
Spatial and Temporal Variation in Primary Forest Growth in the Northern Daxing’an Mountains Based on Tree-Ring and NDVI Data

Abstract
:
1. Introduction
2. Materials and Methods
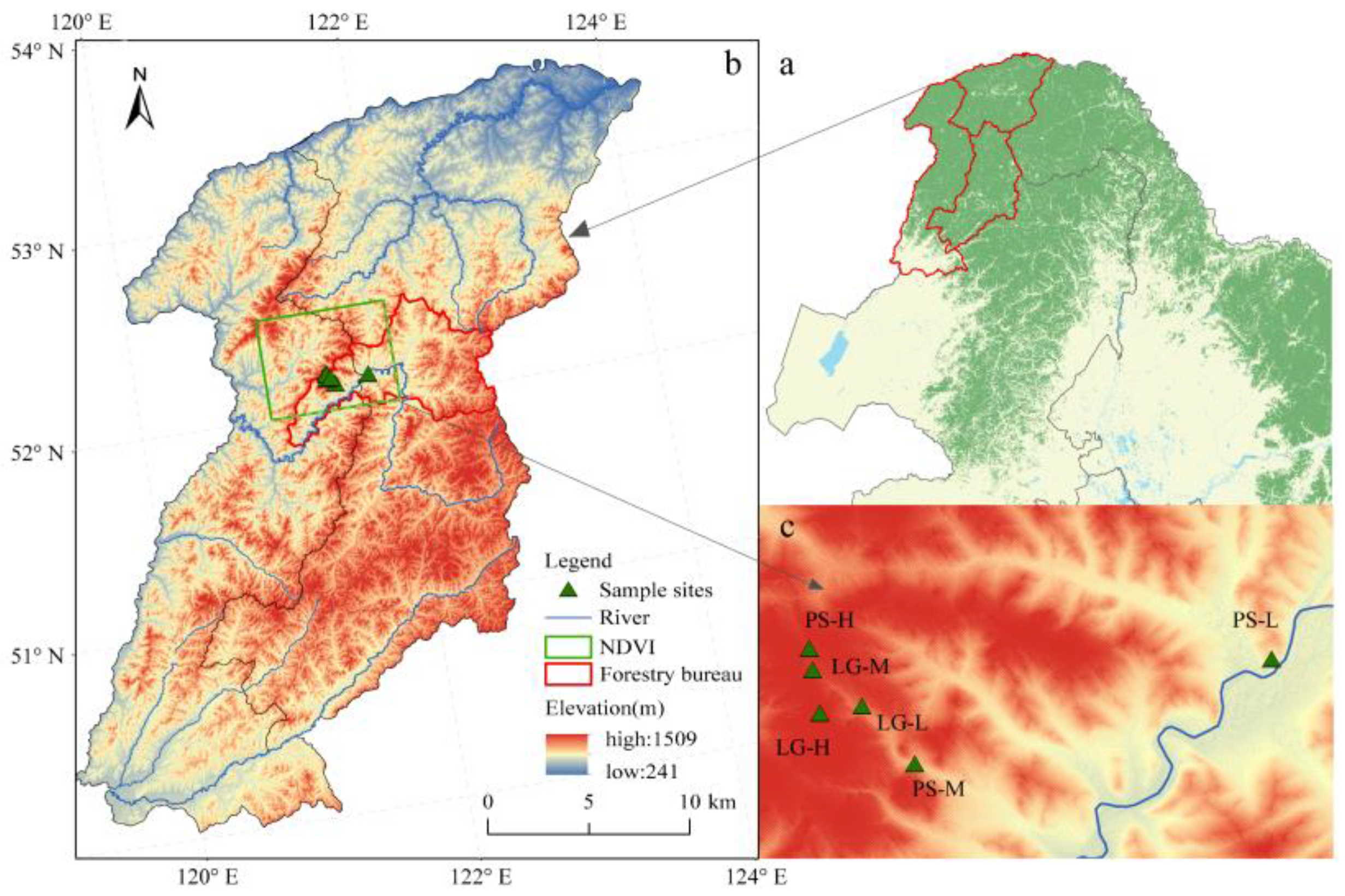
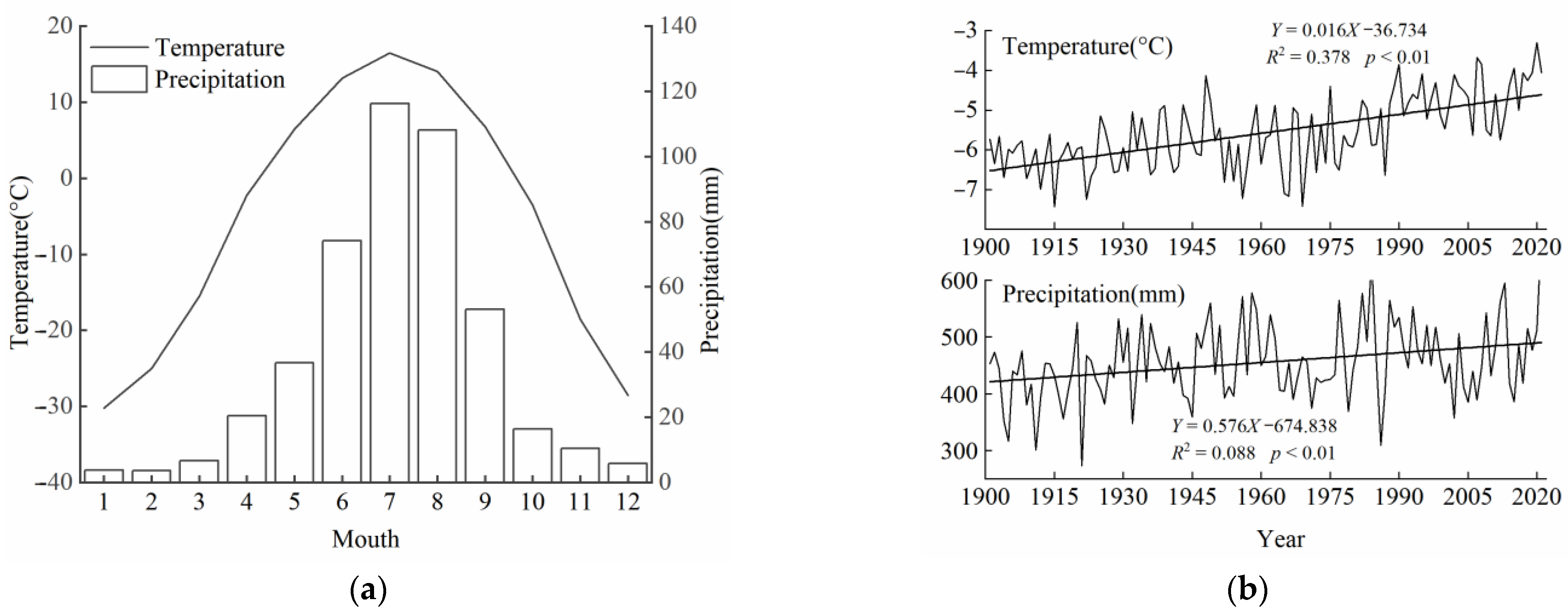
2.1. Study Area
2.2. Dendrochronological Methods
2.3. Meteorological Data and Remote Sensing Images
2.4. Data Analysis
2.4.1. Statistical Analyses
2.4.2. Indicators of Decline in Single-Tree Growth
2.4.3. BFAST Trend Breakpoints
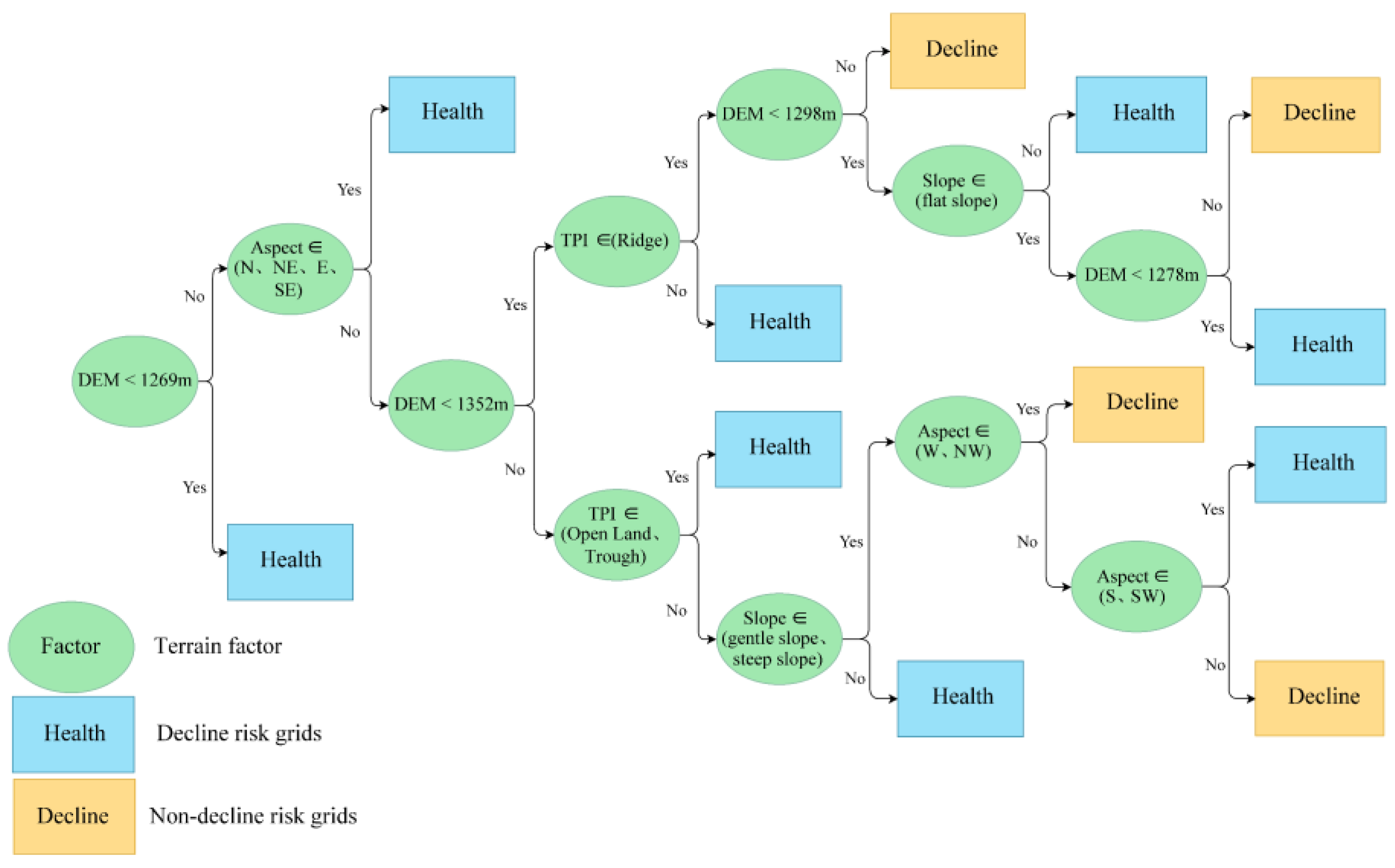
2.4.4. C5.0 Decision Trees
3. Results
3.1. Standardized Chronological Characteristics of Tree Rings
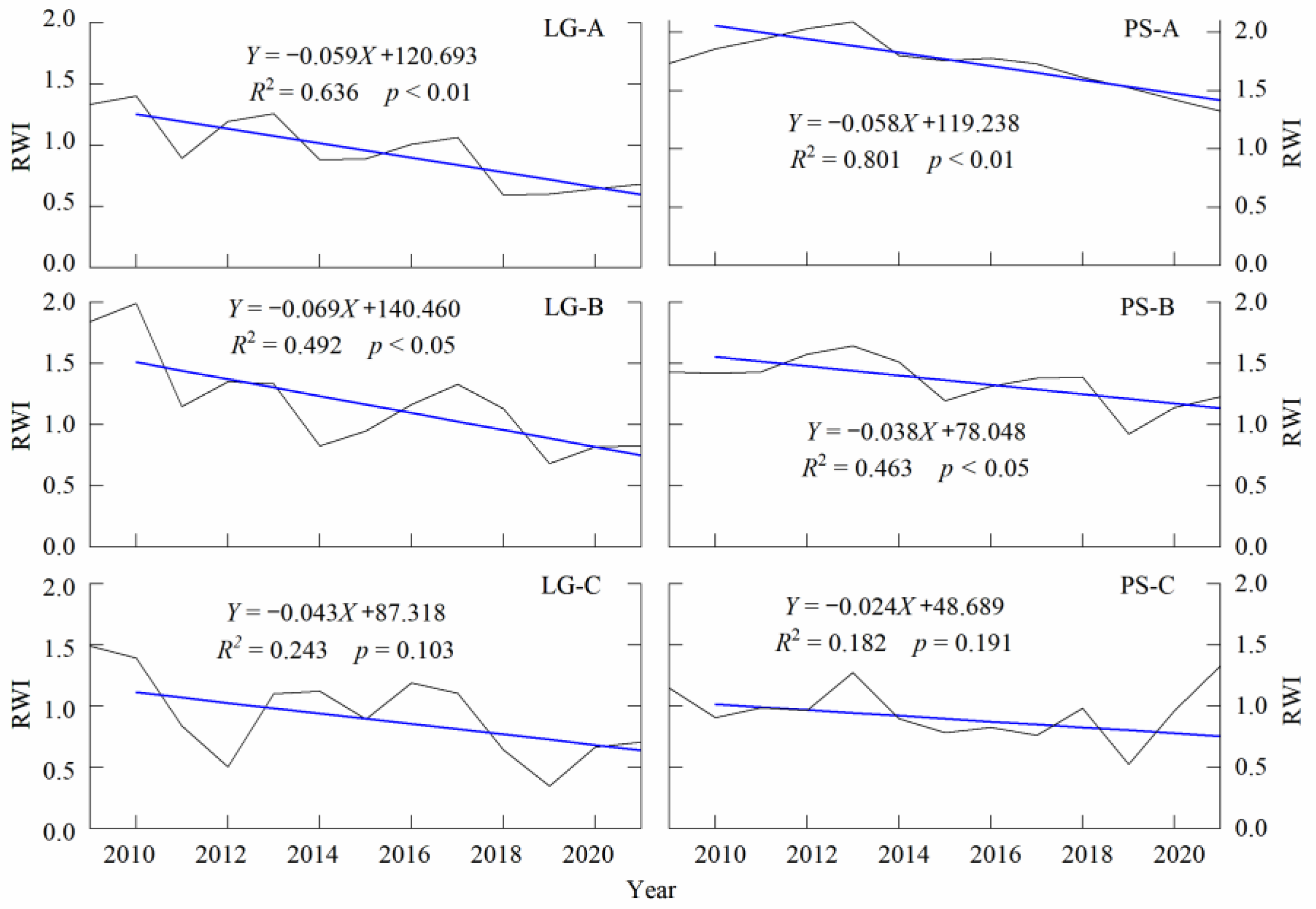
3.2. Decline in Tree Growth
3.3. Principal Component Analysis and Correlation between NDVI and RWI
3.4. Growth Decline Risks
4. Discussion
4.1. Decline in Tree Growth
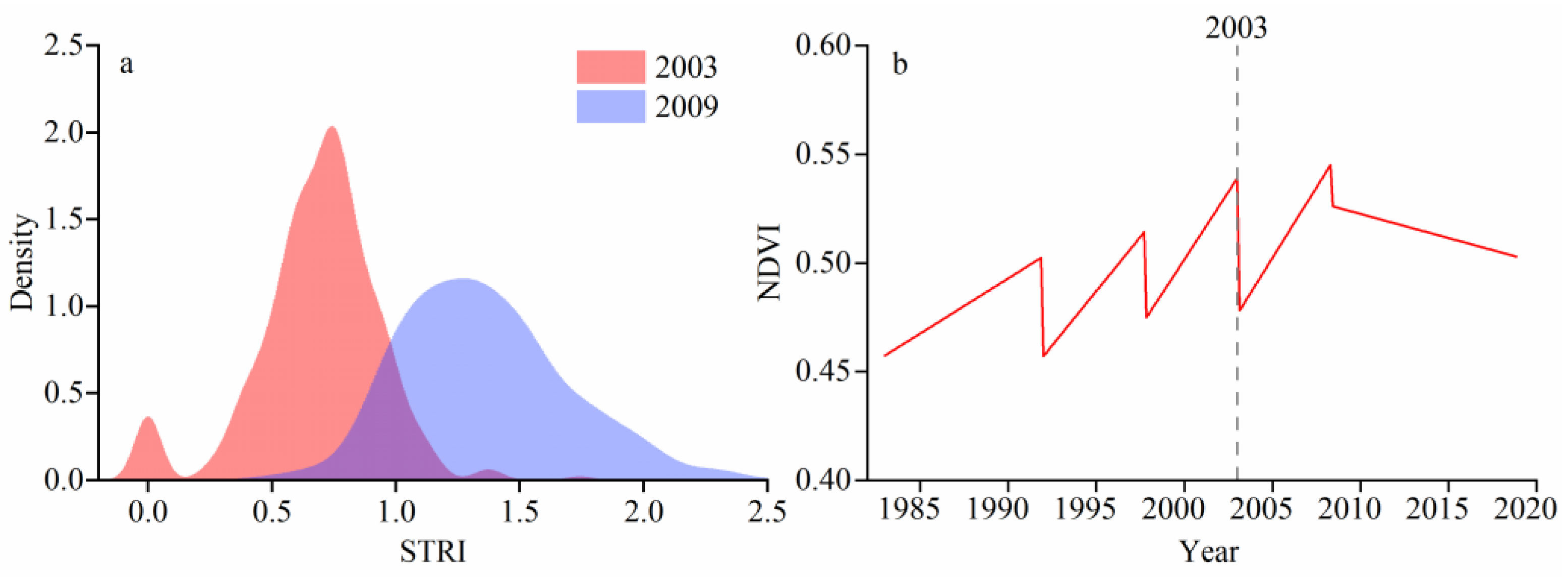
4.2. Trends in Vegetation Canopy Dynamics
4.3. Relationship between NDVI and RWI
4.4. Effects of Topographic Factors on Tree Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Y.H.; Shi, Y.; Sun, W.J.; Chang, J.F. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality. Sci. Sin. 2022, 52, 534–574. [Google Scholar]
- Speizer, S.; Raymond, C.; Ivanovich, C.C.; Horton, R.M. Concentrated and Intensifying Humid Heat Extremes in the IPCC AR6 Regions. Geophys. Res. Lett. 2022, 49, e2021GL097261. [Google Scholar] [CrossRef]
- Orgogozo, L.; Prokushkin, A.; Pokrovsky, O.; Grenier, C.; Quintard, M.; Viers, J.; Audry, S. Water and energy transfer modeling in a permafrost-dominated, forested catchment of Central Siberia: The key role of rooting depth. Permafr. Periglac. Process. 2019, 30, 75–89. [Google Scholar] [CrossRef]
- Liu, H.B.; Mu, M.; Mu, C.C.; Wu, X.D. Interpretation of IPCC Sixth Assessment Report: Observations and Predictions of Permafrost Carbon in the Northern Hemisphere. J. Glaciol. Geocryol. 2023, 45, 318–326. [Google Scholar]
- Prăvălie, R. Major perturbations in the Earth’s forest ecosystems. Possible implications for global warming. Earth Sci. Rev. 2018, 185, 544–571. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Rafał, P. Variability in radial increment can predict an abrupt decrease in tree growth during forest decline: Tree-ring patterns of Abies alba Mill. in near-natural forests. For. Ecol. Manag. 2021, 479, 118579. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.B. History of Tree Growth Declines Recorded in Old Trees at Two Sacred Sites in Northern China. Front. Plant Sci. 2017, 8, 1779. [Google Scholar] [CrossRef]
- Liu, H.Y.; Park Williams, A.; Allen, C.D.; Guo, D.L.; Wu, X.C.; Anenkhonov, O.A.; Liang, E.; Sandanov, D.V.; Yin, Y.; Qi, Z.H.; et al. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob. Chang. Biol. 2013, 19, 2500–2510. [Google Scholar] [CrossRef]
- Takahashi, M.; Feng, Z.Z.; Mikhailova, T.A.; Kalugina, O.V.; Shergina, O.V.; Afanasieva, L.V.; Heng, R.K.J.; Majid, N.M.A.; Sase, H. Air pollution monitoring and tree and forest decline in East Asia: A review. Sci. Total Environ. 2020, 742, 140288. [Google Scholar] [CrossRef]
- Tian, Y.H.; Liu, Y.Q.; Wu, X.C.; Yan, M.X.; Liu, F.H.; Luan, J.Y. The Status Quo of Forest Decline and The Impact of Population Competition on Its Growth in The Northeast of The Inner Mongolia. Acta Ecol. Sin. 2022, 42, 7222–7233. [Google Scholar]
- Wu, X.D.; Shao, X.M. Status of dendroclimatological study and its prospects in China. Adv. Earth Sci. 1993, 8, 31–35. [Google Scholar]
- Wang, T.; Bao, A.M.; Xu, W.Q.; Zheng, G.X.; Nzabarinda, V.; Yu, T.; Huang, X.R.; Long, G.; Naibi, S. Dynamics of forest net primary productivity based on tree ring reconstruction in the Tianshan Mountains. Ecol. Indic. 2023, 146, 109713. [Google Scholar] [CrossRef]
- Lambert, J.; Drenou, C.; Denux, J.-P.; Balent, G.; Cheret, V. Monitoring forest decline through remote sensing time series analysis. GIScience Remote Sens. 2013, 50, 437–457. [Google Scholar] [CrossRef]
- Wei, Y.Y.; Sun, S.G.; Liang, D.; Jia, Z.H. Spatial–temporal variations of NDVI and its response to climate in China from 2001 to 2020. Int. J. Digit. Earth 2022, 15, 1463–1484. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sanchez, G.; Penuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef]
- Berner, L.T.; Beck, P.S.A.; Bunn, A.G.; Lloyd, A.H.; Goetz, S.J. High-latitude tree growth and satellite vegetation indices: Correlations and trends in Russia and Canada (1982–2008). J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Camarero, J.J.; Olano, J.M.; Martín-Hernández, N.; Peña-Gallardo, M.; Tomás-Burguera, M.; Gazol, A.; Azorin-Molina, C.; Bhuyan, U.; El Kenawy, A. Diverse relationships between forest growth and the Normalized Difference Vegetation Index at a global scale. Remote Sens. Environ. 2016, 187, 14–29. [Google Scholar] [CrossRef]
- D’Andrea, G.; Šimůnek, V.; Castellaneta, M.; Vacek, Z.; Vacek, S.; Pericolo, O.; Zito, R.G.; Ripullone, F. Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe. Forests 2022, 13, 1417. [Google Scholar] [CrossRef]
- Bunn, A.; Hughes, M.; Kirdyanov, A.V.; Losleben, M.; Shishov, V.; Berner, L.; Olchev, A.; Vaganov, E.A. Comparing forest measurements from tree rings and a space-based index of vegetation activity in Siberia. Environ. Res. Lett. 2013, 8, 035034. [Google Scholar] [CrossRef]
- Coulthard, B.L.; Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Sivrikaya, F. Tree growth and vegetation activity at the ecosystem-scale in the eastern Mediterranean. Environ. Res. Lett. 2017, 12, 084008. [Google Scholar] [CrossRef]
- Zhang, T.W.; Zhang, R.B.; Lu, B.; Mambetov, B.; Kelgenbayev, N.; Dosmanbetov, D.; Maisupova, B.; Chen, F.; Yu, S.L.; Shang, H.M.; et al. Picea schrenkiana tree-ring chronologies development and vegetation index reconstruction for the Alatau Mountains, Central Asia. Geochronometria 2018, 45, 107–118. [Google Scholar] [CrossRef]
- Zhirnova, D.; Belokopytova, L.; Barabantsova, A.; Babushkina, E.; Vaganov, E. What prevails in climatic response of Pinus sylvestris in-between its range limits in mountains: Slope aspect or elevation? Int. J. Biometeorol. 2019, 6, 333–344. [Google Scholar] [CrossRef]
- Correa-Díaz, A.; Silva, L.; Horwath, W.R.; Gomez-Guerrero, A.; Vargas-Hernández, J.; Villanueva-Díaz, J.; Suárez-Espinoza, J.; Velázquez-Martínez, A. From Trees to Ecosystems: Spatiotemporal Scaling of Climatic Impacts on Montane Landscapes Using Dendrochronological, Isotopic, and Remotely Sensed Data. Glob. Biogeochem. Cycles 2020, 34, e2019GB006325. [Google Scholar] [CrossRef]
- Kermavnar, J.; Levanič, T.; Kutnar, L. Stable isotope composition in tree rings of Fagus sylvatica L. saplings reflects environmental variation induced by silviculture and microsite factors. For. Ecol. Manag. 2023, 537, 120949. [Google Scholar] [CrossRef]
- Chen, S.S.; Ma, M.H.; Wu, S.J.; Tang, Q.Q.; Wen, Z.F. Topography intensifies variations in the effect of human activities on forest NPP across altitude and slope gradients. Environ. Dev. 2023, 45, 100826. [Google Scholar] [CrossRef]
- Deng, C.H.; Bai, H.Y.; Gao, S.; Zhao, T.; Ma, X.P. Differences and variations in the elevation-dependent climatic growing season of the northern and southern slopes of the Qinling Mountains of China from 1985 to 2015. Theor. Appl. Climatol. 2019, 137, 1159–1169. [Google Scholar] [CrossRef]
- Wang, Z.; Lyu, L.; Liu, W.; Liang, H.; Huang, J.; Zhang, Q.-B. Topographic patterns of forest decline as detected from tree rings and NDVI. Catena 2021, 198, 105011. [Google Scholar] [CrossRef]
- Wu, X.D. Tree Rings and Climate Change; Meteorological Press: Beijing, China, 1990. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Schweingruber, F.H. Morphological, Anatomical, and Tree-Ring Analytical Characteristics of Trees Frequently Used in Dendrochronology. In Trees and Wood in Dendrochronology; Springer: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Cook, E.R. A Time Series Analysis Approach to Tree-Ring Standardization; University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Vermote, E.; NOAA CDR Program. NOAA Climate Data Record (CDR) of AVHRR Normalized Difference Vegetation Index (NDVI), Version 5; NOAA National Centers for Environmental Information: Boulder, CO, USA, 2019. [Google Scholar] [CrossRef]
- Correa-Díaz, A.; Silva, L.; Horwath, W.; Gomez-Guerrero, A.; Vargas-Hernández, J.; Villanueva-Díaz, J.; Velázquez-Martínez, A.; Suárez-Espinoza, J. Linking Remote Sensing and Dendrochronology to Quantify Climate-Induced Shifts in High-Elevation Forests Over Space and Time. J. Geophys. Res. Biogeosci. 2019, 124, 166–183. [Google Scholar] [CrossRef]
- Karra, K.; Kontgis, C.; Statman-Weil, Z.; Mazzariello, J.C.; Mathis, M.; Brumby, S.P. Global land use/land cover with Sentinel 2 and deep learning. In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium IGARSS, Brussels, Belgium, 11–16 July 2021; pp. 4704–4707. [Google Scholar]
- Singh, R.; Kumar, P.; Pandey, R.; Bala, N. Forest disturbance detection in Garhwal Himalayas using MODIS NDVI time-series and BFAST model. Geocarto Int. 2022, 37, 12689–12708. [Google Scholar] [CrossRef]
- Li, M.Q.; Im, J.H.; Beier, C.L. Machine learning approaches for forest classification and change analysis using multi-temporal Landsat TM images over Huntington Wildlife Forest. GIScience Remote Sens. 2013, 50, 361–384. [Google Scholar] [CrossRef]
- Saito, H.; Nakayama, D.; Matsuyama, H. Comparison of landslide susceptibility based on a decision-tree model and actual landslide occurrence: The Akaishi Mountains, Japan. Geomorphology 2009, 109, 108–121. [Google Scholar] [CrossRef]
- Febriantono, M.; Hadi Pramono, S.; Rahmadwati, R.; Naghdy, G. Classification of multiclass imbalanced data using cost-sensitive decision tree C5.0. IAES Int. J. Artif. Intell. 2020, 9, 65. [Google Scholar] [CrossRef]
- Tanyu, B.F.; Abbaspour, A.; Alimohammadlou, Y.; Tecuci, G. Landslide susceptibility analyses using Random Forest, C4.5, and C5.0 with balanced and unbalanced datasets. Catena 2021, 203, 105355. [Google Scholar] [CrossRef]
- Salzberg, S.L. C4.5: Programs for Machine Learning by J. Ross Quinlan. Morgan Kaufmann Publishers, Inc., 1993. Mach. Learn. 1994, 16, 235–240. [Google Scholar] [CrossRef]
- Yu, L.; Wu, T.J. Assemble learning: A survey of boosting algorithms. Pattern Recognit. Artif. Intell. 2004, 17, 52–59. [Google Scholar]
- Lantz, B. Machine Learning with R; Packt Pub Ltd.: Birmingham, UK, 2013. [Google Scholar]
- Sun, Y.; Wang, L.L.; Chen, J.; Duan, J.P. Reconstructing mean maximum temperatures of May–August from tree-ring maximum density in North DaXing’an Mountains, China. Chin. Sci. Bull. 2012, 57, 1791–1799. [Google Scholar] [CrossRef]
- Farooq, T.; Yasmeen, S.; Shakoor, A.; Nawaz, M.; Rashid, M.; Ahmad, S.; Rasheed, M.; Li, H.; Li, Q. Xylem anatomical responses of Larix Gmelinii and Pinus Sylvestris influenced by the climate of Daxing’an mountains in Northeastern China. Front. Plant Sci. 2023, 14, 1095888. [Google Scholar] [CrossRef]
- Wang, S.W.; Ye, J.L.; Gong, D.Y.; Zhu, J.H.; Yao, T.D.; Zhu, J.H.; Yao, T.D. Establishment of annual temperature series in China in the last 100 years. Q. J. Appl. Meteorol. 1998, 9, 392–401. [Google Scholar]
- Jiang, Y.; Liu, C.; Zhang, J.; Han, S.; Coombs, C.; Wang, X.; Wang, J.; Hao, L.; Dong, S. Tree ring width-based January–March mean minimum temperature reconstruction from Larix gmelinii in the Greater Khingan Mountains, China since A.D. 1765. Int. J. Climatol. 2020, 41, E842–E854. [Google Scholar] [CrossRef]
- Gai, X.R.; Yu, D.P.; Wang, S.L.; Jia, X.; Wu, J.; Zhou, W.M.; Zhou, L.; Dai, L.M. A review of formation mechanism on the ‘divergence problem’ of tree growth-climate relationship. Chin. J. Ecol. 2017, 36, 3273–3280. [Google Scholar] [CrossRef]
- Yasmeen, S.; Wang, X.; Zhao, H.; Zhu, L.; Yuan, D.; Li, Z.; Zhang, Y.; Ahmad, S.; Han, S. Contrasting climate-growth relationship between Larix gmelinii and Pinus sylvestris var. mongolica along a latitudinal gradient in Daxing’an Mountains, China. Dendrochronologia 2019, 58, 125645. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Zhu, X.; Yue, Y.; Xue, J.; Shi, L. How permafrost degradation threatens boreal forest growth on its southern margin? Sci. Total Environ. 2021, 762, 143154. [Google Scholar] [CrossRef]
- Jing, M.; Zhu, L.; Cherubini, P.; Yuan, D.; Li, Z.; Wang, X.; Liu, S. Responses of radial growth of Pinus massoniana and Castanopsis eyrei to climate change at different elevations in south China. Ecol. Indic. 2022, 145, 109602. [Google Scholar] [CrossRef]
- Zheng, P.F.; Wang, D.D.; Jia, G.D.; Yu, X.X.; Liu, Z.Q.; Wang, Y.S.; Zhang, Y. Variation in water supply leads to different responses of tree growth to warming. For. Ecosyst. 2022, 9, 100003. [Google Scholar] [CrossRef]
- Guo, X.M.; Wang, Z.P.; Zhang, N.; Zhang, D.Y. Responses of radial growth of Pinus sylvestris var. mongolica and Larix gmelinii to climate change. Chin. J. Appl. Ecol. 2021, 32, 3405–3414. [Google Scholar] [CrossRef]
- Zhang, J.S.; Lun, B.P.; Shen, J.; Cao, Y. The characteristics of forest diseases, insects and rodents in forest areas and their mitigation countermeasures were analyzed. Inn. Mong. For. Sci. Technol. 2001, 46–48+50. [Google Scholar]
- Sun, Y.F.; Piao, Z.H.; Zhang, Y.W.; Ha, S.B.G. Hazards of Forest Pests and Diseases in Hulunbuir City and Countermeasures for Prevention and Control. Inn. Mong. For. 2003, 33–34. [Google Scholar]
- Koprowski, M.; Duncker, P. Tree ring width and wood density as the indicators of climatic factors and insect outbreaks affecting spruce growth. Ecol. Indic. 2012, 23, 332–337. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.; Gessler, A. Physiological Responses of Forest Trees to Heat and Drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Zhu, X.F. Differential analysis of vegetation response to extreme climate in different vegetation regions of China. Acta Ecol. Sin. 2023, 43, 10202–10215. [Google Scholar] [CrossRef]
- Davis, T.S.; Meddens, A.J.H.; Stevens-Rumann, C.S.; Jansen, V.S.; Sibold, J.S.; Battaglia, M.A. Monitoring resistance and resilience using carbon trajectories: Analysis of forest management-disturbance interactions. Ecol. Appl. A Publ. Ecol. Soc. Am. 2022, 32, e2704. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yong, G.W.; Maya, P.; Zheng, N.; Li, W.; Fang, C.; Chu, Q.Y.; Christian, S.J. Human fingerprint on structural density of forests globally. Nat. Sustain. 2023, 6, 368–379. [Google Scholar] [CrossRef]
- Matskovsky, V.; Venegas-González, A.; Garreaud, R.; Roig, F.A.; Gutiérrez, A.G.; Muñoz, A.A.; Le Quesne, C.; Klock, K.; Canales, C. Tree growth decline as a response to projected climate change in the 21st century in Mediterranean mountain forests of Chile. Glob. Planet. Chang. 2021, 198, 103406. [Google Scholar] [CrossRef]
- Wang, H.; Muller, J.D.; Tatarinov, F.; Yakir, D.; Rotenberg, E. Disentangling Soil, Shade, and Tree Canopy Contributions to Mixed Satellite Vegetation Indices in a Sparse Dry Forest. Remote Sens. 2022, 14, 681. [Google Scholar] [CrossRef]
- Wang, Y.K.; Song, J. Field-Measured Hydraulic Traits and Remotely Sensed NDVI of Four Subtropical Tree Species Showed Transient Declines during the Drought–Heatwave Event. Forests 2023, 14, 1420. [Google Scholar] [CrossRef]
- Qi, Y. The effect of climate change on vegetation at high latitudes of the northern Hemisphere: Afunctional analysis. Acta Ecol. Sin. 1999, 151, 474–477. [Google Scholar]
- Ma, J.N.; Zhang, C.; Hao, G.; Chen, W.L.; Yun, W.J.; Gao, L.L.; Wang, H. Analyzing Ecological Vulnerability and Vegetation Phenology Response Using NDVI Time Series Data and the BFAST Algorithm. Remote Sens. 2020, 12, 3371. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Martín-Hernández, N.; Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R.; Peña-Gallardo, M.; El Kenawy, A.; Domínguez-Castro, F.; Tomas-Burguera, M.; Gutiérrez, E.; et al. Linking tree-ring growth and satellite-derived gross primary growth in multiple forest biomes. Temporal-scale matters. Ecol. Indic. 2020, 108, 105753. [Google Scholar] [CrossRef]
- Bao, G.; Liu, Z.Y.; Liu, N.; Wu, M.L. Simulation analysis of the radial growth characteristics of Pinus sylvestris var. mongolica inHulunbuir Sandy Land by Vaganov-Shashkin Model. Chin. J. Appl. Ecol. 2021, 32, 3448–3458. [Google Scholar] [CrossRef]
- Feng, Y.L.; Wang, W.Z.; Ao, H. Drought Resistance of Larix olgensis Henry and Pinus sylvestris var. mongolica etc. J. Northeast. For. Univ. 1998, 26, 16–20. [Google Scholar]
- Kooch, Y.; Parsapour, M.K.; Nouraei, A.; Kartalaei, Z.M.; Wu, D.; Gómez-Brandón, M.; Lucas-Borja, M.E. The effect of silvicultural systems on soil function depends on bedrock geology and altitude. J. Environ. Manag. 2023, 345, 118657. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Altitude/m | Confidence Year | Mean Sensitivity | Standard Deviation | Within-Trees Rbar | First-Order Autocorrelation | Signal-to-Noise Ratio |
|---|---|---|---|---|---|---|---|
| PS-A | 1100–1200 | 1910–2021 | 0.115 | 0.214 | 0.844 | 0.957 | 101.910 |
| PS-B | 800–900 | 1814–2021 | 0.123 | 0.279 | 0.670 | 0.802 | 25.359 |
| PS-C | 700–800 | 1809–2021 | 0.232 | 0.169 | 0.659 | 0.502 | 32.300 |
| LG-A | 1200–1300 | 1905–2021 | 0.197 | 0.172 | 0.663 | 0.604 | 31.225 |
| LG-B | 1100–1200 | 1759–2021 | 0.155 | 0.211 | 0.626 | 0.770 | 26.870 |
| LG-C | 800–900 | 1904–2021 | 0.191 | 0.211 | 0.649 | 0.379 | 22.950 |
| Principal Component | Initial Eigenvalue Variance (%) | Cumulative (%) | LG-A | LG-B | LG-C | PS-A | PS-B | PS-C |
|---|---|---|---|---|---|---|---|---|
| PC1 | 37.706 | 37.706 | 0.320 | 0.839 | 0.548 | 0.685 | 0.823 | 0.100 |
| PC2 | 33.566 | 71.272 | 0.887 | 0.238 | 0.695 | 0.626 | 0.505 | 0.202 |
| PC3 | 18.751 | 90.023 | 0.055 | 0.172 | 0.258 | 0.258 | 0.146 | 0.961 |
| Location | LG-A | LG-B | LG-C | PS-A | PS-B | PS-C |
|---|---|---|---|---|---|---|
| LG-A | 1 | 0.438 ** | 0.710 ** | −0.29 | −0.165 | −0.18 |
| LG-B | 1 | 0.472 ** | 0.393 * | 0.485 ** | −0.081 | |
| LG-C | 1 | −0.095 | 0.149 | 0.108 | ||
| PS-A | 1 | 0.836 ** | −0.084 | |||
| PS-B | 1 | 0.305 | ||||
| PS-C | 1 |
| Predict | Error | |||
|---|---|---|---|---|
| Decline Risk | Non-Decline Risk | |||
| Actual | Decline risk | 75,776 | 42,848 | 0.36 |
| Non-Decline risk | 189,894 | 958,329 | 0.13 | |
| Error | 0.71 | 0.04 | 0.16 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Wang, Z.; Zhang, D.; Li, L.; Zhao, Y.; Luo, T.; Wang, X. Spatial and Temporal Variation in Primary Forest Growth in the Northern Daxing’an Mountains Based on Tree-Ring and NDVI Data. Forests 2024, 15, 317. https://doi.org/10.3390/f15020317
Wang B, Wang Z, Zhang D, Li L, Zhao Y, Luo T, Wang X. Spatial and Temporal Variation in Primary Forest Growth in the Northern Daxing’an Mountains Based on Tree-Ring and NDVI Data. Forests. 2024; 15(2):317. https://doi.org/10.3390/f15020317
Chicago/Turabian StyleWang, Bing, Zhaopeng Wang, Dongyou Zhang, Linlin Li, Yueru Zhao, Taoran Luo, and Xinrui Wang. 2024. "Spatial and Temporal Variation in Primary Forest Growth in the Northern Daxing’an Mountains Based on Tree-Ring and NDVI Data" Forests 15, no. 2: 317. https://doi.org/10.3390/f15020317