | 1 | 0 | 0 | 0 |
| DBF vs. GRA | 1623 | 481 | 262 | 181 | 124 | 85 | 62 |
| MF vs. GRA | 1550 | 519 | 330 | 224 | 164 | 126 | 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, W.; Wang, Y. Optimizing China’s Afforestation Strategy: Biophysical Impacts of Afforestation with Five Locally Adapted Forest Types. Forests 2024, 15, 182. https://doi.org/10.3390/f15010182
Ma W, Wang Y. Optimizing China’s Afforestation Strategy: Biophysical Impacts of Afforestation with Five Locally Adapted Forest Types. Forests. 2024; 15(1):182. https://doi.org/10.3390/f15010182
Chicago/Turabian StyleMa, Wei, and Yue Wang. 2024. "Optimizing China’s Afforestation Strategy: Biophysical Impacts of Afforestation with Five Locally Adapted Forest Types" Forests 15, no. 1: 182. https://doi.org/10.3390/f15010182
APA StyleMa, W., & Wang, Y. (2024). Optimizing China’s Afforestation Strategy: Biophysical Impacts of Afforestation with Five Locally Adapted Forest Types. Forests, 15(1), 182. https://doi.org/10.3390/f15010182