


Potential Distribution and Identification of Critical Areas for the Preservation and Recovery of Three Species of Cinchona L. (Rubiaceae) in Northeastern Peru

 , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
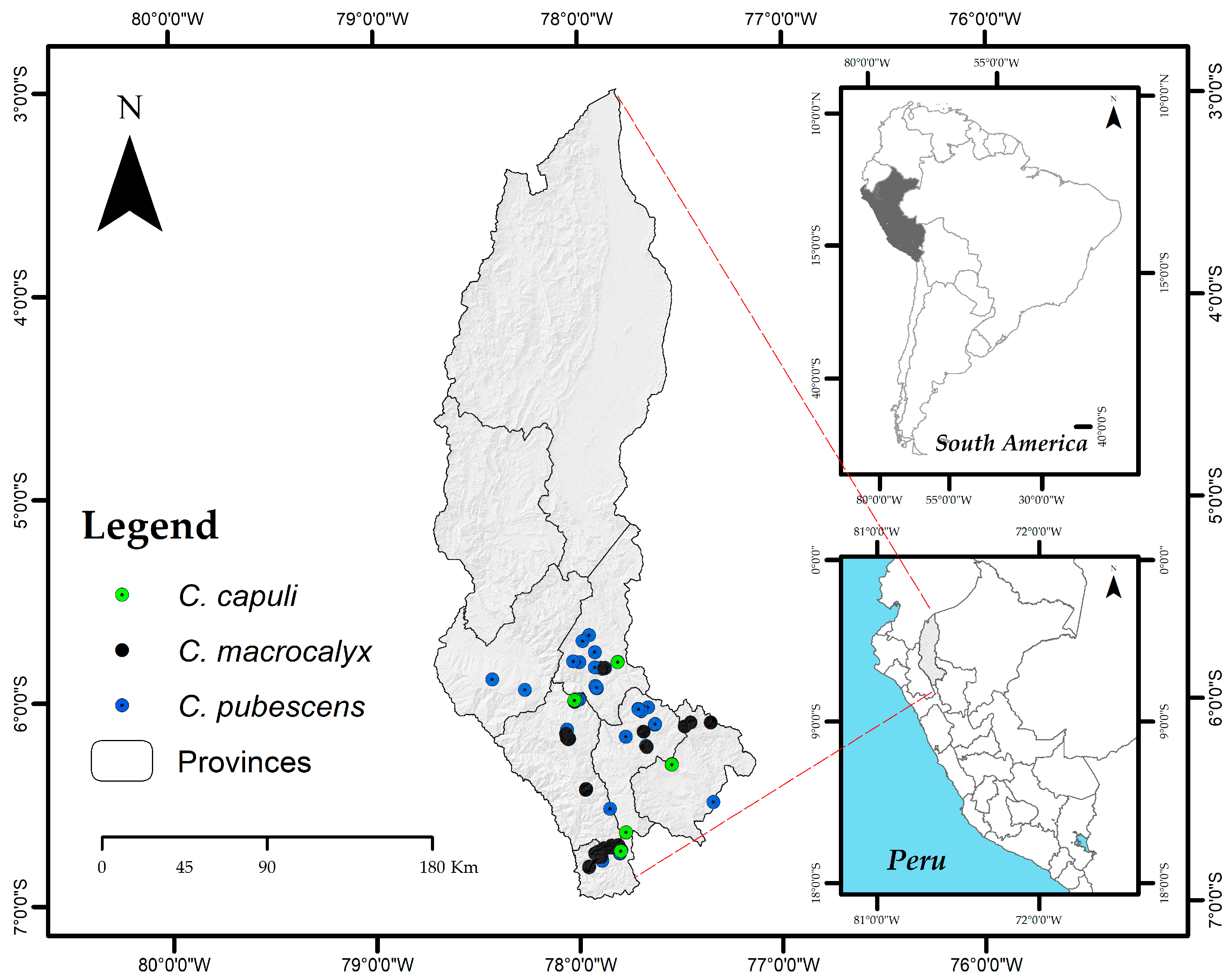
2.2. Records of Cinchona Species
2.3. Bioclimatic, Environmental, Topographic, and Edaphic Variables
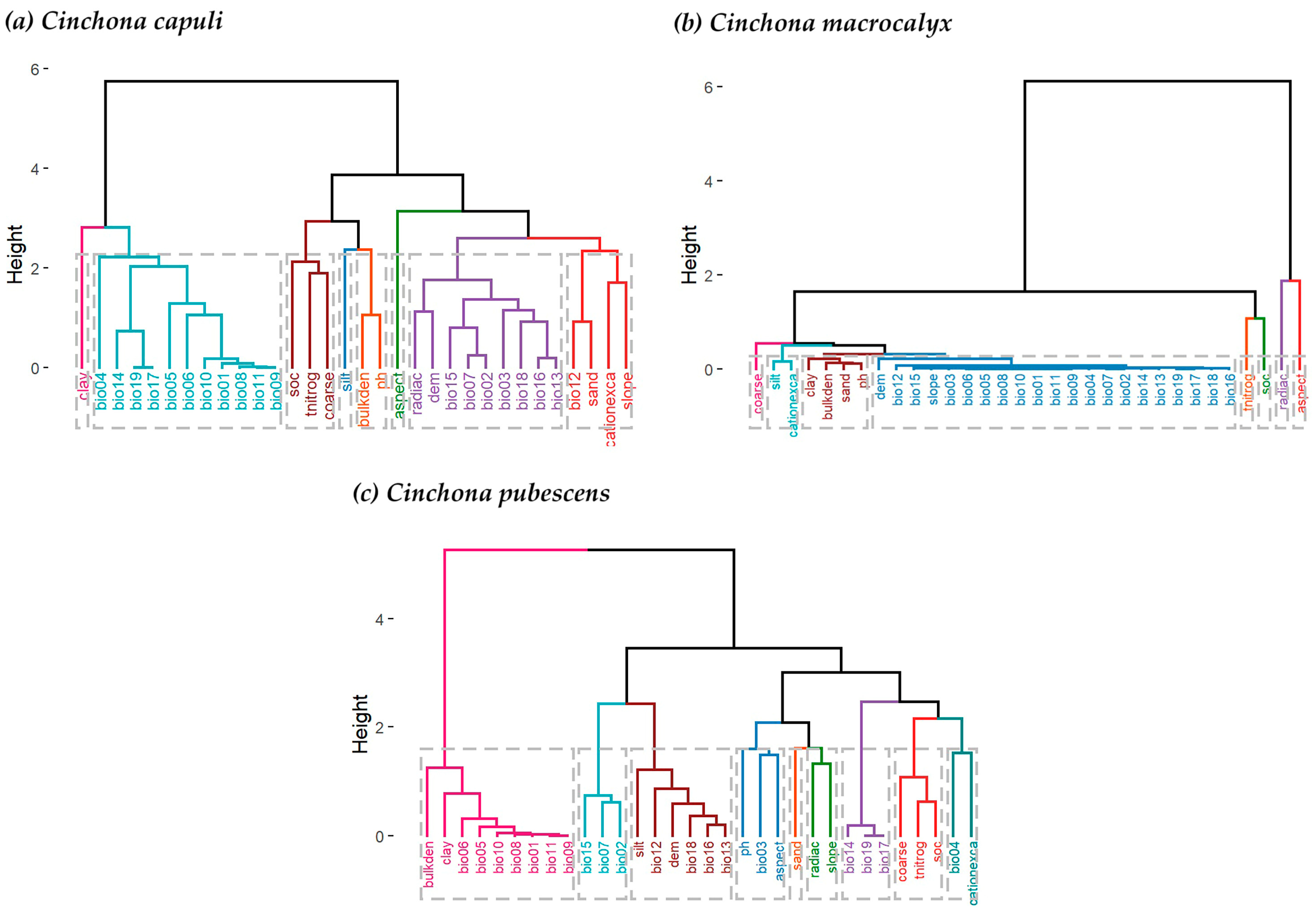
2.4. Variable Selection
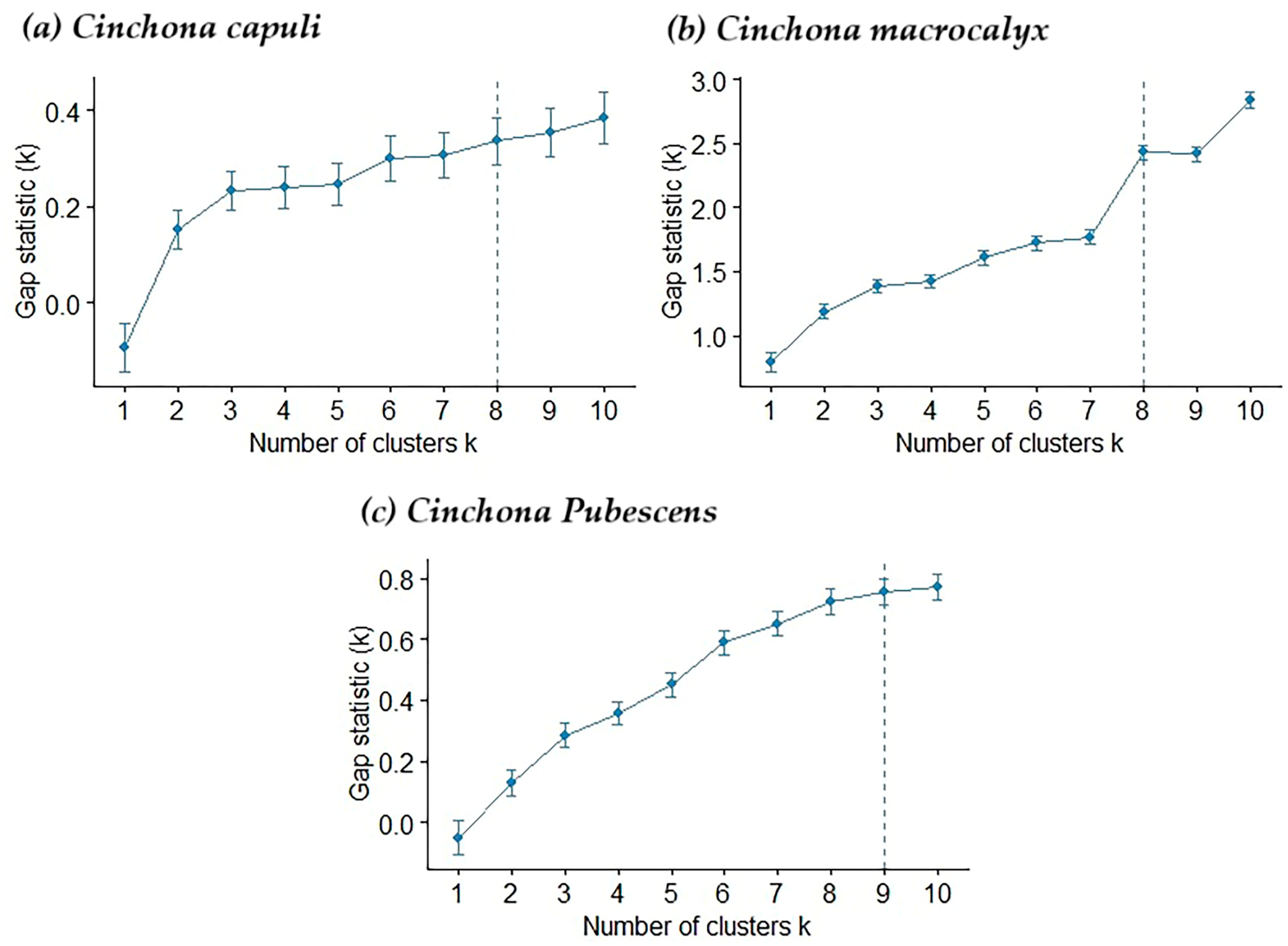
2.5. Model Construction
2.6. Conservation and Recovery of Cinchona Species
3. Results
3.1. Contribution of the Variables
3.2. Performance of the Distribution Pattern of the Model
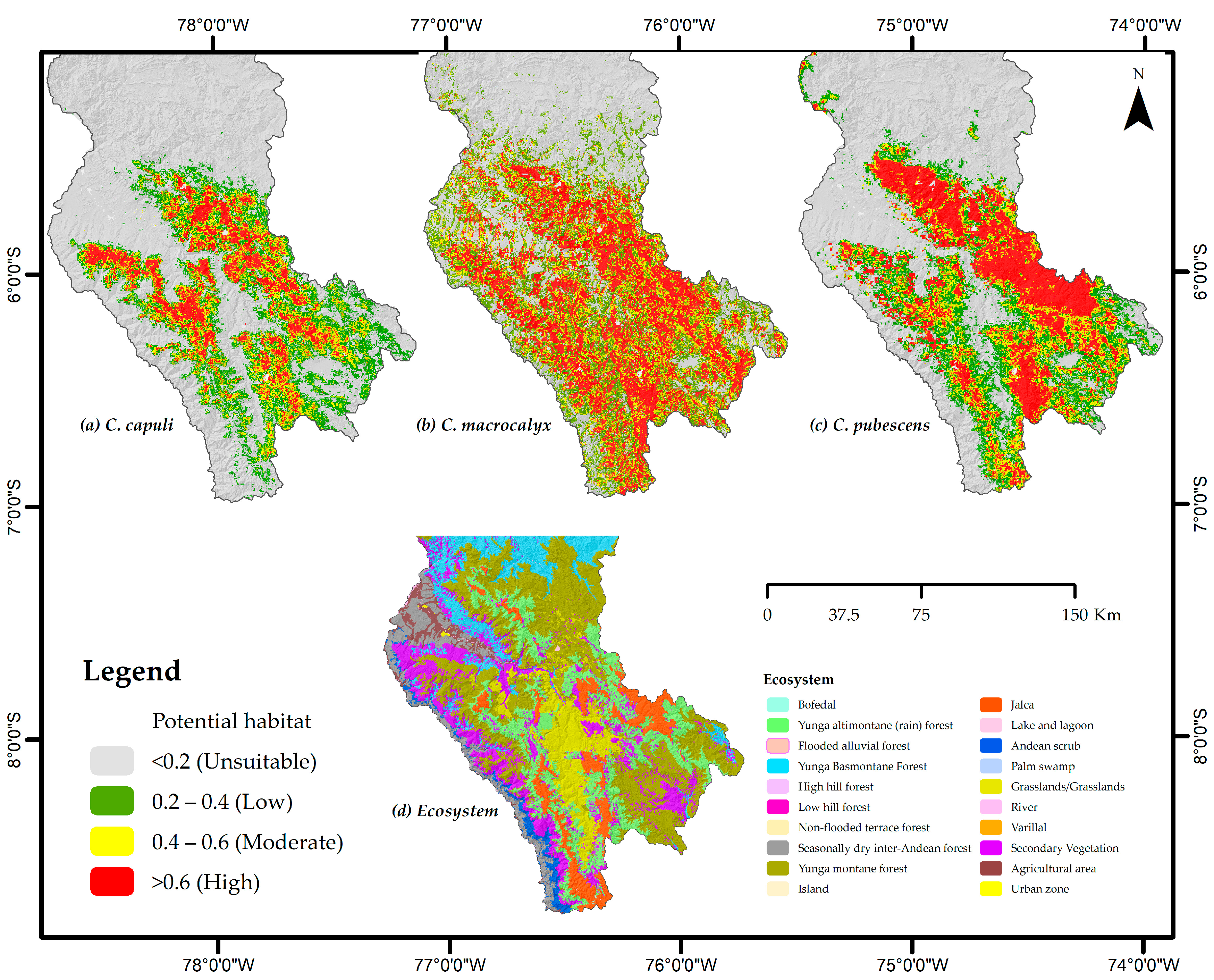
3.3. Occurrence of Cinchona Species
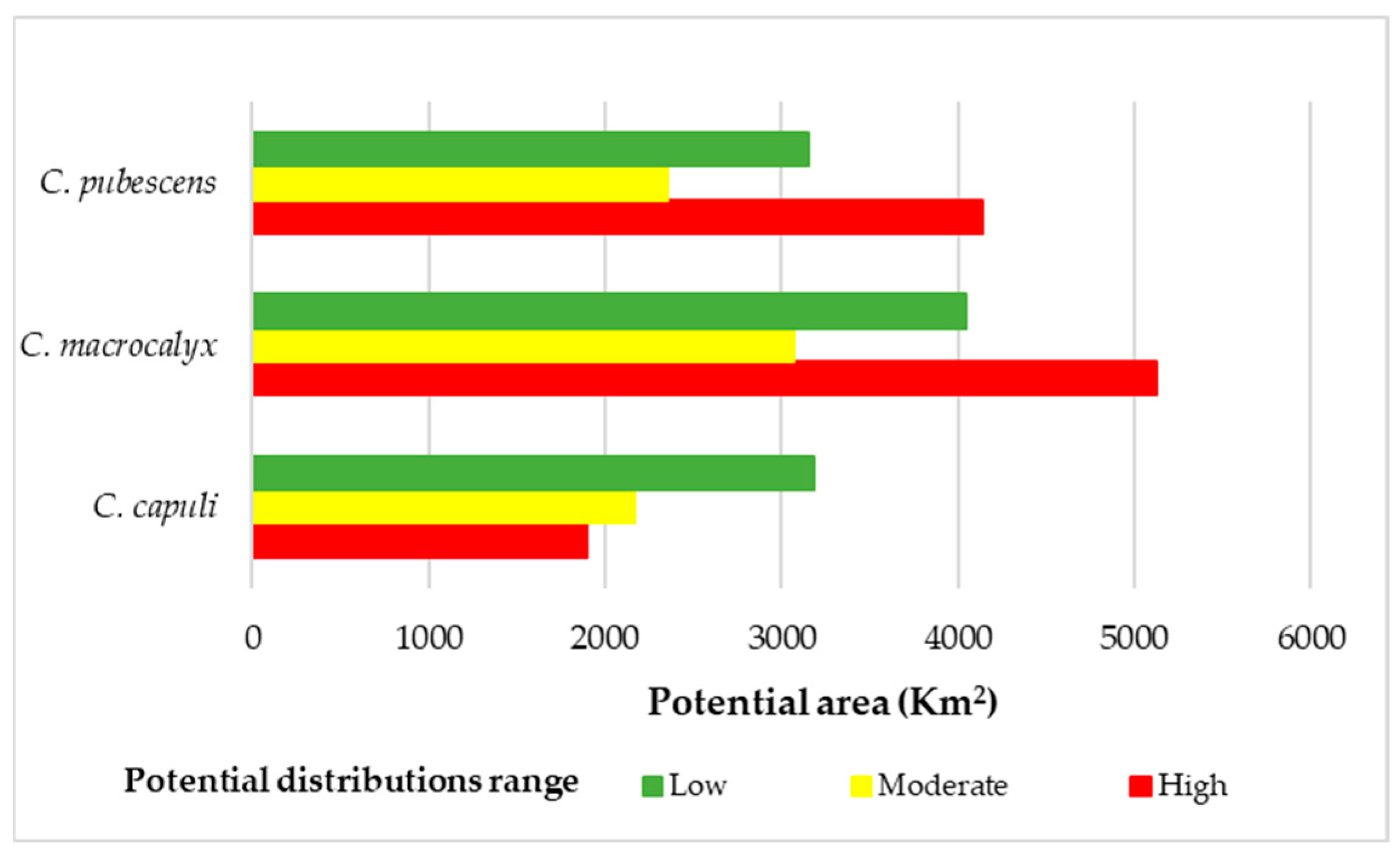
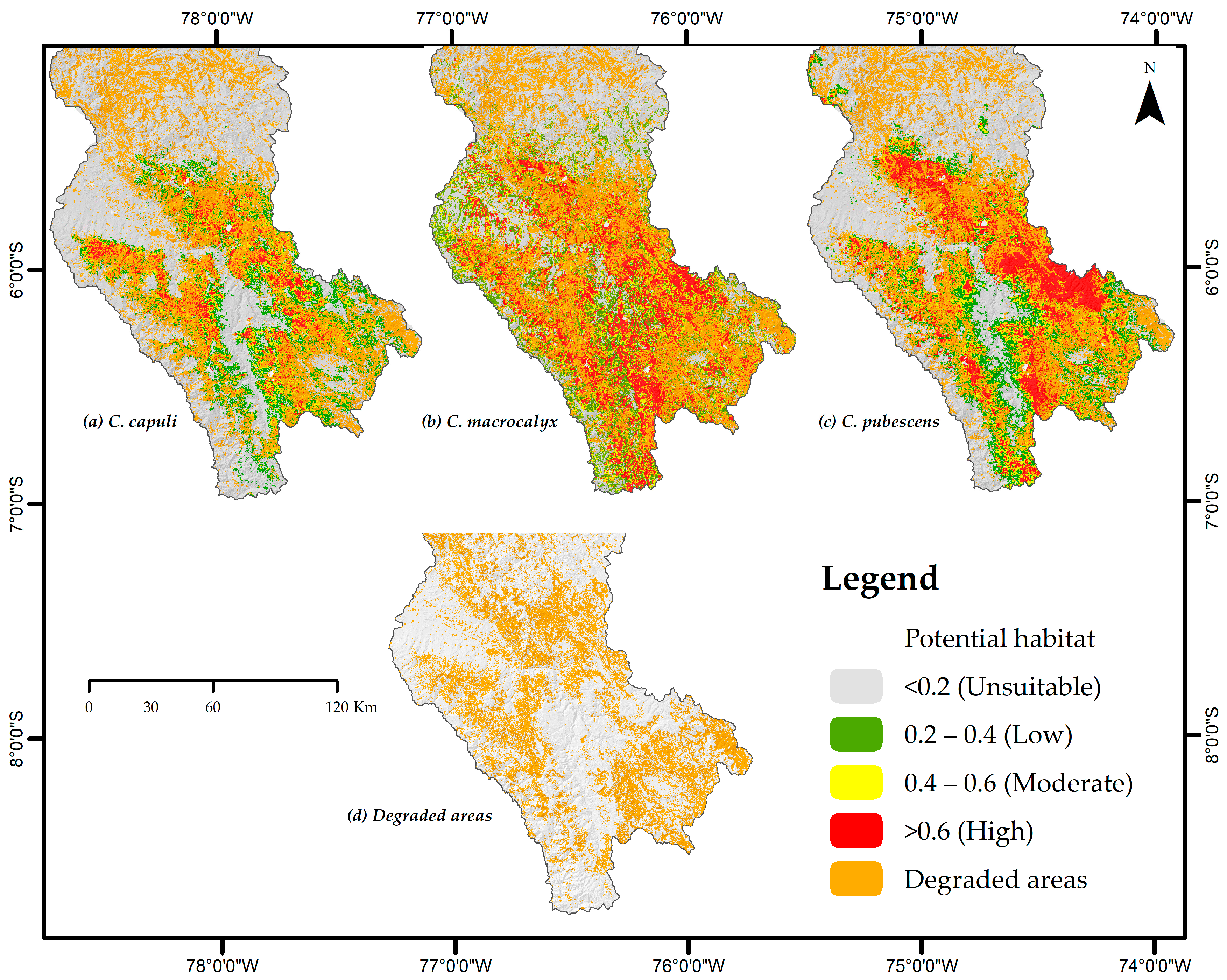
3.4. Conservation of Cinchona Species and Recovery Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, K.R.; León, B. Biodiversity conservation in Peru’s eastern montane forests. Mt. Res. Dev. 2000, 20, 208–211. [Google Scholar] [CrossRef]
- Marian, F.; Ramírez Castillo, P.; Iñiguez Armijos, C.; Günter, S.; Maraun, M.; Scheu, S. Conversion of Andean montane forests into plantations: Effects on soil characteristics, microorganisms, and microarthropods. Biotropica 2020, 52, 1142–1154. [Google Scholar] [CrossRef]
- Souza, C.R.; Mariano, R.F.; Maia, V.A.; Pompeu, P.V.; Dos Santos, R.M.; Fontes, M.A.L. Carbon stock and uptake in the high-elevation tropical montane forests of the threatened Atlantic Forest hotspot: Ecosystem function and effects of elevation variation. Sci. Total Environ. 2023, 882, 163503. [Google Scholar] [CrossRef]
- Bruijnzeel, L.A. Hydrological Impacts of Converting Tropical Montane Cloud Forest to Pasture, with Initial Reference to Northern Costa Rica. Final Technical Report DFID-FRP Project No. R7991; Vrije Universiteit Amsterdam: Amsterdam, The Netherlands, 2006; pp. 2–4. [Google Scholar]
- Osorio, A.; Bahamon, C. Dinámica de la Humedad del Suelo en Bosque alto Andino en el Páramo de Guerrero, Cundinamarca-Colombia. Master’s Thesis, Universidad Nacional de Colombia, Medellín, Colombia, 2008. [Google Scholar]
- Tobón, C.; Bruijnzeel, L.A.; Frumau, A. Physical and hydraulic properties of Tropical Montane Cloud Forest soils and their changes after conversion to pasture. In Proceedings of the Second International Symposium: Science for Conserving and Managing Tropical Montane Cloud Forests, Waimea, HI, USA, July 27–1 August 2004. [Google Scholar]
- Leigh, E.G. Tropical Forest Ecology: A View from Barro Colorado Island; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Brummitt, N.; Lughadha, E.N. Biodiversity: Where’s hot and where’s not. Conserv. Biol. 2003, 17, 1442–1448. Available online: https://www.jstor.org/stable/3588969 (accessed on 4 September 2023). [CrossRef]
- Barthlott, W.; Mutke, J.; Rafiqpoor, M.D.; Kier, G.; Kreft, H. Global centres of vascular plant diversity. Nova Acta Leopold 2005, 92, 61–83. [Google Scholar]
- Rands, M.R.; Adams, W.M.; Bennun, L.; Butchart, S.H.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Aymard, G.A. Breve reseña de los aspectos taxonómicos y nomenclaturales actuales del género Cinchona (Rubiaceae-Cinchoneae). Rev. Acad. Colomb. Cienc. Exactas Físicas Nat. 2019, 43, 234–241. [Google Scholar] [CrossRef]
- Garmendia, A. El Árbol de la Quina (Cinchona spp.), Distribución, Caracterización de su Hábitat y Arquitectura; Universidad Técnica Particular de Loja: Loja, Ecuador, 2005. [Google Scholar]
- Huamán, L.K. Evaluación Morfológica y del Estado de Conservación de Seis Especies del Género Cinchona L. (Rubiaceae) en los Andes del Norte y Centro de Perú. Master’s Thesis, Universidad Nacional Mayor de San Marcos, Lima, Perú, 2020. [Google Scholar]
- Andersson, L. A revision of the genus Cinchona (Rubiaceae-Cinchoneae). Mem. N. Y. Bot. Gard. 1998, 80, 1–75. [Google Scholar]
- Huamán, L.; Albán, J.; Chilquillo, E. Aspectos taxonómicos y avances en el conocimiento del estado actual del árbol de la Quina (Cinchona officinalis L.) en el Norte de Perú. Ecol. Apl. 2019, 18, 145–153. [Google Scholar] [CrossRef]
- Sánchez, T.; Silva, G.; Chichipe, A.K.; Trigoso, M.; Bobadilla, L.G.; Jiménez, G.Y. Influencia de Sustratos y Concentraciones de ácido indol butírico en la Propagación Vegetativa de Cinchona officinalis L.(quina) en Amazonas, Perú. Rev. Cuba. Cienc. For. 2020, 8, 241–251. Available online: http://scielo.sld.cu/pdf/cfp/v8n2/2310-3469-cfp-8-02-241.pdf (accessed on 8 September 2023).
- Castañeda, J.H.V.; Sabaleta, E.L.; Hernández, M.K.B.; Meza, S.N.V.; Sánchez, L.M.Q. Comparación de sustratos en la propagación sexual y asexual del árbol de la quina (Cinchona officinalis). Rev. Investig. Agroprod. Sustentable 2019, 2, 77–85. [Google Scholar] [CrossRef]
- Perez Ocampo, J. Diversidad y Distribución de las Quinas (Cinchona, Rubiaceae) Usando DNA Barcoding para su Conservación en Amazonas. Master’s Thesis, Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas, Chachapoyas, Perú, 2021. [Google Scholar]
- Huamán Vela, M.H. Efecto de Reguladores de Crecimiento y Sustratos en la Propagación Vegetativa de Cinchona pubescens Vahl (Quina) en Microtunel, Amazonas, Perú. Bachelor’s Thesis, Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas, Chachapoyas, Perú, 2023. [Google Scholar]
- García, L.; Veneros, J.; Chavez, S.G.; Oliva, M.; Rojas-Briceno, N.B. World historical mapping and potential distribution of Cinchona spp. in Peru as a contribution for its restoration and conservation. J. Nat. Conserv. 2022, 70, 126290. [Google Scholar] [CrossRef]
- Vargas Aguilar, C.A. Mapas de Distribución Geográfica Histórica y Actual de Cinchona sp. en la Región Amazonas y el Perú. Bachelor’s Thesis, Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas, Chachapoyas, Perú, 2021. [Google Scholar]
- Vergara, A.J.; Cieza-Tarrillo, D.; Ocaña, C.; Quiñonez, L.; Idrogo-Vasquez, G.; Muñoz-Astecker, L.D.; Auquiñivin-Silva, E.A.; Cruzalegui, R.J.; Arbizu, C.I. Current and Future Spatial Distribution of the Genus Cinchona in Peru: Opportunities for Conservation in the Face of Climate Change. Sustainability 2023, 15, 14109. [Google Scholar] [CrossRef]
- Gómez-Díaz, J.A.; Baena, M.L.; González-Zamora, A.; Delfín-Alfonso, C.A. Potential present and future distributions of the genus Atta of Mexico. PLoS ONE 2023, 18, e0292072. [Google Scholar] [CrossRef]
- Tafesse, B.; Bekele, T.; Demissew, S.; Dullo, B.W.; Nemomissa, S.; Chala, D. Conservation implications of mapping the potential distribution of an Ethiopian endemic versatile medicinal plant, Echinops kebericho Mesfin. Ecol. Evol. 2023, 13, e10061. [Google Scholar] [CrossRef]
- Qi, Y.; Xian, X.; Zhao, H.; Yang, M.; Zhang, Y.; Yu, W.; Liu, W. World Spread of Tropical Soda Apple (Solanum viarum) under Global Change: Historical Reconstruction, Niche Shift, and Potential Geographic Distribution. Biology 2023, 12, 1179. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Carvalheiro, L.G.; Polce, C.; van Loon, E.E.; Raes, N.; Reemer, M.; Biesmeijer, J.C. Fit-for-purpose: Species distribution model performance depends on evaluation criteria–Dutch hoverflies as a case study. PLoS ONE 2013, 8, e63708. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. PeerJ 2018, 6, e4287. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, R.A.D.S.; Nascimento, F.A.O.D.; Pereira, M.A.S.B.F.; Souza, D.P.D.; Fontana, A.P.; Cotarelli, V.M.; Oliveira, M.A.; Moura Júnior, E.G.D. Ampliação do conhecimento biogeográfico de Pleurophora pulchra (Lythraceae) com enfoque em biologia da conservação. Rodriguésia 2019, 70, e02772017. [Google Scholar] [CrossRef]
- Cotrina Sánchez, D.A.; Barboza Castillo, E.; Rojas Briceño, N.B.; Oliva, M.; Torres Guzman, C.; Amasifuen Guerra, C.A.; Bandopadhyay, S. Distribution models of timber species for forest conservation and restoration in the Andean-Amazonian landscape, North of Peru. Sustainability 2020, 12, 7945. [Google Scholar] [CrossRef]
- Rojas Briceño, N.B.; Cotrina Sanchez, D.A.; Barboza Castillo, E.; Barrena Gurbillon, M.A.; Sarmiento, F.O.; Sotomayor, D.A.; Oliva, M.; Salas López, R. Current and future distribution of five timber forest species in Amazonas, Northeast Peru: Contributions towards a restoration strategy. Diversity 2020, 12, 305. [Google Scholar] [CrossRef]
- Song, X.; Jiang, Y.; Zhao, L.; Jin, L.; Yan, C.; Liao, W. Predicting the Potential Distribution of the Szechwan Rat Snake (Euprepiophis perlacea) and Its Response to Climate Change in the Yingjing Area of the Giant Panda National Park. Animals 2023, 13, 3828. [Google Scholar] [CrossRef] [PubMed]
- Driver, S.M.; Eversole, C.B.; Unger, D.R.; Kulhavy, D.L.; Schalk, C.M.; Hung, I.K. Assessing the Impacts of Climate Change on the At-Risk Species Anaxyrus microscaphus (The Arizona Toad): A Local and Range-Wide Habitat Suitability Analysis. Ecologies 2023, 4, 762–778. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Li, M.; Zhang, H. Predicting the Distribution of the Invasive Species Atractosteus spatula, the Alligator Gar, in China. Water 2023, 15, 4291. [Google Scholar] [CrossRef]
- Ouyang, X.; Chen, A.; Li, Y.; Han, X.; Lin, H. Predicting the potential distribution of pine wilt disease in China under climate change. Insects 2022, 13, 1147. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Liu, Y.; Jiang, H.; Chen, X.; Feng, H. Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia. Plants 2023, 12, 3187. [Google Scholar] [CrossRef]
- Vargas, R.; Clima, J. Estudios Temáticos para la Zonificación Ecológica Económica del Departamento de Amazonas; Instituto de Investigaciones de la Amazonía Peruana (IIAP) & Programa de Investigaciones en Cambio Climático, Desarrollo Territorial y Ambiente (PROTERRA): Chachapoyas, Perú, 2010; Volume 1, pp. 1–27. [Google Scholar]
- SERNANP. Áreas Naturales Protegidas de Administración Nacional con Categoría Definitiva; SERNANP: Lima, Perú, 2020. [Google Scholar]
- MINAM. Estudio para la Identificación de Áreas Degradadas y Propuesta de Monitoreo; MINAM: Lima, Perú, 2021; Available online: https://geoservidor.minam.gob.pe/recursos/intercambio-de-datos/ (accessed on 11 September 2023).
- Yonz, J.A.V.; Quispe, W.S.A. Deforestación y pérdida de bosques en la Provincia de Condorcanqui, Amazonas, Perú, durante el período 2001–2021. Ing. Investig. 2023, 5, e790. [Google Scholar] [CrossRef]
- López, R.S.; Calampa, N.J.R.; Castillo, E.B.; Chchiple, M.E.M.; Oliva, M. Microzonificación agroecológica de sistemas agrosilvopastoriles empleando un modelo de procesamiento basado en SIG en parcelas en la provincia de Bongará, Amazonas (Perú). Rev. Investig. Agroprod. Sustentable 2019, 1, 40–50. [Google Scholar] [CrossRef]
- Raymundo Jáuregui, A.A. Evaluación de la Pérdida del Suelo Mediante el Modelo Rusle en la Microcuenca del río Jucusbamba, Provincia de Luya, Región Amazonas (Perú). Master’s Thesis, Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas, Chachapoyas, Perú, 2018. [Google Scholar]
- Shanee, N.; Shanee, S. Land Trafficking, Migration, and Conservation in the “No-Man’s Land” of Northeastern Peru. Trop. Conserv. Sci. 2016, 9, 1940082916682957. [Google Scholar] [CrossRef]
- Salas, L.R.; Barboza, C.E.; Oliva, C.M. Dinámica multitemporal de índices de deforestación en el distrito de Florida, departamento de Amazonas, Perú. Rev. INDES 2014, 2, 18–27. [Google Scholar] [CrossRef]
- Rojas, B.N.B.; Barboza, C.E.; Maicelo, Q.J.L.; Oliva, C.S.M.; Salas, L.R. Deforestación en la Amazonía peruana: Índices de cambios de cobertura y uso del suelo basado en SIG. Boletín Asoc. Geógrafos Españoles 2019, 81, 1–34. [Google Scholar] [CrossRef]
- Mendoza, C.M.E.; Salas, L.R.; Barboza, C.E. Análisis multitemporal de la deforestación usando la clasificación basada en objetos, distrito de Leymebamba (Perú). Rev. INDES 2015, 3, 67–76. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2020; Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 12 September 2023).
- Armas, A.; De Lombaerde, E.; Sanczuk, P.; van Meerbeek, K.; Carón, M.; Honnay, O.; Verbeeck, H.; Duchateau, L.; Schrevens, E.; Verheyen, K.; et al. Distribucion Actual de la Cinchona Pubescens y Cinchona Calisaya y el Impacto Futuro en sus Hábitats a Causa del Cambio Climático. 2022. Available online: https://www.researchgate.net/publication/362610014_Distribucion_actual_de_la_Cinchona_pubescens_y_Cinchona_calisaya_y_el_impacto_futuro_en_sus_habitats_a_causa_del_cambio_climatico (accessed on 15 September 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Meza Mori, G.; Rojas-Briceño, N.B.; Cotrina Sánchez, A.; Oliva-Cruz, M.; Olivera Tarifeño, C.M.; Hoyos Cerna, M.Y.; Ramos Sandoval, J.D.; Torres Guzmán, C. Potential Current and Future Distribution of the Long-Whiskered Owlet (Xenoglaux loweryi) in Amazonas and San Martin, NW Peru. Animals 2022, 12, 1794. [Google Scholar] [CrossRef]
- Tanner, E.P.; Papeş, M.; Elmore, R.D.; Fuhlendorf, S.D.; Davis, C.A. Incorporating abundance information and guiding variable selection for climate-based ensemble forecasting of species’ distributional shifts. PLoS ONE 2017, 12, e0184316. [Google Scholar] [CrossRef]
- Kariyawasam, C.S.; Kumar, L.; Ratnayake, S.S. Invasive plant species establishment and range dynamics in Sri Lanka under climate change. Entropy 2019, 21, 571. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Ren, G.; Zhao, K.; Wang, X. Global potential distribution prediction of Xanthium italicum based on Maxent model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- Marchi, M.; Sinjur, I.; Bozzano, M.; Westergren, M. Evaluating WorldClim Version 1 (1961–1990) as the baseline for sustainable use of forest and environmental resources in a changing climate. Sustainability 2019, 11, 3043. [Google Scholar] [CrossRef]
- Yang, M.; Sun, L.; Yu, Y.; Zhang, H.; Malik, I.; Wistuba, M.; Yu, R. Predicting the Potential Geographical Distribution of Rhodiola L. in China under Climate Change Scenarios. Plants 2023, 12, 3735. [Google Scholar] [CrossRef]
- Meza Mori, G.; Barboza Castillo, E.; Torres Guzmán, C.; Cotrina Sánchez, D.A.; Guzman Valqui, B.K.; Oliva, M.; Bandopadhyay, S.; Salas López, R.; Rojas Briceño, N.B. Predictive modelling of current and future potential distribution of the spectacled bear (Tremarctos ornatus) in Amazonas, northeast Peru. Animals 2020, 10, 1816. [Google Scholar] [CrossRef]
- De Marco, P.; Nóbrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef]
- Brun, P.; Thuiller, W.; Chauvier, Y.; Pellissier, L.; Wüest, R.O.; Wang, Z.; Zimmermann, N.E. Model complexity affects species distribution projections under climate change. J. Biogeogr. 2020, 47, 130–142. [Google Scholar] [CrossRef]
- Bekele, M.; Seifu, A.; Wube, S.; Tilaye, G. Modeling the Current and future potential distribution of Arabis alpina using MaxEnt softwarein selected mountains of Ethiopia. Sustain. Agric. Food Environ. Res. 2023, 12. [Google Scholar] [CrossRef]
- Guzman, B.K.; Cotrina-Sánchez, A.; Allauja-Salazar, E.E.; Olivera Tarifeno, C.M.; Ramos Sandoval, J.D.; Hoyos Cerna, M.Y.; Barboza, E.; Torres Guzmán, C.; Oliva, M. Predicting potential distribution and identifying priority areas for conservation of the Yellow-tailed Woolly Monkey (Lagothrix flavicauda) in Peru. J. Nat. Conserv. 2022, 70, 126302. [Google Scholar] [CrossRef]
- He, Y.; Ma, J.; Chen, G. Potential geographical distribution and its multi-factor analysis of Pinus massoniana in China based on the maxent model. Ecol. Indic. 2023, 154, 110790. [Google Scholar] [CrossRef]
- Zvidzai, M.; Mawere, K.K.; N’andu, R.J.; Ndaimani, H.; Zanamwe, C.; Zengeya, F.M. Application of maximum entropy (MaxEnt) to understand the spatial dimension of human–wildlife conflict (HWC) risk in areas adjacent to Gonarezhou National Park of Zimbabwe. Ecol. Soc. 2023, 28, 18. [Google Scholar] [CrossRef]
- Hentschel, A.E.; Beijert, I.J.; Bosschieter, J.; Kauer, P.C.; Vis, A.N.; Lissenberg-Witte, B.I.; van Moorselaar, R.J.A.; Steenbergen, R.D.M.; Nieuwenhuijzen, J.A. Bladder cancer detection in urine using DNA methylation markers: A technical and prospective preclinical validation. Clin. Epigenet. 2022, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Haq, S.M.; Aziz, R.; Bussmann, R.W.; Ali, K.; Subhan, F.; Jones, D.A.; Zaitouny, A. Potential distribution of a noxious weed (Solanum viarum Du-nal), current status, and future invasion risk based on MaxEnt modeling. Geol. Ecol. Landsc. 2023, 1–16. [Google Scholar] [CrossRef]
- Llatas Mires, E. Modelamiento de la Distribución Natural del Género Cinchona en el norte del Perú. Bachelor’s Thesis, Universidad Nacional Autónoma de Chota, Chota, Perú, 2023. [Google Scholar]
- González-Orozco, C.E.; Guillén, E.G.; Cuvi, N. Changes of Cinchona distribution over the past two centuries in the northern Andes. R. Soc. Open Sci. 2023, 10, 230229. [Google Scholar] [CrossRef]
- Varela, S.; Lobo, J.M.; Hortal, J. Using species distribution models in paleobiogeography: A matter of data, predictors and concepts. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 451–463. [Google Scholar] [CrossRef]
- Varela, S.; Mateo, R.G.; García-Valdés, R.; Fernández-González, F. Macroecología y ecoinformática: Sesgos, errores y predicciones en el modelado de distribuciones. Ecosistemas 2014, 23, 46–53. [Google Scholar] [CrossRef]
- Godsoe, W.; Franklin, J.; Blanchet, F.G. Effects of biotic interactions on modeled species’ distribution can be masked by environmental gradients. Ecol. Evol. 2017, 7, 654–664. [Google Scholar] [CrossRef]
- Cotrina Sanchez, A.; Rojas Briceno, N.B.; Bandopadhyay, S.; Ghosh, S.; Torres Guzman, C.; Oliva, M.; Guzman, B.K.; Salas Lopez, R. Biogeographic distribution of Cedrela spp. genus in Peru using MaxEnt modeling: A conservation and restoration approach. Diversity 2021, 13, 261. [Google Scholar] [CrossRef]
- Gonzalez-Orozco, C. Regiones biogeográficas del género Cinchona L. (Rubiaceae-Cinchoneae). Rev. Noved. Colomb. 2021, 16, 135–156. [Google Scholar] [CrossRef]
- Albán, J. Etnobotánica de Rubiaceae peruanas. Ph.D. Thesis, Universidad Nacional Mayor de San Marcos, Lima, Perú, 2013. [Google Scholar]
- Asicona Caba, P. Evaluación de Cuatro Sustratos en Semilleros de Quina (Cinchona Ledgeriana; Rubiaceae) en Escuintla. Master’s Thesis, Universidad Rafael Landívar, Escuintla, Guatemala, 2013. [Google Scholar]
- Fernandez Zarate, F.H.; Huaccha Castillo, A.E.; Quiñones Huatangari, L.; Seminario Cunya, A.; Vaca Marquina, S.P. Producción del Árbol de la Quina (Cinchona micrantha R. y P.); Universidad Nacional de Jaén: Jaén, Perú, 2022. [Google Scholar]
- Padilla, T. Estudio Fenológico y Análisis de las Características del Suelo Donde se Desarrolla Cinchona officinalis L. Cuatro Relictos Boscosos de la Provincia de Loja. Master’s Thesis, Universidad Nacional de Loja, Loja, Ecuador, 2017. [Google Scholar]
- Monteferri, B.; Cuba, A.; Zari, L.; Grandez, P. Áreas de Conservación Privada en el Perú: Avances y Propuestas a 20 Años de su Creación; Sociedad Peruana de Derecho Ambiental: Lima, Perú, 2019. [Google Scholar]
- Bonn Challenge. A Global Effort to Bring 150 Million Hectares of Deforested and Degraded Land into Restoration by 2020 and 350 Million Hectares by 2030. Available online: https://www.bonnchallenge (accessed on 18 September 2023).
- MINAGRI. Resolución Directoral Ejecutiva N° 067-2020-MINAGRI-DVDIAR-AGRO RURAL-DE: Aprobación del Plan de Acción para el Repoblamiento Forestal con Especies del género cinchona (Árbol de la Quina) 2020–2022. Available online: https://www.gob.pe/institucion/agrorural/normas-legales/2778257-067-2022-midagri-dvdafir-agro-rural-de (accessed on 20 September 2023).
- Albán-Castillo, J.; Chilquillo, E.; Melchor-Castro, B.; Arakaki, M.; León, B.; Suni, M. Cinchona L. “Árbol de la Quina”: Repoblamiento y reforestación en el Perú. Rev. Peru. Biol. 2020, 27, 423–426. [Google Scholar] [CrossRef]
- Mostacero-Leon, J.; Mejia-Coico, F.; Pelaez-Pelaez, F.; Charcape-Ravelo, M. Especies madereras nativas del norte del perú. Rebiol 1998, 16, 67–78. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 21 September 2023).
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined use of systematic conservation planning, species distribution modelling, and connectivity analysis reveals severe conservation gaps in a megadiverse country (Peru). PLoS ONE 2014, 9, e0114367. [Google Scholar] [CrossRef]
- IITO. ITTO Guidelines for the Restoration, Management and Rehabilitation of Degraded and Secondary Tropical Forests; Internaitonal Tropical Timber Organization: Yokohama, Japan, 2002. [Google Scholar]
- Burnett, K.M.; Ticktin, T.; Bremer, L.L.; Quazi, S.A.; Geslani, C.; Wada, C.A.; Kurashima, N.; Mandle, L.; Pascua, P.; Depraetere, T.; et al. Restoring to the future: Environmental, cultural, and management trade-offs in historical versus hybrid restoration of a highly modified ecosystem. Conserv. Lett. 2019, 12, e12606. [Google Scholar] [CrossRef]
- Griscom, B.W.; Adams, J.; Ellis, P.W.; Houghton, R.A.; Lomax, G.; Miteva, D.A.; Schlesinger, W.H.; Shoch, D.; Siikamäki, J.V.; Smith, P.; et al. Natural climate solutions. Proc. Natl. Acad. Sci. USA 2017, 114, 11645–11650. [Google Scholar] [CrossRef]
- Dai, X.; Wu, W.; Ji, L.; Tian, S.; Yang, B.; Guan, B.; Wu, D. MaxEnt model-based prediction of potential distributions of Parnassiawightiana (Celastraceae) in China. Biodivers. Data J. 2022, 10, e81073. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, R.; Zhou, X.; Zhang, X.; Zhao, G.; Zhang, F. Prediction of potential distribution areas and priority protected areas of Agastache rugosa based on Maxent model and Marxan model. Front. Plant Sci. 2023, 14, 1200796. [Google Scholar] [CrossRef]
- Zhao, X.; Zheng, Y.; Wang, W.; Wang, Z.; Zhang, Q.; Liu, J.; Zhang, C. Habitat Suitability Evaluation of Different Forest Species in Lvliang Mountain by Combining Prior Knowledge and MaxEnt Model. Forests 2023, 14, 438. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Symbol | Units |
|---|---|---|
| 1. Environmental variables | ||
| 1.1. Bioclimatic | ||
| Annual Mean Temperature | bio01 | °C |
| Mean Diurnal Range | bio02 | °C |
| Isothermality | bio03 | °C |
| Temperature Seasonality | bio04 | °C |
| Max Temperature of Warmest Month | bio05 | °C |
| Min Temperature of Warmest Month | bio06 | °C |
| Annual Temperature Range | bio07 | °C |
| Mean Temperature of Wettest Quarter | bio08 | °C |
| Mean Temperature of Driest Quarter | bio09 | °C |
| Mean Temperature of Warmest Quarter | bio10 | °C |
| Mean Temperature of Coldest Quarter | bio11 | °C |
| Annual Precipitation | bio12 | Millimeter |
| Precipitation of Wettest Month | bio13 | Millimeter |
| Precipitation of Driest Month | bio14 | Millimeter |
| Precipitation Seasonality | bio15 | Millimeter |
| Precipitation of Wettest Quarter | bio16 | Millimeter |
| Precipitation of Driest Quarter | bio17 | Millimeter |
| Precipitation of Warmest Quarter | bio18 | Millimeter |
| Precipitation of Coldest Quarter | bio19 | Millimeter |
| 1.2. Radiation | radiac | MJ m−2 día−1 |
| 2. Topographies | ||
| Elevation above mean sea level | dem | mals |
| Cardinal orientation of the slope | aspect | ° |
| Slope of the terrain | slope | ° |
| 3. Edaphic variables | ||
| Bulk density of the fine earth fraction | bdod | cg/cm3 |
| Clay content | clay | % |
| Volumetric fraction of coarse fragments | coarse | cm3/dm3 (vol %) |
| Sand content | sand | % |
| Slime content | Silt | % |
| Cation exchange capacity | cec | cmol kg−1 |
| Total nitrogen | nitrog | cg/kg |
| Organic carbon | soc | g kg−1 |
| pH in H2O | pH | pH × 10 |
| Specie | Variable 1 (%) | Variable 2 (%) | Variable 3 (%) | Total Contribution |
|---|---|---|---|---|
| C. capuli | cec (80.2%) | bio03 (13.9%) | bio04 (5.6%) | 99.7% |
| C. macrocalyx | cec (69.2%) | aspect (18.4%) | clay (7.1%) | 94.7% |
| C. pubescens | bio06 (62.4%) | bio14 (20.6%) | cec (8.1%) | 91.1% |
| Specie | C. capuli | C. macrocalyx | C. pubescens |
|---|---|---|---|
| AUC | 0.992 | 0.981 | 0.995 |
| Specie | PNA Modalities | Geographic Area (km2) | Potential Area (km2) | Total (km2) | % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low | % | Moderate | % | High | % | |||||
| C. capuli | NS | 2475.25 | 73.9 | 2.99 | 26.58 | 1.07 | 13.44 | 0.54 | 113.92 | 4.60 |
| RCA | 717.35 | 228.39 | 31.84 | 47.68 | 6.65 | 4.1 | 0.57 | 280.17 | 39.06 | |
| PCA | 1109.25 | 268.73 | 24.23 | 263.12 | 23.72 | 222.09 | 20.02 | 753.89 | 67.96 | |
| RA | 1366.85 | 9.15 | 0.67 | 0 | 0.00 | 0 | 0.00 | 9.15 | 0.67 | |
| BZ | 6668.07 | 348.18 | 5.22 | 211.87 | 3.18 | 131.21 | 1.97 | 691.26 | 10.37 | |
| Total | 12,336.77 | 928.35 | 7.53 | 549.25 | 4.45 | 370.84 | 3.01 | 1848.39 | 14.98 | |
| C. macrocalyx | NS | 2475.25 | 120.04 | 4.85 | 44.84 | 1.81 | 85.57 | 3.46 | 250.45 | 10.12 |
| RCA | 717.35 | 164.18 | 22.89 | 91.26 | 12.72 | 89.09 | 12.42 | 344.53 | 48.03 | |
| PCA | 1109.25 | 235.56 | 21.24 | 259.18 | 23.37 | 471.41 | 42.50 | 963.15 | 86.83 | |
| RA | 1366.85 | 67.77 | 4.96 | 13.26 | 0.97 | 1.25 | 0.09 | 82.28 | 6.02 | |
| BZ | 6668.07 | 331.54 | 4.97 | 214.87 | 3.22 | 386.66 | 5.80 | 932.54 | 13.99 | |
| Total | 12,336.77 | 919.09 | 7.45 | 623.41 | 5.05 | 1033.98 | 8.38 | 2572.95 | 20.86 | |
| C. pubescens | NS | 2475.25 | 75.48 | 3.05 | 38.54 | 1.56 | 140.65 | 5.68 | 254.67 | 10.29 |
| RCA | 717.35 | 199.31 | 27.78 | 114.23 | 15.92 | 92.83 | 12.94 | 406.37 | 56.65 | |
| PCA | 1109.25 | 167.62 | 15.11 | 227.66 | 20.52 | 561.26 | 50.60 | 956.24 | 86.21 | |
| RA | 1366.85 | 9.62 | 0.70 | 2.05 | 0.15 | 1.03 | 0.08 | 12.7 | 0.93 | |
| BZ | 6668.07 | 161.85 | 2.43 | 153.56 | 2.30 | 596.47 | 8.95 | 911.88 | 13.68 | |
| Total | 12,336.77 | 613.88 | 4.98 | 536.04 | 4.35 | 1251.59 | 10.15 | 2541.86 | 20.60 | |
| Specie | Degraded Areas | Geographic Area (km2) | Potential Area (km2) | Total | % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low | % | Moderate | % | High | % | (km2) | ||||
| C. capuli | Department of Amazonas | 10,987.22 | 1203.47 | 10.95 | 1052.31 | 9.58 | 1018.42 | 9.27 | 3274.2 | 29.80 |
| C. macrocalyx | 1232.15 | 11.21 | 1059.99 | 9.65 | 1962.38 | 17.86 | 4254.52 | 38.72 | ||
| C. pubescens | 1118.52 | 10.18 | 987.98 | 8.99 | 1719.69 | 15.65 | 3826.19 | 34.82 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coronel-Castro, E.; Meza-Mori, G.; Torres, J.M.C.; Mondragón, E.P.; Cotrina-Sanchez, A.; Oliva Cruz, M.; López, R.S.; Campo Ramos, R.E. Potential Distribution and Identification of Critical Areas for the Preservation and Recovery of Three Species of Cinchona L. (Rubiaceae) in Northeastern Peru. Forests 2024, 15, 321. https://doi.org/10.3390/f15020321
Coronel-Castro E, Meza-Mori G, Torres JMC, Mondragón EP, Cotrina-Sanchez A, Oliva Cruz M, López RS, Campo Ramos RE. Potential Distribution and Identification of Critical Areas for the Preservation and Recovery of Three Species of Cinchona L. (Rubiaceae) in Northeastern Peru. Forests. 2024; 15(2):321. https://doi.org/10.3390/f15020321
Chicago/Turabian StyleCoronel-Castro, Elver, Gerson Meza-Mori, Jose M. Camarena Torres, Elí Pariente Mondragón, Alexander Cotrina-Sanchez, Manuel Oliva Cruz, Rolando Salas López, and Ricardo E. Campo Ramos. 2024. "Potential Distribution and Identification of Critical Areas for the Preservation and Recovery of Three Species of Cinchona L. (Rubiaceae) in Northeastern Peru" Forests 15, no. 2: 321. https://doi.org/10.3390/f15020321
APA StyleCoronel-Castro, E., Meza-Mori, G., Torres, J. M. C., Mondragón, E. P., Cotrina-Sanchez, A., Oliva Cruz, M., López, R. S., & Campo Ramos, R. E. (2024). Potential Distribution and Identification of Critical Areas for the Preservation and Recovery of Three Species of Cinchona L. (Rubiaceae) in Northeastern Peru. Forests, 15(2), 321. https://doi.org/10.3390/f15020321