Can the Impact of Gravel Roads on Organic Layer Thickness Explain the Distribution of Populus tremuloides along Road Networks in the Boreal Forest of Eastern Canada?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Does the thickness of the organic layer (hereafter OLT) have an impact on the distribution of aspen in the study landscape?
- Do forest roads have an impact on the OLT via the transport of mineral particles from the roads, thus promoting the establishment of aspen?
- Can the impact of roads on OLT explain the distribution of aspen along the road network, and in particular, the role of roads as habitat corridors?
2. Materials and Methods
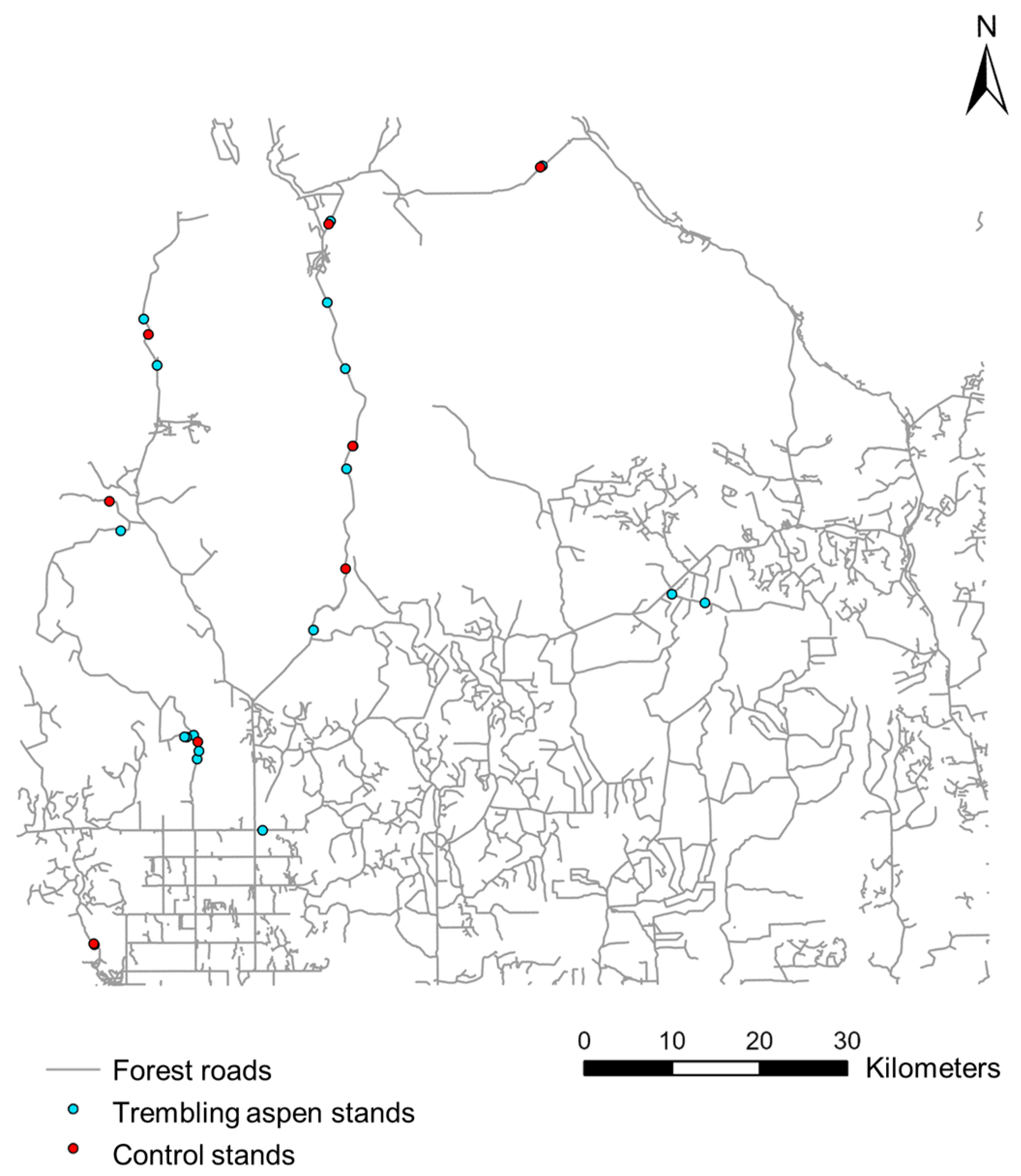
2.1. Study Area
2.2. Organic Layer Thickness and Aspen Distribution
2.2.1. Databases
2.2.2. Data Analyses
2.3. Organic Layer Thickness and Aspen Distribution
2.3.1. Stand Sampling
2.3.2. Data Analyses
2.4. Mineral Soil and Germination of Aspen: Experimental Setup
- Mineral soil only;
- Mosses covered with 2 cm of mineral soil;
- Mosses covered with 0.5 cm of mineral soil;
- Mosses only.
3. Results
3.1. Organic Layer Thickness and Aspen Distribution
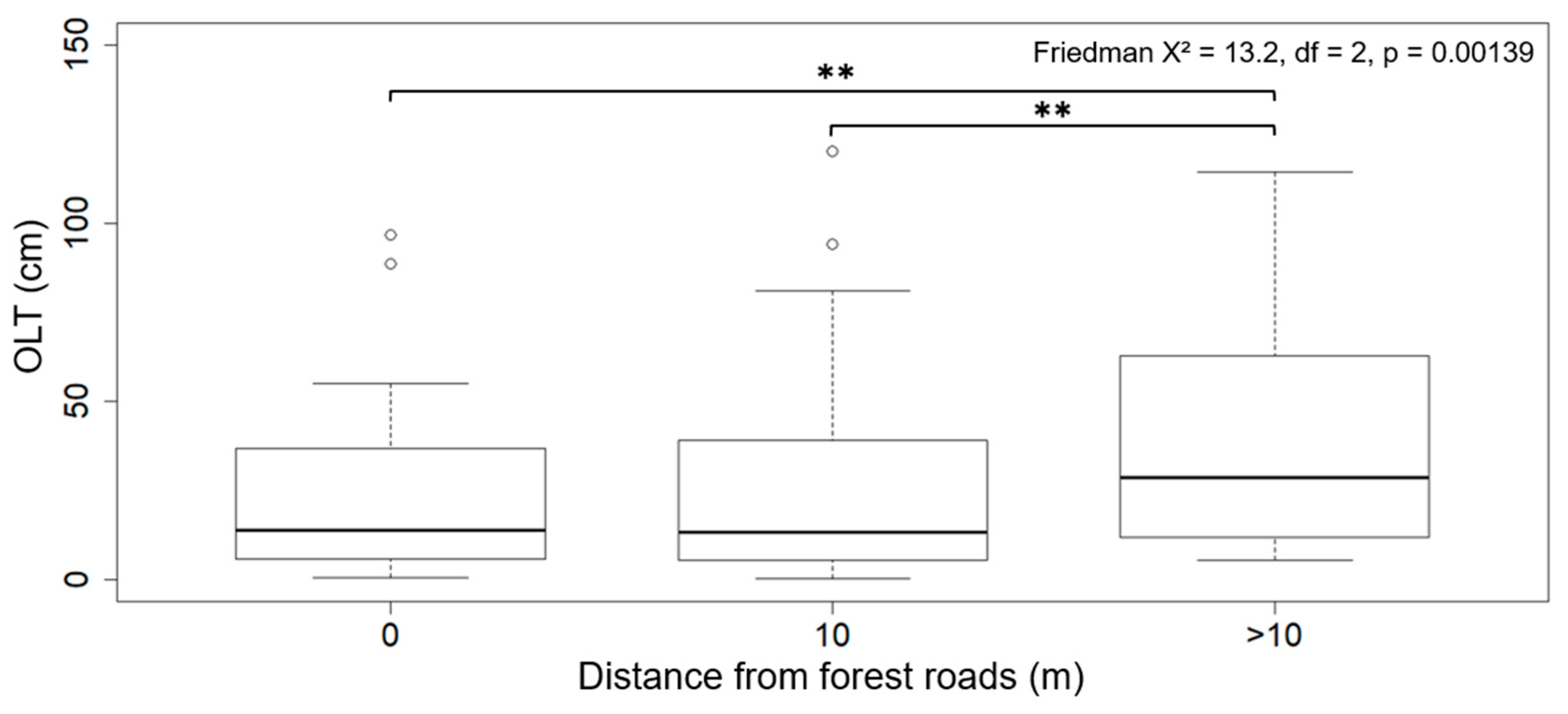
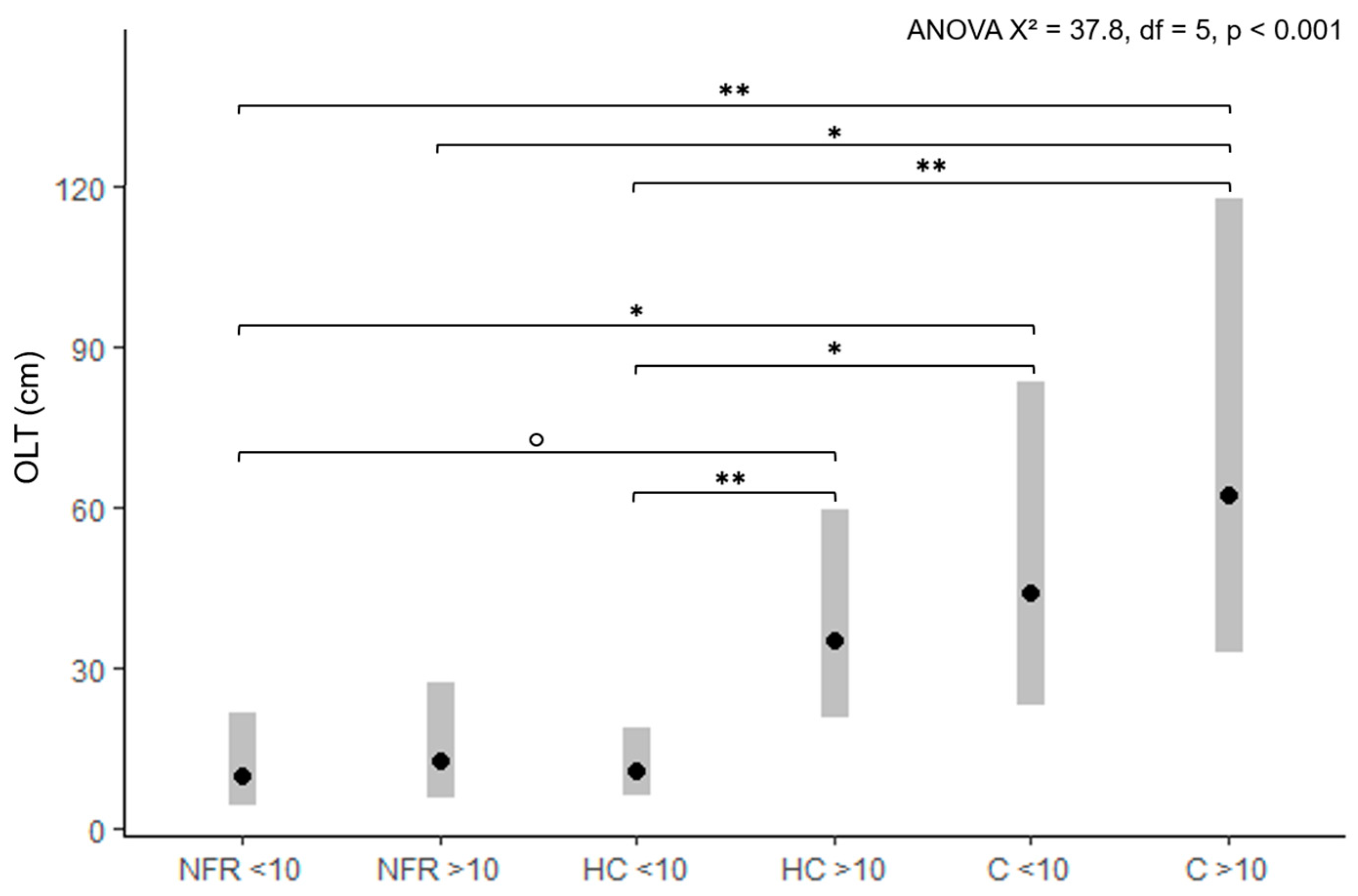
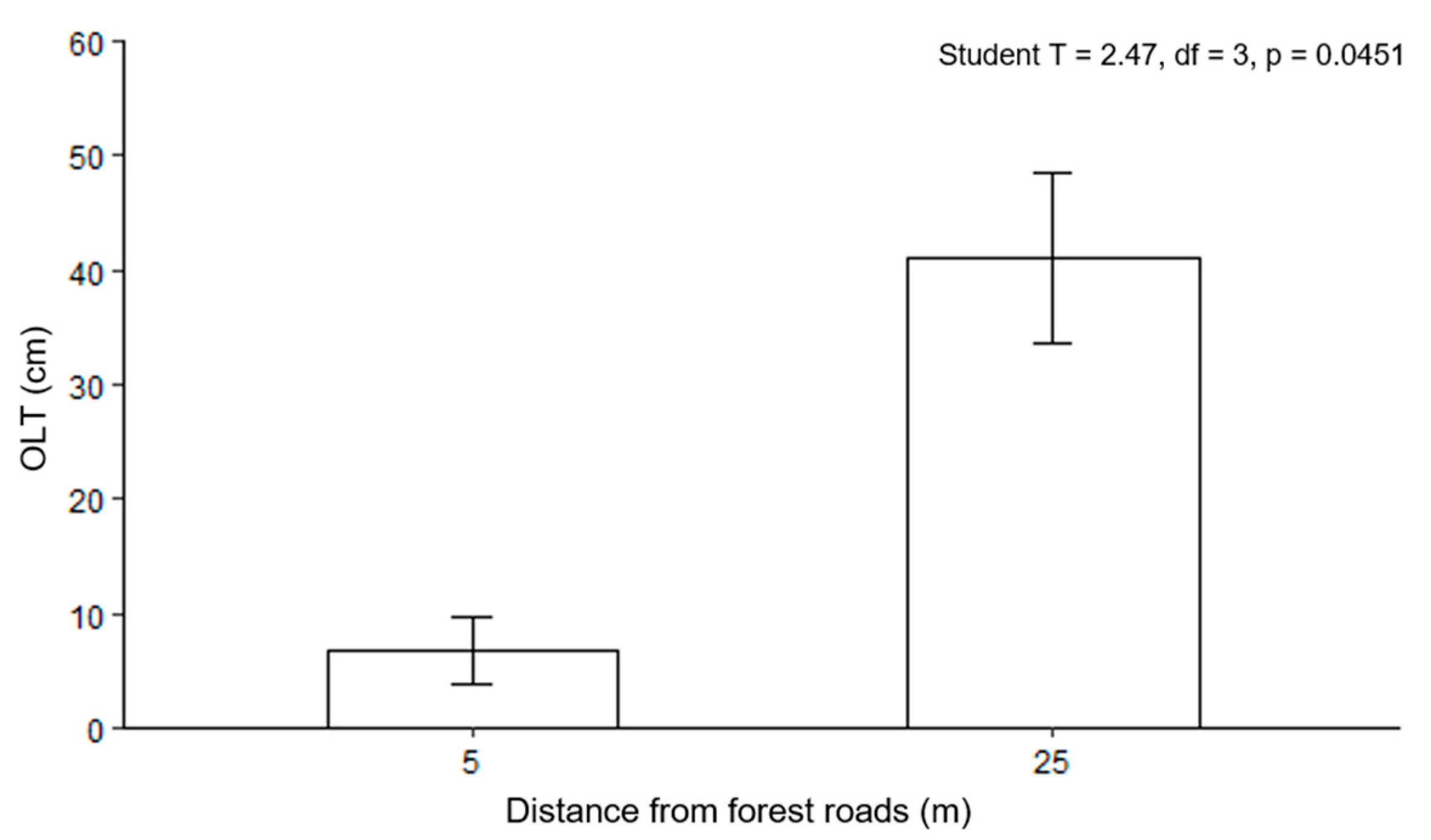
3.2. Forest Roads and Organic Layer Thickness
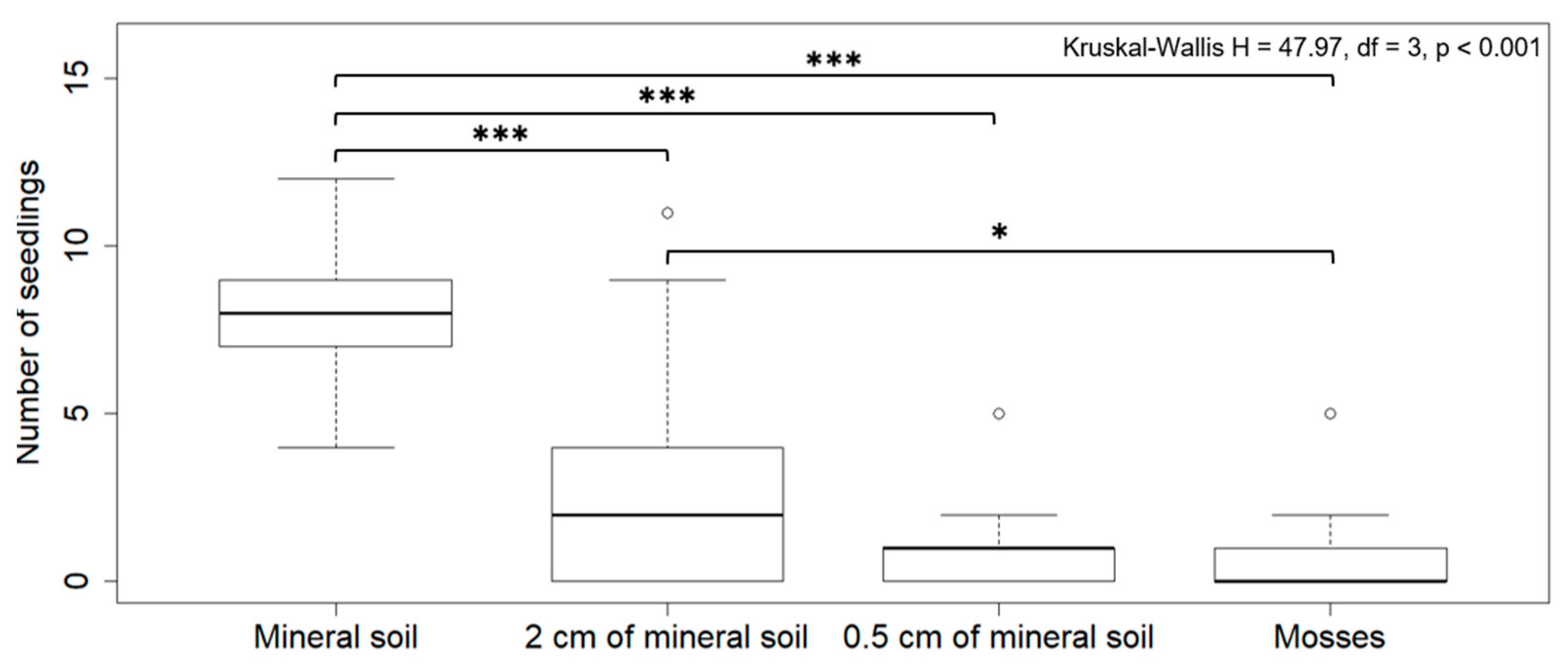
3.3. Mineral Soil and Aspen Germination
4. Discussion
4.1. Organic Layer Thickness and Aspen Distribution
4.2. Forest Roads and Organic Layer Thickness
4.3. Mineral Soil and Aspen Germination
4.4. Aspen Distribution along the Road Network
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forman, R.; Sperling, D.; Bissonette, J.; Clevenger, A.; Cutshall, C.; Dale, V.; Fahrig, L.; France, R.; Goldman, C.; Heanue, K.; et al. Road Ecology: Science and Solutions; Island Press: Washington DC, USA, 2003; p. 481. [Google Scholar]
- Harper, K.; Macdonald, S.; Burton, P.; Chen, J.; Brosofske, K.; Saunders, S.; Euskirchen, E.; Roberts, D.; Jaiteh, M.; Esseen, P.-A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Pohlman, C.; Turton, S.; Goosem, M. Temporal variation in microclimatic edge effects near powerlines, highways and streams in Australian tropical rainforest. Agric. For. Meteorol. 2009, 149, 84–95. [Google Scholar] [CrossRef]
- Schmidt, M.; Jochheim, H.; Kersebaum, K.-C.; Lischeid, G.; Nendel, C. Gradients of microclimate, carbon and nitrogen in transition zones of fragmented landscapes—A review. Agric. For. Meteorol. 2017, 232, 659–671. [Google Scholar] [CrossRef]
- Stern, E.; Riva, F.; Nielsen, S. Effects of narrow linear disturbances on light and wind patterns in fragmented boreal forests in northeastern Alberta. Forests 2018, 9, 486. [Google Scholar] [CrossRef]
- Trombulak, S.; Frissell, C. Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef]
- Bergès, L.; Chevalier, R.; Avon, C. Influence of forest road, road-surfacing material and stand age on floristic diversity and composition in a nutrient-poor environment. Appl. Veg. Sci. 2013, 16, 470–479. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedam, N. The impact of forest paths upon adjacent vegetation: Effects of the path surfacing material on the species composition and soil compaction. Biol. Conserv. 2004, 119, 405–419. [Google Scholar] [CrossRef]
- Greenberg, C.; Crownover, S.; Gordon, D. Roadside soils: A corridor for invasion of xeric scrub by nonindigenous plants. Nat. Areas J. 1997, 17, 99–109. [Google Scholar]
- Auerbach, N.; Walker, M.; Walker, D. Effects of roadside disturbance on substrate and vegetation properties in arctic tundra. Ecol. Appl. 1997, 7, 218–235. [Google Scholar] [CrossRef]
- Avon, C.; Dumas, Y.; Bergès, L. Management practices increase the impact of roads on plant communities in forests. Biol. Conserv. 2013, 159, 24–31. [Google Scholar] [CrossRef]
- Müellerová, J.; Vítková, M.; Vítek, O. The impacts of road and walking trails upon adjacent vegetation: Effects of road building materials on species composition in a nutrient poor environment. Sci. Total Environ. 2011, 409, 3839–3849. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.; Alexander, L. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Santelmann, M.V.; Gorham, E. The influence of airborne road dust on the chemistry of Sphagnum mosses. J. Ecol. 1988, 76, 1219–1231. [Google Scholar] [CrossRef]
- Andrews, A. Fragmentation of habitat by roads and utility corridors: A review. Aust. Zool. 1990, 26, 130–141. [Google Scholar] [CrossRef]
- Laurance, W.; Goosem, M.; Laurance, S. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Seiler, A. Ecological Effects of Roads: A Review; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2001; p. 40. [Google Scholar]
- Spellerberg, I. Ecological effects of roads and traffic: A literature review. Glob. Ecol. Biogeogr. Lett. 1998, 7, 317–333. [Google Scholar] [CrossRef]
- Najafi, A.; Torabi, M.; Nowbakht, A.-A.; Moafi, M.; Eslami, A.; Sotoudeh Foumani, B. Effect of forest roads on adjacent tree regeneration in a mountainous forest. Ann. Biol. Res. 2012, 3, 1700–1703. [Google Scholar]
- Picchio, R.; Tavankar, F.; Venanzi, R.; Lo Monaco, A.; Nikooy, M. Study of forest road effect on tree community and stand structure in three italian and iranian temperate forests. Croat. J. For. Eng. 2018, 39, 57–70. [Google Scholar]
- Vepakomma, U.; Kneeshaw, D.; De Grandpré, L. Influence of natural and anthropogenic linear canopy openings on forest structural patterns investigated using lidar. Forests 2018, 9, 540. [Google Scholar] [CrossRef]
- Franklin, C.; Harper, K.; Clarke, M. Trends in studies of edge influence on vegetation at human-created and natural forest edges across time and space. Can. J. For. Res. 2021, 51, 274–282. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Mayerhofer, M.S.; Biswas, S.R.; Esseen, P.A.; Hylander, K.; Stewart, K.J.; Mallik, A.U.; Drapeau, P.; Jonsson, B.G.; et al. Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. J. Ecol. 2015, 103, 550–562. [Google Scholar] [CrossRef]
- Perala, D. Populus tremuloides Michx. quaking aspen. In Silvics of North America: Hardwoods; Burns, R., Honkala, B., Eds.; US Department of Agriculture, Forest Service: Washington DC, USA, 1990; Volume 2, pp. 555–569. [Google Scholar]
- Brumelis, G.; Carleton, T. The vegetation of postlogged black spruce lowlands in central Canada. I. Trees and tall shrubs. Can. J. For. Res. 1988, 18, 1470–1478. [Google Scholar] [CrossRef]
- Carleton, T.; MacLellan, P. Woody vegetation responses to fire versus clear-cutting logging: A comparative survey in the central Canadian boreal forest. Ecoscience 1994, 1, 141–152. [Google Scholar] [CrossRef]
- Chen, H.; Vasiliauskas, S.; Kayahara, G.; Ilisson, T. Wildfire promotes broadleaves and species mixture in boreal forest. For. Ecol. Manag. 2009, 257, 343–350. [Google Scholar] [CrossRef]
- Weir, J.; Johnson, E. Effects of escaped settlement fires and logging on forest composition in the mixedwood boreal forest. Canadian J. For. Res. 1998, 28, 459–467. [Google Scholar] [CrossRef]
- Bartos, D.; Mueggler, W. Early succession in aspen communities following fire in western Wyoming. J. Range Manag. 1981, 34, 315–318. [Google Scholar] [CrossRef]
- Bartos, D.; Mueggler, W. Early succession following clearcutting of aspen communities in Northern Utah. J. Range Manag. 1982, 35, 764–768. [Google Scholar] [CrossRef]
- Kay, C. Aspen seedlings in recently burned areas of Grand Teton and Yellowstone National Parks. Northwest Sci. 1993, 67, 94–104. [Google Scholar]
- Quinn, R.; Wu, L. Quaking aspen reproduce from seed after wildfire in the mountains of southeastern Arizona. In Proceedings of the Sustaining Aspen in Western Landscapes: Symposium Proceedings, Grand Junction, CO, USA, 13–15 June 2000; pp. 369–376. [Google Scholar]
- Jobidon, R. Autécologie de Quelques Espèces de Compétition D’importance pour la Régénération Forestière au Québec. Revue de Littérature; Mémoire de Recherche Forestière No. 117; Gouvernement du Québec, Ministère des Ressources Naturelles, Direction de la Recherche Forestière: Québec, QC, Canada, 1995; p. 180.
- Turner, M.; Romme, W.; Reed, R.; Tuskan, G. Post-fire aspen seedling recruitment across the Yellowstone (USA) landscape. Landsc. Ecol. 2003, 18, 127–140. [Google Scholar] [CrossRef]
- Whitbeck, K.; Oetter, D.; Perry, D.; Fyles, J. Interactions between macroclimate, microclimate, and anthropogenic disturbance affect the distribution of aspen near its northern edge in Quebec: Implications for climate change related range expansions. For. Ecol. Manag. 2016, 368, 194–206. [Google Scholar] [CrossRef]
- Marchais, M.; Arseneault, D.; Bergeron, Y. Forest roads act as habitat corridors for Populus tremuloides in the boreal forest of eastern Canada. Front. Ecol. Evol. 2024, 11, 1336409. [Google Scholar] [CrossRef]
- Gewehr, S.; Drobyshev, I.; Berninger, F.; Bergeron, Y. Soil characteristics mediate the distribution and response of boreal trees to climatic variability. Can. J. For. Res. 2014, 44, 487–498. [Google Scholar] [CrossRef]
- Lafleur, B.; Cazal, A.; Leduc, A.; Bergeron, Y. Soil organic layer thickness influences the establishment and growth of trembling aspen (Populus tremuloides) in boreal forests. For. Ecol. Manag. 2015, 347, 209–216. [Google Scholar] [CrossRef]
- Lavoie, M.; Paré, D.; Fenton, N.; Groot, A.; Taylor, K. Paludification and management of forested peatlands in Canada: A literature review. Environ. Rev. 2005, 13, 21–50. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef]
- Taylor, S.; Carleton, T.; Adams, R. Understorey vegetation change in a Picea mariana chronosequence. Vegetatio 1987, 73, 63–72. [Google Scholar] [CrossRef]
- van Cleve, K.; Dyrness, C.; Viereck, L.; Fox, J.; Chapin, F.; Oechel, W. Taiga ecosystems in interior Alaska. Bioscience 1983, 33, 39–44. [Google Scholar] [CrossRef]
- Clymo, R. The limits to peat bog growth. Philos. Trans. R. Soc. Lond. B 1984, 303, 605–654. [Google Scholar] [CrossRef]
- Fenton, N.; Légaré, S.; Bergeron, Y.; Paré, D. Soil oxygen within boreal forests across an age gradient. Can. J. Soil Sci. 2006, 86, 1–9. [Google Scholar] [CrossRef]
- Fenton, N.; Lecomte, N.; Légaré, S.; Bergeron, Y. Paludification in black spruce (Picea mariana) forests of eastern Canada: Potential factors and management implications. For. Ecol. Manag. 2005, 213, 151–159. [Google Scholar] [CrossRef]
- Fenton, N.; Bergeron, Y. Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. J. Veg. Sci. 2006, 17, 65–76. [Google Scholar] [CrossRef]
- Krasny, M.; Zasada, J.; Vogt, K. Adventitious rooting of four Salicaceae species in response to a flooding event. Can. J. Bot. 1988, 66, 2597–2598. [Google Scholar] [CrossRef]
- Landhäusser, S.; Lieffers, V.; Mulak, T. Effects of soil temperature and time of decapitation on sucker initiation of intact Populus tremuloides root systems. Scand. J. For. Res. 2006, 21, 299–305. [Google Scholar] [CrossRef]
- Belleau, A.; Leduc, A.; Lecomte, N.; Bergeron, Y. Forest succession rate and pathways on different surface deposit types in the boreal forest of northwestern Quebec. Ecoscience 2011, 18, 320–340. [Google Scholar] [CrossRef]
- Bourgeois, L.; Kneeshaw, D.; Boisseau, G. Les routes forestières au Québec: Les impacts environnementaux, sociaux et économiques. VertigO 2005, 6, 1–9. [Google Scholar] [CrossRef]
- MFFP. Ressources et Industries Forestières du Québec—Portrait Statistique 2018; Gouvernement du Québec, Ministère des Forêts, de la Faune et des Parcs, Direction de la Modernisation de L’industrie des Produits Forestiers: Québec, QC, Canada, 2019.
- MRNF. Ressources et Industries Forestières du Québec—Portrait Statistique 2021; Gouvernement du Québec, Ministère des Ressources Naturelles et des Forêts, Direction du Dévloppement et de L’innovation de L’industrie: Québec, QC, Canada, 2022; p. 138.
- Farmer, A. The effects of dust on vegetation—A review. Environ. Pollut. 1993, 79, 63–75. [Google Scholar] [CrossRef]
- Fenton, N.; Frego, K.; Sims, M. Changes in forest floor bryophyte (moss and liverwort) communities 4 years after forest harvest. Can. J. Bot. 2003, 81, 714–731. [Google Scholar] [CrossRef]
- Moen, J.; Jonsson, B. Edge effects on liverworts and lichens in forest patches in a mosaic of boreal forest and wetland. Conserv. Biol. 2003, 17, 380–388. [Google Scholar] [CrossRef]
- Ackerman, D.; Breen, A. Infrastructure development accelerates range expansion of trembling aspen (Populus tremuloides, Salicaceae) into the arctic. Arctic 2016, 69, 130–136. [Google Scholar] [CrossRef]
- McKendrick, J. Soils and Vegetation of the Trans-Alaska Pipeline Route: A 1999 Survey. Bulletin 109; School of Agriculture and Land Resources Management, Agricultural and Forestry Experiment Station: Fairbanks, AK, USA, 2002. [Google Scholar]
- Veillette, J. Evolution and paleohydrology of glacial lakes Barlow and Ojibway. Quat. Sci. Rev. 1994, 13, 945–971. [Google Scholar] [CrossRef]
- Blouin, J.; Berger, J.-P. Guide de Reconnaissance des Types Écologiques de la Région Écologique 6a-PLAINE du lac Matagami et 6b-Plaine de la baie de Rupert; Gouvernement du Québec, Ministère des Ressources naturelles et de la Faune, Direction des inventaires forestiers, Division de la classification écologique et productivité des stations: Québec, QC, Canada, 2005; p. 97.
- EC. Normales Climatiques Canadiennes: Normales et Moyennes Climatiques de 1981–2010. Available online: https://climat.meteo.gc.ca/climate_normals/ (accessed on 1 October 2023).
- Baldwin, K.; MacKenzie, W.; Pfalz, J.; Meades, W.; Meidinger, D.; Robitaille, A.; Saucier, J.-P.; Uhlig, P. Level 4 Map, Version 1, Canadian Component of the Circumboreal Vegetation Map (CBVM); Canadian Regional Team of the Circumboreal Vegetation Map Project; Natural Resources Canada, Canadian Forest Service, Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 2012. [Google Scholar]
- Boulanger, Y.; Gauthier, S.; Gray, D.; Le Goff, H.; Lefort, P.; Morissette, J. Fire regime zonation under current and future climate over eastern Canada. Ecol. Appl. 2013, 23, 904–923. [Google Scholar] [CrossRef] [PubMed]
- Erni, S.; Wang, X.; Taylor, S.; Boulanger, Y.; Swystun, T.; Flannigan, M.; Parisien, M.-A. Developing a two-level fire regime zonation system for Canada. Can. J. For. Res. 2020, 50, 259–273. [Google Scholar] [CrossRef]
- FEC. Possibilités forestières 2023-2028. Rapport du Calcul De L’unité D’aménagement 085-51, Région du Nord-du-Québec; Gouvernement du Québec, Bureau du Forestier en Chef: Roberval, QC, Canada, 2021; p. 46.
- SIFORT. Système d’information Forestière par Tesselle (SIFORT). Available online: https://www.donneesquebec.ca/recherche/dataset/systeme-d-information-forestiere-par-tesselle-sifort (accessed on 1 October 2023).
- MFFP. Normes de Stratification Écoforestière—Quatrième Inventaire Écoforestier du Québec Méridionnal; Gouvernement du Québec, Ministère des Forêts, de la Faune et des Parcs, Direction des Inventaires Forestiers: Québec, QC, Canada, 2015; p. 78.
- Mansuy, N.; Valeria, O.; Laamrani, A.; Fenton, N.; Guindon, L.; Bergeron, Y.; Beaudoin, A.; Légaré, S. Digital mapping of paludification in soils under black spruce forests of eastern Canada. Geoderma Reg. 2018, 15, e00194. [Google Scholar] [CrossRef]
- R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, R Core Team: Vienna, Austria, 2020. [Google Scholar]
- MRNF. AQréseau+. Available online: https://www.donneesquebec.ca/recherche/dataset/adresses-quebec/resource/0d5df103-f856-4183-9a15-eb06fce9c8bf#:~:text=Ce%20produit%20propose%20une%20nouvelle,fichier%20plat%20(Flat%20File) (accessed on 1 October 2023).
- GTCS. Le Système Canadien de Classification des Sols, 3rd ed.; Direction Générale de la Recherche, Ministère de l’Agriculture et de l’Agro-Alimentaire du Canada, Groupe de Travail Sur la Classification des Sols: Ottawa, ON, Canada, 2002. [Google Scholar]
- Eisinga, R.; Heskes, T.; Pelzer, B.; Te Grotenhuis, M. Exact p-values for pairwise comparison of Friedman rank sums, with application to comparing classifiers. BMC Bioinform. 2017, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR), R Package; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Kassambara, A. Package ‘rstatix’. Pipe-Friendly Framework for Basic Statistical Tests, R Package; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R.; Bolker, B.; Buerkner, P.; Giné-Vázquez, I.; Herve, M.; Jung, M.; Love, J.; Miguez, F.; Riebl, H.; Singmann, H. emmeans: Estimated Marginal Means, aka Least-Squares Means, R Package; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Landhäusser, S.; Pinno, B.; Mock, K. Tamm Review: Seedling-based ecology, management, and restoration in aspen (Populus tremuloides). For. Ecol. Manag. 2019, 432, 231–245. [Google Scholar] [CrossRef]
- Alberti, G. GmAMisc: Gianmarco Alberti Miscellaneous, R Package; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Prescott, C.; Maynard, D.; Laiho, R. Humus in northern forests: Friend or foe? For. Ecol. Manag. 2000, 133, 23–36. [Google Scholar] [CrossRef]
- Landhäusser, S.; DesRochers, A.; Lieffers, V. A comparison of growth and physiology in Picea glauca and Populus tremuloides at different soil temperatures. Can. J. For. Res. 2001, 31, 1922–1929. [Google Scholar] [CrossRef]
- Wan, X.; Landhäusser, S.; Zwiazek, J.; Lieffers, V. Root water flow and growth of aspen (Populus tremuloides) at low root temperatures. Tree Physiol. 1999, 19, 879–884. [Google Scholar] [CrossRef]
- Landhäusser, S.; Silins, U.; Lieffers, V.; Liu, W. Response of Populus tremuloides, Populus balsamifera, Betula papyrifera and Picea glauca seedlings to low soil temperature and water-logged soil conditions. Scand. J. For. Res. 2003, 18, 391–400. [Google Scholar] [CrossRef]
- Schier, G.; Campbell, R. Differences among Populus species in ability to form adventitious shoots and roots. Can. J. For. Res. 1976, 6, 253–261. [Google Scholar] [CrossRef]
- Myers-Smith, I.; Arnesen, B.; Thompson, R.; Chapin, F. Cumulative impacts on Alaskan arctic tundra of a quarter century of road dust. Ecoscience 2006, 13, 503–510. [Google Scholar] [CrossRef]
- Johnston, F.; Johnston, S. Impacts of road disturbance on soil properties and on exotic plant occurrence in subalpine areas of the Australian Alps. Arct. Antarct. Alp. Res. 2004, 36, 201–207. [Google Scholar] [CrossRef]
- Kalisz, P.; Powell, J. Effect of calcareous road dust on land snails (Gastropoda: Pulmonata) and millipedes (Diplopoda) in acid forest soils of the Daniel Boone National Forest of Kentucky, USA. For. Ecol. Manag. 2003, 186, 177–183. [Google Scholar] [CrossRef]
- Clymo, R. The growth of Sphagnum: Some effects of environment. J. Ecol. 1973, 61, 849–869. [Google Scholar] [CrossRef]
- Spatt, P.; Miller, M. Growth conditions and vitality of Sphagnum in a tundra community along the Alaska pipeline haul road. Arctic 1981, 34, 48–54. [Google Scholar] [CrossRef]
- Walker, D.A.; Everett, K.R. Road dust and its environmental impact on Alaskan taiga and tundra. Arct. Alp. Res. 1987, 19, 479–489. [Google Scholar] [CrossRef]
- DesRochers, A.; van den Driessche, R.; Thomas, B. Nitrogen fertilization of trembling aspen seedlings grown on soils of different pH. Can. J. For. Res. 2003, 33, 552–560. [Google Scholar] [CrossRef]
- Gifford, G. The Influence of Growth Media, Temperatures, and Light Intensities on Aspen Root and Top Growth; Research Note INT-67; US Department of Agriculture, Forest Service, Intermountain Forest & Range Experiment Station: Ogden, UT, USA, 1967; p. 4.
- Stoeckeler, J. Soil Factors Affecting the Growth of Quaking Aspen Forests in the Lake States; Technical Bulletin; University of Minnesota: St. Paul, MN, USA, 1960; p. 48. [Google Scholar]
- Gill, H.; Lantz, T.; O’Neill, B.; Kokelj, S. Cumulative impacts and feedbacks of a gravel road on shrub tundra ecosystems in the Peel Plateau, Northwest Territories, Canada. Arct. Antarct. Alp. Res. 2014, 46, 947–961. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R. Linking the abundance of aspen with soil faunal communities and rates of belowground processes within single stands of mixed aspen–black spruce. Appl. Soil Ecol. 2009, 41, 19–28. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R. How does a tree species influence litter decomposition? Separating the relative contribution of litter quality, litter mixing, and forest floor conditions. Can. J. For. Res. 2010, 40, 465–475. [Google Scholar] [CrossRef]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of aspen on forest floor properties in black spruce-dominated stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Meininger, C.; Spatt, P. Variations of tardigrade assemblages in dust-impacted Arctic mosses. Arct. Alp. Res. 1988, 20, 24–30. [Google Scholar] [CrossRef]
- Ackerman, D.; Finlay, J. Road dust biases NDVI and alters edaphic properties in Alaskan arctic tundra. Sci. Rep. 2019, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Creuzer, J.; Hargiss, C.; Norland, J.; DeSutter, T.; Casey, F.; DeKeyser, E.; Ell, M. Does Increased Road Dust Due to Energy Development Impact Wetlands in the Bakken Region? Water Air Soil Pollut. 2016, 227, 39–52. [Google Scholar] [CrossRef]
- Tamm, C.; Troedsson, T. An example of the amounts of plant nutrients supplied to the ground in road dust. Oikos 1955, 6, 61–70. [Google Scholar] [CrossRef]
- Landhäusser, S.; Deshaies, D.; Lieffers, V. Disturbance facilitates rapid range expansion of aspen into higher elevations of the Rocky Mountains under a warming climate. J. Biogeogr. 2010, 37, 68–76. [Google Scholar] [CrossRef]
- Johnstone, J.; Chapin, F. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 2006, 9, 14–31. [Google Scholar] [CrossRef]
- Faust, M. Germination of Populus grandidentata and P. tremuloides, with particular reference to oxygen consumption. Bot. Gaz. 1936, 97, 808–821. [Google Scholar] [CrossRef]
- Moss, E. Longevity of seed and establishment of seedlings in species of Populus. Bot. Gaz. 1938, 99, 529–542. [Google Scholar] [CrossRef]
- Duchesne, S.; Sirois, L. Phase initiale de régénération après feu des populations conifériennes subarctiques. Can. J. For. Res. 1995, 25, 307–318. [Google Scholar] [CrossRef]
- Greene, D.; Zasada, J.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Wolken, J.; Landhäusser, S.; Lieffers, V.; Dyck, M. Differences in initial root development and soil conditions affect establishment of trembling aspen and balsam poplar seedlings. Botany 2010, 88, 275–285. [Google Scholar] [CrossRef]
- McDonough, W. Quaking Aspen—Seed Germination and Early Seedling Growth; Research Paper INT-234; US Department of Agriculture, Forest Service, Intermountain Forest & Range Experiment Station: Ogden, UT, USA, 1979; p. 13.
- Marchais, M.; Arseneault, D.; Bergeron, Y. The rapid expansion of Populus tremuloides due to anthropogenic disturbances in eastern Canada. Can. J. For. Res. 2022, 52, 991–1001. [Google Scholar] [CrossRef]
- Day, M. The root system of aspen. Am. Midl. Nat. 1944, 32, 502–509. [Google Scholar] [CrossRef]
- Ilintsev, A.; Nakvasina, E.; Bogdanov, A. Effects of site preparation methods on soil physical properties and outplanting success of coniferous seedlings in boreal forests. J. For. Res. 2024, 35, 15. [Google Scholar] [CrossRef]
- Prévost, M. Effets du scarifiage sur les propriétés du sol, la croissance des semis et la compétition: Revue des connaissances actuelles et perspectives de recherches au Québec. Ann. For. Sci. 1992, 49, 277–296. [Google Scholar] [CrossRef]
- Sutton, R. Mounding site preparation: A review of European and North American experience. New For. 1993, 7, 151–192. [Google Scholar] [CrossRef]
- Thiffault, N.; Elferjani, R.; Hébert, F.; Paré, D.; Gagné, P. Intensive mechanical site preparation to establish short rotation hybrid poplar plantations—A case-study in Québec, Canada. Forests 2020, 11, 785. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchais, M.; Arseneault, D.; Bergeron, Y. Can the Impact of Gravel Roads on Organic Layer Thickness Explain the Distribution of Populus tremuloides along Road Networks in the Boreal Forest of Eastern Canada? Forests 2024, 15, 298. https://doi.org/10.3390/f15020298
Marchais M, Arseneault D, Bergeron Y. Can the Impact of Gravel Roads on Organic Layer Thickness Explain the Distribution of Populus tremuloides along Road Networks in the Boreal Forest of Eastern Canada? Forests. 2024; 15(2):298. https://doi.org/10.3390/f15020298
Chicago/Turabian StyleMarchais, Mathilde, Dominique Arseneault, and Yves Bergeron. 2024. "Can the Impact of Gravel Roads on Organic Layer Thickness Explain the Distribution of Populus tremuloides along Road Networks in the Boreal Forest of Eastern Canada?" Forests 15, no. 2: 298. https://doi.org/10.3390/f15020298