
Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Acorn Collection
2.2. Morphological Traits of the Acorn and Seed
2.3. Seed Pretreatment and Germination Test
2.4. Calculation of Seed Germination Indices
2.5. Statistical Analyses
3. Results
3.1. Morphological Characteristics of the Acorn and Seed Germination
3.2. Seed Germination Traits
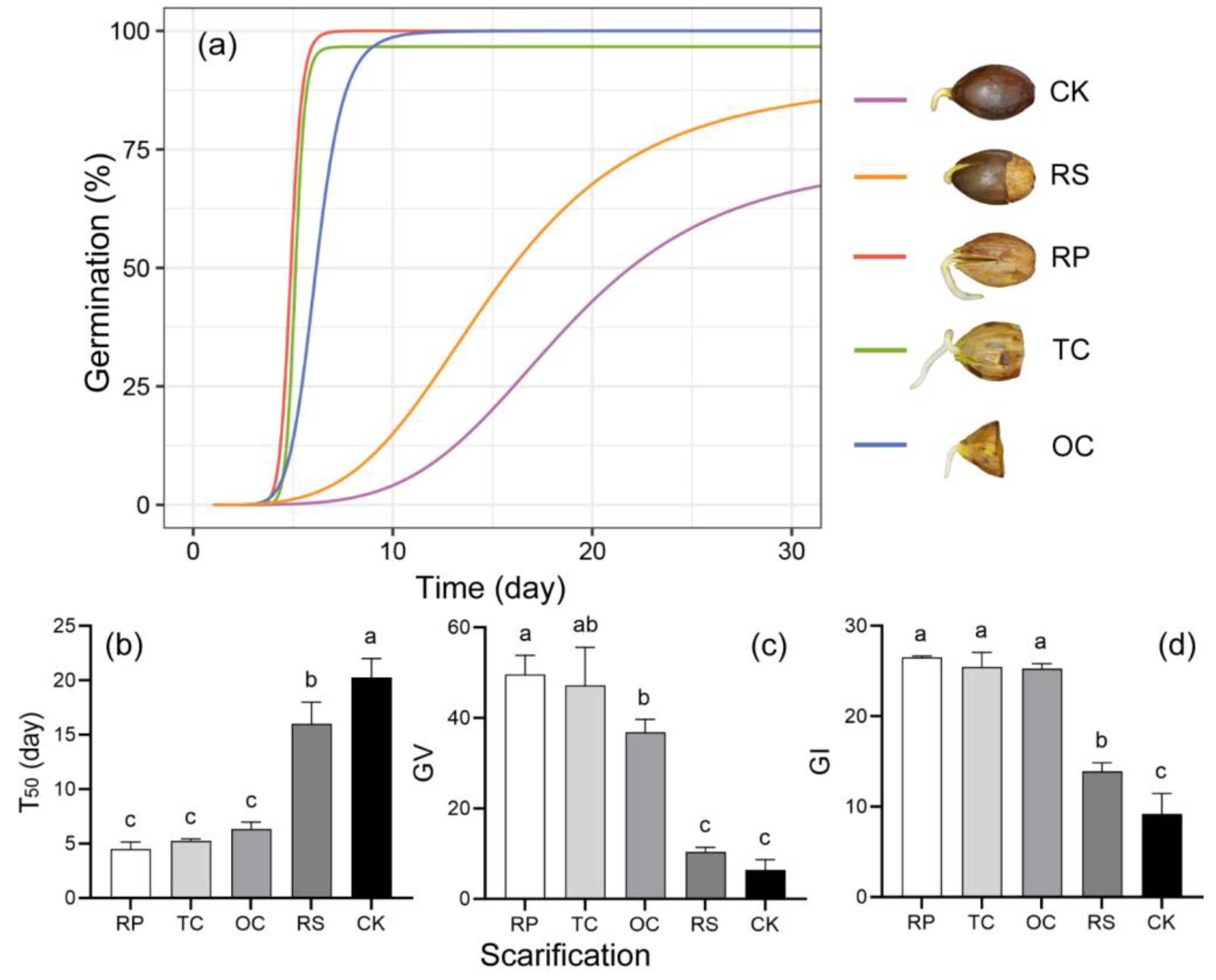
3.2.1. Scarification and Germination
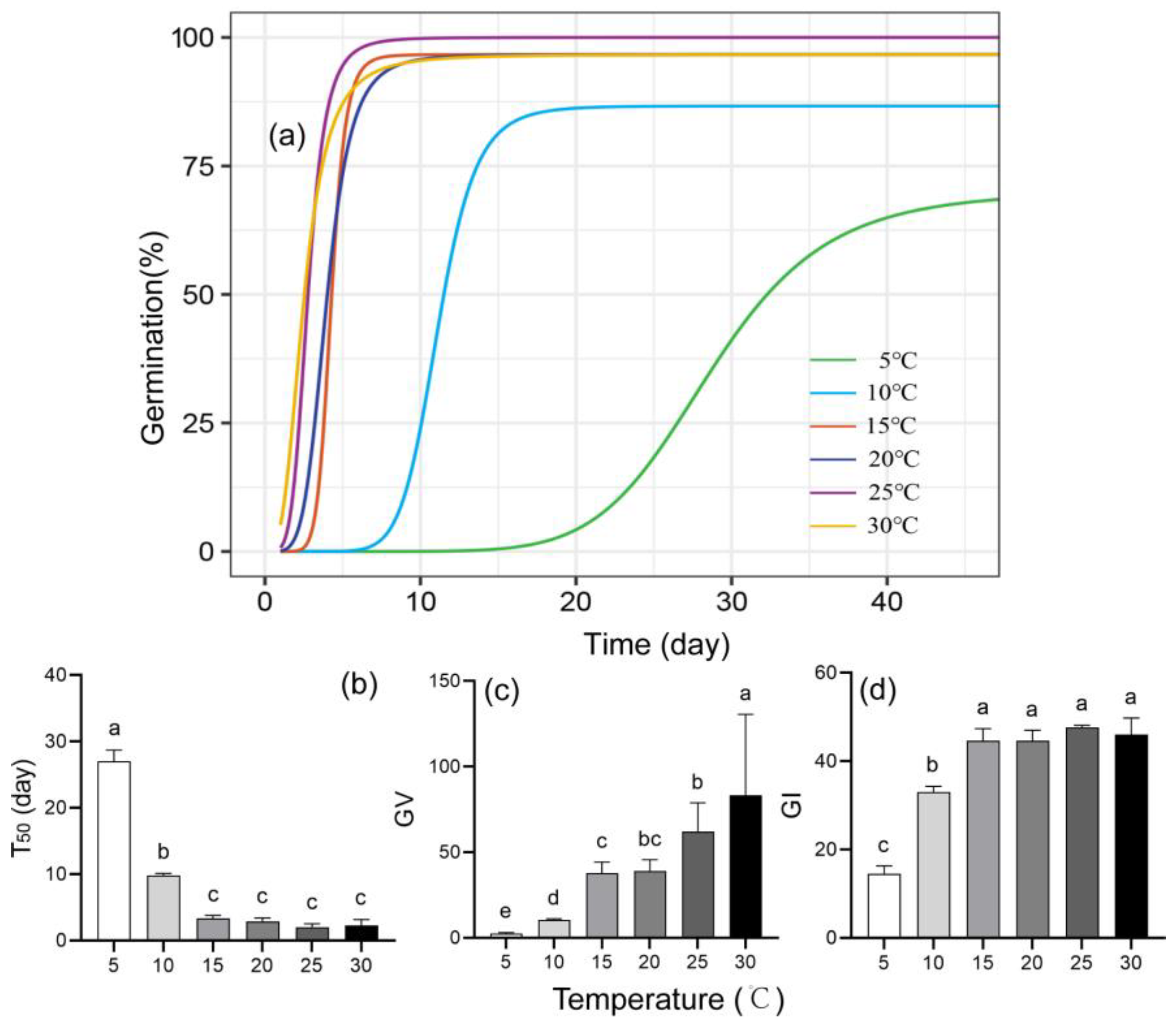
3.2.2. Temperature and Germination
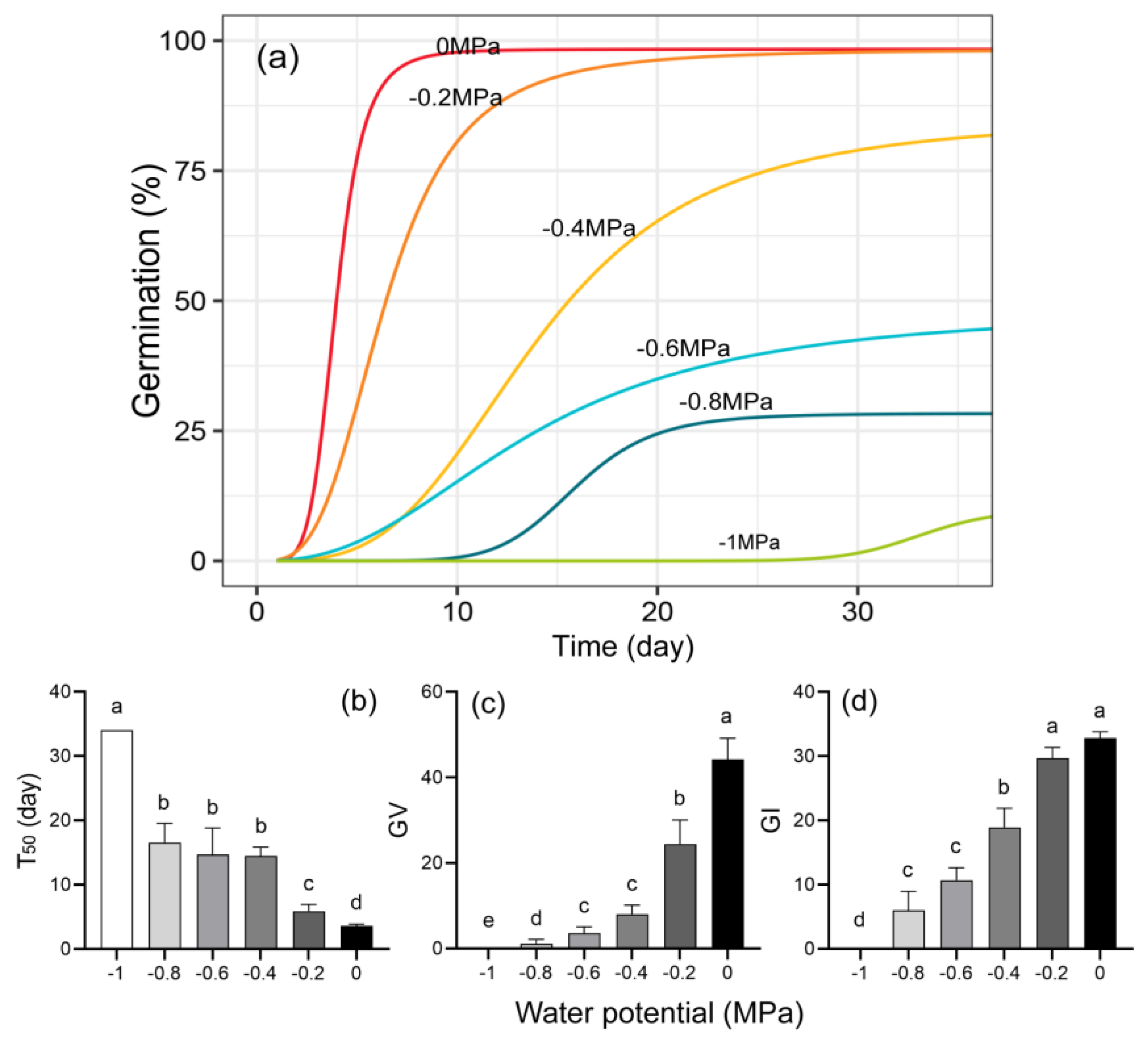
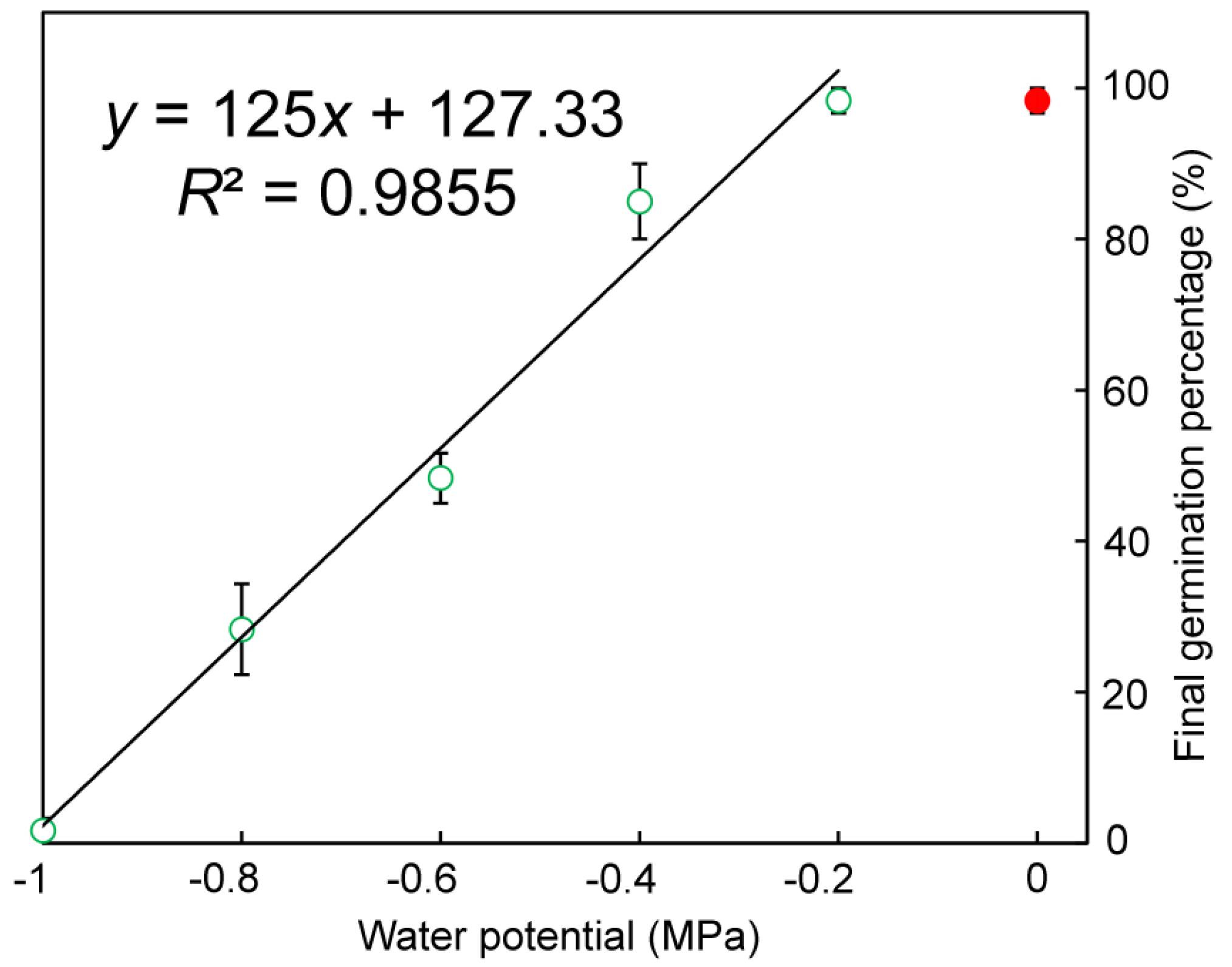
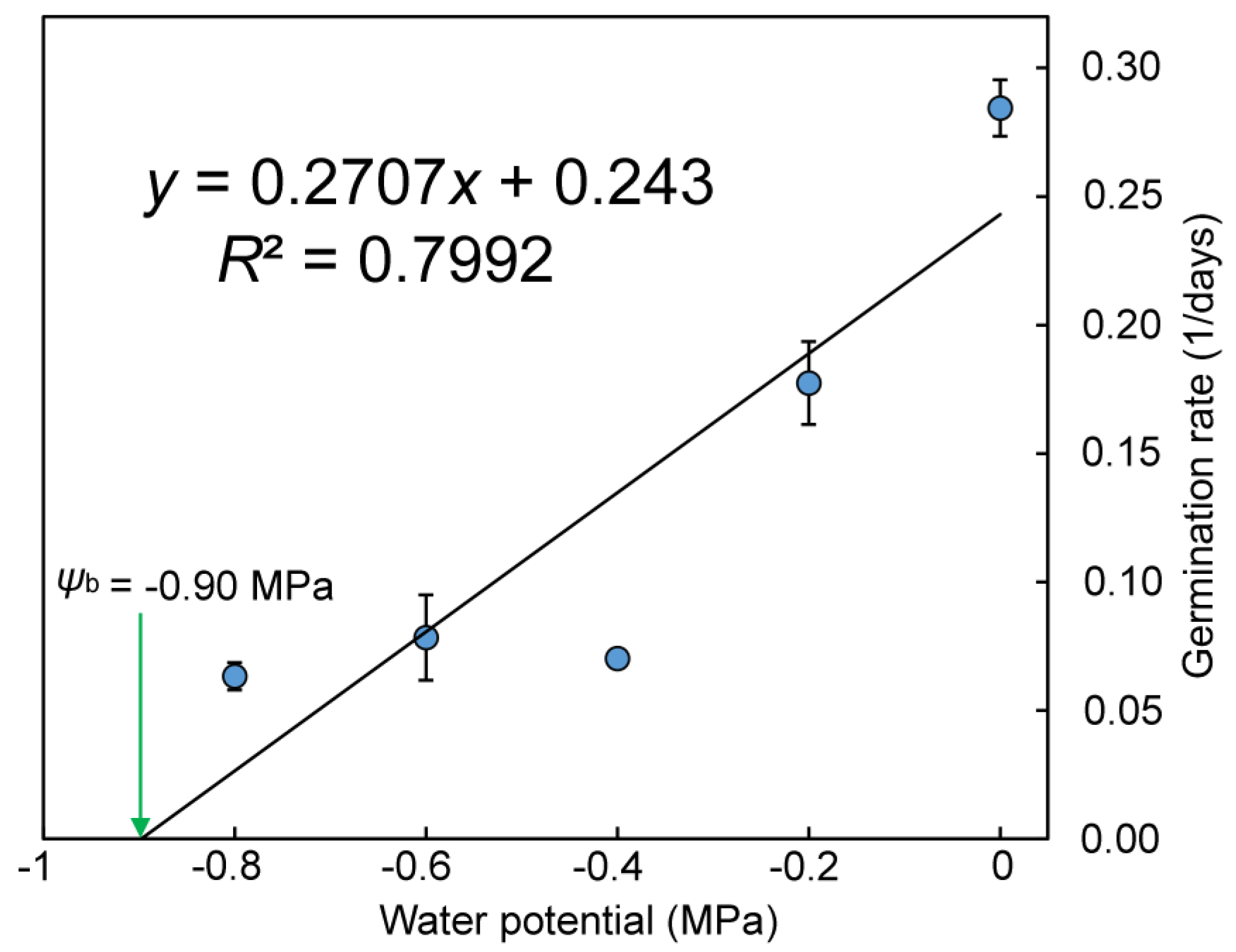
3.2.3. Water Potential and Germination
4. Discussion
4.1. Impacts of Pericarp and Cotyledon Removal on the Germination of Quercus marlipoensis
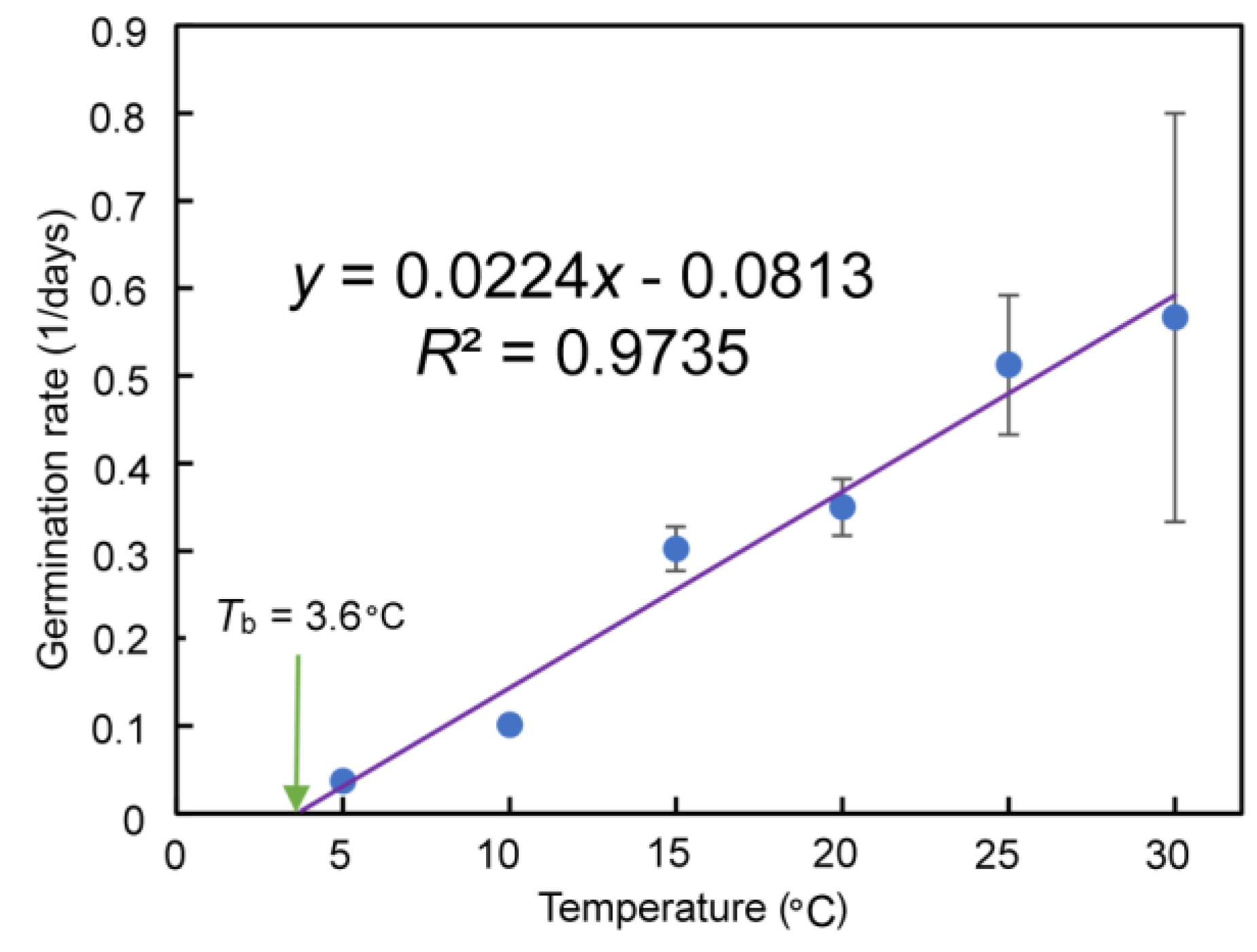
4.2. Effect of Temperature on Seed Germination and the Base Temperature
4.3. Effect of Water Potential on Seed Germination
4.4. Conservation Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kappelle, M.; Burley, S.; Evans, J.; Youngquist, J. Tropical montane forests. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Amsterdam, The Netherland, 2004; Volume 4, pp. 1782–1793. [Google Scholar] [CrossRef]
- Ray, D.K. Tropical montane cloud forests. In Climate Vulnerability; Pielke, R.A., Ed.; Academic Press: New York, NY, USA, 2013; Volume 5, pp. 79–85. [Google Scholar] [CrossRef]
- van de Weg, M.J.; Meir, P.; Williams, M.; Girardin, C.; Malhi, Y.; Silva-Espejo, J.; Grace, J. Gross primary productivity of a high elevation tropical montane cloud forest. Ecosystems 2014, 17, 751–764. [Google Scholar] [CrossRef]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Körner, O.; Rezaei Nejad, A.; Delis, C.; Bouranis, D.; Koubouris, G.; Kambourakis, E.; et al. Stomatal behavior following mid or long–term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Eller, C.B.; Meireles, L.D.; Sitch, S.; Burgess, S.S.O.; Oliveira, R.S. How climate shapes the functioning of tropical montane cloud forests. Curr. For. Rep. 2020, 6, 97–114. [Google Scholar] [CrossRef]
- Shi, J.P.; Zhu, H. Tree species composition and diversity of tropical mountain cloud forest in the Yunnan, southwestern China. Ecol. Res. 2009, 24, 83–92. [Google Scholar] [CrossRef]
- Tang, C.Q. Evergreen sclerophyllous Quercus communities. In The Subtropical Vegetation of Southwestern China; Tang, C.Q., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2015; Volume 11, pp. 113–125. [Google Scholar] [CrossRef]
- Hu, H.H. Additional notes on the Fagaceae of Yunnan, II. Acta Phytotax. Sin. 1951, 2, 139. [Google Scholar]
- Deng, M.; Jiang, X.L.; Song, Y.G.; Coombes, A.; Yang, X.R.; Xiong, Y.S.; Li, Q.S. Leaf epidermal features of Quercus group Ilex (Fagaceae) and their application to species identification. Rev. Palaeobot. Palynol. 2017, 237, 10–36. [Google Scholar] [CrossRef]
- Carrero, C.; Strijk, J.S. Quercus marlipoensis. The IUCN Red List of Threatened Species 2020: E.T191481A1985007. Available online: https://www.iucnredlist.org/species/191481/1985007 (accessed on 20 January 2024).
- Sun, W.B. List of Yunnan Protected Plant Species with Extremely Small Populations (2021); Yunnan Science and Technology Press: Kunming, China, 2021. [Google Scholar]
- Iralu, V.; Barbhuyan, H.S.A.; Upadhaya, K. Ecology of seed germination in threatened trees: A review. Energy Ecol. Environ. 2019, 4, 189–210. [Google Scholar] [CrossRef]
- Wade, E.M.; Nadarajan, J.; Yang, X.Y.; Ballesteros, D.; Sun, W.B.; Pritchard, H.W. Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective. Plant Divers. 2016, 38, 209–220. [Google Scholar] [CrossRef]
- Yi, X.F.; Yang, Y.Q. Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena. Plant Ecol. 2010, 209, 291–300. [Google Scholar] [CrossRef]
- Tripathi, R.S.; Khan, M.L. Effects of seed weight and microsite characteristics on germination and seedling fitness in two species of Quercus in a subtropical wet hill forest. Oikos 1990, 57, 289–296. [Google Scholar] [CrossRef]
- Leishman, M.R.; Westoby, M. The role of seed size in seedling establishment in dry soil conditions-experimental evidence from semi-arid species. J. Ecol. 1994, 82, 249–258. [Google Scholar] [CrossRef]
- Xiao, Z.; Harris, M.K.; Zhang, Z. Acorn defenses to herbivory from insects: Implications for the joint evolution of resistance, tolerance and escape. For. Ecol. Manag. 2007, 238, 302–308. [Google Scholar] [CrossRef]
- Yi, X.F.; Zhang, Z.B. Influence of insect-infested cotyledons on early seedling growth of Mongolian oak, Quercus mongolica. Photosynthetica 2008, 46, 139–142. [Google Scholar] [CrossRef]
- Montes-Hernández, B.; López-Barrera, F. Seedling establishment of Quercus insignis: A critically endangered oak tree species in southern Mexico. For. Ecol. Manag. 2013, 310, 927–934. [Google Scholar] [CrossRef]
- García-Hernández, M.d.l.Á.; López-Barrera, F.; Vásquez-Reyes, V.M. Microhabitat affects acorn removal in three sympatric and endangered neotropical oak species. Ecol. Res. 2016, 31, 343–351. [Google Scholar] [CrossRef]
- Cao, L.; Guo, C.; Chen, J. Fluctuation in seed abundance has contrasting effects on the fate of seeds from two rapidly germinating tree species in an Asian tropical forest. Integr. Zool. 2017, 12, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Hopper, G.; Smith, D.; Parrish, D. Germination and seedling growth of northern red oak–effects of stratification and pericarp removal. For. Sci. 1985, 31, 31–39. [Google Scholar]
- Liu, Y.; Hou, L.Y.; Li, Q.M. Effects of different mechanical treatments on Quercus variabilis, Q. wutaishanica and Q. robur acorn germination. iForest 2015, 8, 728–734. [Google Scholar] [CrossRef]
- Armstrong, D.P.; Westoby, M. Seedlings from large seeds tolerated defoliation better: A test using phylogenetically independent contrasts. Ecology 1993, 74, 1092–1100. [Google Scholar] [CrossRef]
- Dalling, J.W.; Harms, K.E.; Aizprúa, R. Seed damage tolerance and seedling resprouting ability of Prioria copaifera in Panamá. J. Trop. Ecol. 1997, 13, 481–490. [Google Scholar] [CrossRef]
- Branco, M.; Branco, C.; Merouani, H.; Almeida, M.H. Germination success, survival and seedling vigour of Quercus suber acorns in relation to insect damage. For. Ecol. Manag. 2002, 166, 159–164. [Google Scholar] [CrossRef]
- Giertych, M.J.; Suszka, J. Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing. Ann. For. Sci. 2011, 68, 433–442. [Google Scholar] [CrossRef]
- Fukumoto, H.; Kajimura, H. Effects of insect predation on hypocotyl survival and germination success of mature Quercus variabilis acorns. J. For. Res. 2000, 5, 31–34. [Google Scholar] [CrossRef]
- Dürr, C.; Dickie, J.B.; Yang, X.Y.; Pritchard, H.W. Ranges of critical temperature and water potential values for the germination of species worldwide: Contribution to a seed trait database. Agric. For. Meteorol. 2015, 200, 222–232. [Google Scholar] [CrossRef]
- Schroeder, W.; Walker, D. Effects of moisture content and storage temperatures on germination of Quercus macrocarpa acorns. J. Environ. Hort. 1987, 5, 22–24. [Google Scholar] [CrossRef]
- Gosling, P.G. The effect of drying Quercus robur acorns to different moisture contents, followed by storage, either with or without imbibition. Forestry 1989, 62, 41–50. [Google Scholar] [CrossRef]
- Connor, K.F.; Sowa, S. Effects of desiccation on the physiology and biochemistry of Quercus alba acorns. Tree Physiol. 2003, 23, 1147–1152. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.; McNabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial regeneration of major oak (Quercus) species in the eastern United States—A review of the literature. For. Sci. 2008, 54, 77–106. [Google Scholar] [CrossRef]
- Mechergui, T.; Pardos, M.; Boussaidi, N.; Jacobs, D.F.; Catry, F.X. Problems and solutions to cork oak (Quercus suber L.) regeneration: A review. iForest 2023, 16, 10–22. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Archidona-Yuste, A.; Abril, N.; Gil-Serrano, A.M.; Meijón, M.; Jorrín-Novo, J.V. Germination and early seedling development in Quercus ilex recalcitrant and non-dormant seeds: Targeted transcriptional, hormonal, and sugar analysis. Front. Plant Sci. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. 2007, 101, 213–228. [Google Scholar] [CrossRef]
- Bonner, F.; Vozzo, J. Seed Biology and Technology of Quercus; US Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1987. [Google Scholar]
- Bonner, F.T. Water uptake and germination of red oak acorns. Bot. Gaz. 1968, 129, 83–85. [Google Scholar] [CrossRef]
- Amen, R.D. A model of seed dormancy. Bot. Rev. 1968, 34, 1–31. [Google Scholar] [CrossRef]
- Jones, L.; Brown, C.L. Cause of slow germination in Cherrybark and Northern Red Oak. Proc. Assoc. Off. Seed Anal. 1966, 56, 82–88. [Google Scholar]
- Abrams, M.D. Fire and the development of oak forests. Bioscience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Kremer, A.; Hipp, A.L. Oaks: An evolutionary success story. New Phytol. 2020, 226, 987–1011. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Daws, M.I.; Peng, L.L. Climate drives patterns of seed traits in Quercus species across China. New Phytol. 2022, 234, 1629–1638. [Google Scholar] [CrossRef]
- Löf, M.; Castro, J.; Engman, M.; Leverkus, A.B.; Madsen, P.; Reque, J.A.; Villalobos, A.; Gardiner, E.S. Tamm review: Direct seeding to restore oak (Quercus spp.) forests and woodlands. For. Ecol. Manag. 2019, 448, 474–489. [Google Scholar] [CrossRef]
- Amimi, N.; Dussert, S.; Vaissayre, V.; Ghouil, H.; Doulbeau, S.; Costantini, C.; Ammari, Y.; Joët, T. Variation in seed traits among mediterranean oaks in Tunisia and their ecological significance. Ann. Bot. 2020, 125, 891–904. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules for Seed Testing (2010); ISTA: Zürich, Switzerland, 2009. [Google Scholar]
- Tilki, F.; Alptekin, C.U. Variation in acorn characteristics in three provenances of Quercus aucheri Jaub. et Spach and provenance, temperature and storage effects on acorn germination. Seed Sci. Technol. 2005, 33, 441–447. [Google Scholar] [CrossRef]
- Peterson, J.K. Mechanisms involved in delayed germination of Quercus nigra L. seeds. Ann. Bot. 1983, 52, 81–92. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Aravind, J.; Vimala Devi, S.; Radhamani, J.; Jacob, S.R.; Srinivasan, K. Germinationmetrics: Seed Germination Indices and Curve Fitting, R Package Version 0.1.8. Available online: https://github.com/aravind-j/germinationmetricshttps://cran.r-project.org/package=germinationmetrics (accessed on 19 October 2023).
- Daws, M.I.; Crabtree, L.M.; Dalling, J.W.; Mullins, C.E.; Burslem, D.F.R.P. Germination responses to water potential in neotropical pioneers suggest large-seeded species take more risks. Ann. Bot. 2008, 102, 945–951. [Google Scholar] [CrossRef]
- Yan, X.F.; Zhou, L.B.; Si, B.B.; Sun, Y.; Gao, Y.F.; Wang, R.X. Stress effects of simulated drought by polyethylene glycol on the germination of Caragana korshinskii Kom. seeds under different temperature conditions. Acta Ecol. Sin. 2016, 36, 1989–1996. [Google Scholar] [CrossRef]
- Li, Z.X.; Pei, X.N.; Yin, S.P.; Lang, X.B.; Zhao, X.Y.; Qu, G.Z. Plant hormone treatments to alleviate the effects of salt stress on germination of Betula Platyphylla seeds. J. For. Res. 2019, 30, 779–787. [Google Scholar] [CrossRef]
- Arnold, C.Y. The determination and significance of the base temperature in a linear heat unit system. Proc. Am. Soc. Hortic. Sci. 1959, 74, 430–445. [Google Scholar]
- Guillemin, J.P.; Gardarin, A.; Granger, S.; Reibel, C.; Munier-Jolain, N.; Colbach, N. Assessing potential germination period of weeds with base temperatures and base water potentials. Weed Res. 2013, 53, 76–87. [Google Scholar] [CrossRef]
- Yi, X.F.; Zhang, M.M.; Bartlow, A.W.; Dong, Z. Incorporating cache management behavior into seed dispersal: The effect of pericarp removal on acorn germination. PLoS ONE 2014, 9, e92544. [Google Scholar] [CrossRef]
- Aniszewska, M.; Błuszkowska, U.; Zychowicz, W.; Brzózko, J. Impact of mechanical treatment of pedunculate oak (Quercus robur L.) seeds on germination time and seedling quality. J. For. Res. 2020, 25, 420–425. [Google Scholar] [CrossRef]
- Rawat, D.C.S.; Thapliyal, P.; Nautiyal, A.R. Pericarp delays germination in Quercus glauca seeds. J. Trop. For. Sci. 1998, 10, 472–477. [Google Scholar]
- Liu, Y.; Liu, G.Q.; Li, Q.M.; Liu, Y.; Hou, L.Y.; Li, G.L. Influence of pericarp, cotyledon and inhibitory substances on sharp tooth oak (Quercus aliena var. acuteserrata) germination. PLoS ONE 2012, 7, e47682. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.G.; Yi, X.F.; Yang, Y.Q.; Liu, W.J. Acorn germination and seedling survival of Q. variabilis: Effects of cotyledon excision. Ann. For. Sci. 2010, 67, 711. [Google Scholar] [CrossRef]
- Yi, X.F.; Bartlow, A.W.; Curtis, R.; Agosta, S.J.; Steele, M.A. Responses of seedling growth and survival to post-germination cotyledon removal: An investigation among seven oak species. J. Ecol. 2019, 107, 1817–1827. [Google Scholar] [CrossRef]
- Yi, X.F.; Wang, Z.Y.; Liu, C.Q.; Liu, G.Q.; Zhang, M.M. Acorn cotyledons are larger than their seedlings’ need: Evidence from artificial cutting experiments. Sci. Rep. 2015, 5, 8112. [Google Scholar] [CrossRef]
- Tilki, F. Influence of acorn size and storage duration on moisture content, germination and survival of Quercus petraea (Mattuschka). J. Environ. Biol. 2010, 31, 325–328. [Google Scholar] [PubMed]
- Elwell, A.L.; Gronwall, D.S.; Miller, N.D.; Spalding, E.P.; Durham Brooks, T.L. Separating parental environment from seed size effects on next generation growth and development in Arabidopsis. Plant Cell Environ. 2011, 34, 291–301. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Manger, K.R. Quantal response of fruit and seed germination rate in Quercus robur L. and Castanea sativa Mill, to constant temperatures and photon dose. J. Exp. Bot. 1990, 41, 1549–1557. [Google Scholar] [CrossRef]
- Xia, K.; Daws, M.I.; Zhou, Z.K.; Pritchard, H.W. Habitat-linked temperature requirements for fruit germination in Quercus species: A comparative study of Quercus subgenus Cyclobalanopsis (Asian evergreen oaks) and Quercus subgenus Quercus. S. Afr. J. Bot. 2015, 100, 108–113. [Google Scholar] [CrossRef]
- Muñiz-Castro, M.A.; Williams-Linera, G.; Benítez-Malvido, J. Restoring montane cloud forest: Establishment of three Fagaceae species in the old fields of central Veracruz, Mexico. Restor. Ecol. 2015, 23, 26–33. [Google Scholar] [CrossRef]
- Valdés-Rodríguez, O.A.; Cruz, Y.G.-D.L.; Frey, B.R. Survival and growth of three endangered oak species in a Mexican montane cloud forest. Ann. For. Sci. 2017, 60, 89–100. [Google Scholar] [CrossRef]
- Pandey, R.; Bargali, K.; Bargali, S.S. Does seed size affect water stress tolerance in Quercus Leucotrichophora a. Camus at germination and early seedling growth stage? Biodivers. Int. J. 2017, 1, 24–30. [Google Scholar] [CrossRef]
- Tozer, M.G.; Ooi, M.K.J. Humidity-regulated dormancy onset in the Fabaceae: A conceptual model and its ecological implications for the australian wattle Acacia saligna. Ann. Bot. 2014, 114, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.; Lazzaro, L.; Calamassi, R.; Fico, G.; Foggi, B.; Mariotti Lippi, M. Induced water stress affects seed germination response and root anatomy in Robinia pseudoacacia (Fabaceae). Trees-Struct. Funct. 2019, 33, 1627–1638. [Google Scholar] [CrossRef]
- Tuyet, D. Characteristics of karst ecosystems of Vietnam and their vulnerability to human impact. Acta Geol. Sin.-Engl. Ed. 2010, 75, 325–329. [Google Scholar] [CrossRef]
- Tang, J.W.; Lü, X.T.; Yin, J.X.; Qi, J.F. Diversity, composition and physical structure of tropical forest over limestone in Xishuangbanna, South-West China. J. Trop. For. Sci. 2011, 23, 425–433. [Google Scholar]
- Fahey, T.J.; Sherman, R.E.; Tanner, E.V.J. Tropical montane cloud forest: Environmental drivers of vegetation structure and ecosystem function. J. Trop. Ecol. 2016, 32, 355–367. [Google Scholar] [CrossRef]
- Nie, Y.P.; Chen, H.S.; Wang, K.L.; Yang, J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of southwest China. J. Hydrol. 2012, 420, 264–274. [Google Scholar] [CrossRef]
- Lin, W.; Wen, C.; Zhou, W.; Huang, G. Drought in southwest China: A review. Atmos. Ocean. Sci. Lett. 2015, 8, 339–344. [Google Scholar] [CrossRef]
- Feng, W.J.; Leung, M.Y.T.; Wang, D.X.; Zhou, W.; Zhang, O.Y.W. An extreme drought over South China in 2020/21 concurrent with an unprecedented warm northwest Pacific and La Niña. Adv. Atmos. Sci. 2022, 39, 1637–1649. [Google Scholar] [CrossRef]
- You, Q.L.; Jiang, Z.H.; Yue, X.; Guo, W.D.; Liu, Y.G.; Cao, J.; Li, W.; Wu, F.Y.; Cai, Z.Y.; Zhu, H.H.; et al. Recent frontiers of climate changes in East Asia at global warming of 1.5 °C and 2 °C. NPJ Clim. Atmos. Sci. 2022, 5, 1–17. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Assessing the vulnerability of oak (Quercus) forest ecosystems under projected climate and land use land cover changes in western Himalaya. Biodivers. Conserv. 2019, 28, 2275–2294. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Meng, Q.X.; Wang, Y.Z.; Zhang, X.L.; Wang, W. Climate change-induced migration patterns and extinction risks of Theaceae species in China. Ecol. Evol. 2020, 10, 4352–4361. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Jiang, X.-L.; Guo, K.Q.; Byrne, A.; Deng, M. Climate change impacts the distribution of Quercus section Cyclobalanopsis (Fagaceae), a keystone lineage in East Asian evergreen broadleaved forests. Plant Divers. 2023, 45, 552–568. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Tu, Y.; Li, Q.; Deng, M. Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications. Forests 2024, 15, 235. https://doi.org/10.3390/f15020235
Liu L, Tu Y, Li Q, Deng M. Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications. Forests. 2024; 15(2):235. https://doi.org/10.3390/f15020235
Chicago/Turabian StyleLiu, Luting, Yu Tu, Qiansheng Li, and Min Deng. 2024. "Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications" Forests 15, no. 2: 235. https://doi.org/10.3390/f15020235
APA StyleLiu, L., Tu, Y., Li, Q., & Deng, M. (2024). Seed Germination Characteristics of a Critically Endangered Evergreen Oak—Quercus marlipoensis (Fagaceae) and Their Conservation Implications. Forests, 15(2), 235. https://doi.org/10.3390/f15020235