1. Introduction
The planting of Scots pine (
Pinus sylvestris L.) and Norway spruce (
Picea abies (L.) Karst) seedlings is currently the most common regeneration practice undertaken in Swedish production forests [
1]. The majority of planted conifer seedlings are grown in nurseries in container systems of different sizes and types. The choice of seedling size depends on different priorities decided by managers. A newly planted seedling is vulnerable to both water stress and nutrient stress since its root soil contact is limited [
2]. Regardless of size, seedlings with a low shoot–root ratio establish faster and tolerate drought stress better than seedlings with a higher shoot–root ratio, due to the increased chance of the roots exploring the soil and acquiring water and soil nutrients for the shoots [
3,
4]. Small seedlings are cheaper to produce and may have a lower initial shoot–root ratio than larger seedlings, but can be more sensitive to competition from other vegetation and to damage compared to larger seedlings [
5,
6]. On fertile sites with relatively high competition from ground vegetation, larger seedlings are at an advantage [
5]. The regeneration success of seedlings of all sizes is, in general, greatly improved by mechanical soil preparation, which is carried out in approximately 85% of regeneration areas prior to planting [
7]. Mechanical soil preparation includes the removal of the organic top layer, which provides a temporary decrease in competition from ground vegetation, as well as exposure and loosening of the mineral soil beneath. The exposed mineral soil warms faster when the organic layer is removed and loosening increases soil permeability [
8,
9,
10]. Increased soil permeability enhances seedling establishment by reducing the physical resistance to root penetration, allowing an additional increase in temperature, which is beneficial for root growth, and increasing nitrogen mineralization [
8,
9,
10,
11]. Furthermore, pine weevils (
Hylobius abietis L.), which are a major threat to planted seedlings in large parts of Sweden [
12,
13], tend to move faster and in straighter lines across exposed areas of flat mineral soil [
14]. Hence, the mineral soil cover in the area surrounding a seedling decreases its susceptibility to damage by pine weevils [
13,
15,
16,
17,
18,
19,
20,
21,
22]. Studies have also revealed that the threat of lethal pine weevil damage is highest before the seedling reaches a stem basal diameter of 10–12 mm [
18,
19].
The most common mechanical soil preparation method is disc-trenching (60%), followed by mounding (30%) and patch scarification (10%) [
23]. Disc-trenching results in continuous furrows where mineral soil is exposed, with an adjacent ridge of the removed soil. The ridges have similar attributes to the capped mounds created during mounding: turning the soil results in a double layer of organic material covered by mineral soil. These methods offer different choices of planting positions, in general with trade-offs between nutrient and water availability. The exposed mineral soil areas adjacent to the capped mounds provide positions that have a continuous connection to soil moisture via capillary water flow, but the mineral soil in Nordic conditions is generally very poor in nutrients. The organic material within the capped mounds provides a source of nutrients for the planted seedling and the elevated position results in an increase in soil temperature, increasing the decomposition of organic material and giving the seedlings a growth advantage over seedlings planted in exposed mineral soil positions [
8,
9,
10,
15,
24] (a graphical representation of the planting positions is shown in
Figure 1b,d). On moist sites, the elevated spots provided by the capped mounds also provide better soil aeration and, thus, a decreased risk of oxygen deficiency in the roots [
9,
11]. However, capped mounds are susceptible to drought because the capillary flow of moisture from below is interrupted by the organic material within [
9]. It is, therefore, important to consider the site conditions carefully before choosing the preferred planting position.
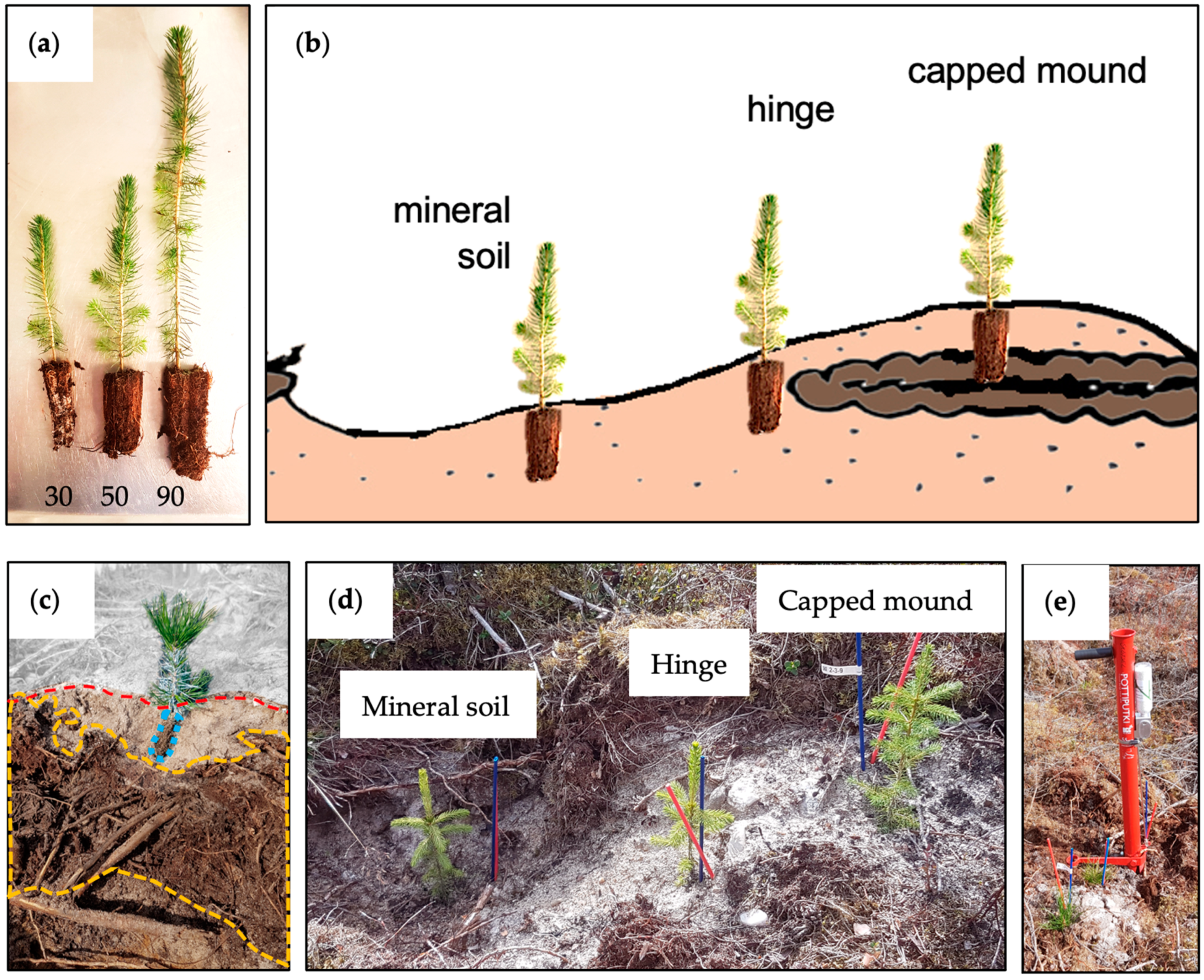
Mounds should ideally be capped by mineral soil, both for the purpose of weighing the organic material down to ensure good contact with the underlying ground and to decrease the risk of attacks by pine weevils. However, there is high variability in the quality of capped mounds depending on site conditions and the way the soil preparation is carried out. Even within a single site, there is a great variability in the occurrence of obstacles such as rocks, stumps, residual logs, ground vegetation density, etc. This heterogeneity prevents any mechanical soil preparation method from creating homogenous planting positions [
10,
25]. Each seedling should ideally be planted deep enough in the capped mound for the root substrate to penetrate the organic layers in order to ensure the availability of capillary water (see the capped-mound illustration in
Figure 1b). However, it is often difficult to plant the seedling deep enough and it is not possible to assess the interior of each capped mound to ensure the optimal seedling root substrate position (see the example in
Figure 1d). With respect to mineral soil positions, the deepest parts of patches/depressions should be avoided since these can become filled with water as a result of precipitation and melting snow. Planting in depressions can thus increase the risk of oxygen deficiency for the seedling roots, especially on flat areas and in fine-textured soil with low permeability. However, on slopes with permeable soil, such as gravel and sand, the risk of waterlogging is minimal. An intermediate choice is to plant the seedlings in the mineral soil close to the capped mound, i.e., the “hinge” of the capped mound, where the seedlings will have ready access to nutrients from the decomposing material in the adjacent mound and, simultaneously, access to capillary water from the mineral soil. An additional option is to plant in positions in the exposed mineral soil further away from the mound and add slow-release fertilizer at the time of planting, to compensate for the low nutrient status, while the position in mineral soil simultaneously ensures water availability from below. An organic slow-release nitrogen fertilizer composed of arginine–phosphate (AP) is available and used commercially for this purpose.
In this case study, we examined the effects of planting position, seedling size, and the addition of AP, as well as the potential interactions between these factors, on the establishment and early growth of Scots pine and Norway spruce seedlings. We hypothesized that (1) survival would be affected by planting position, (2) larger seedlings would attract a higher frequency of pine weevil damage, (3) seedlings of all sizes would grow best in the capped-mound positions, followed by the hinge positions and lastly the mineral soil positions, and (4) the addition of AP would improve growth so that seedlings planted in the mineral soil and hinge positions would achieve similar growth to the seedlings planted in the mound positions.
2. Materials and Methods
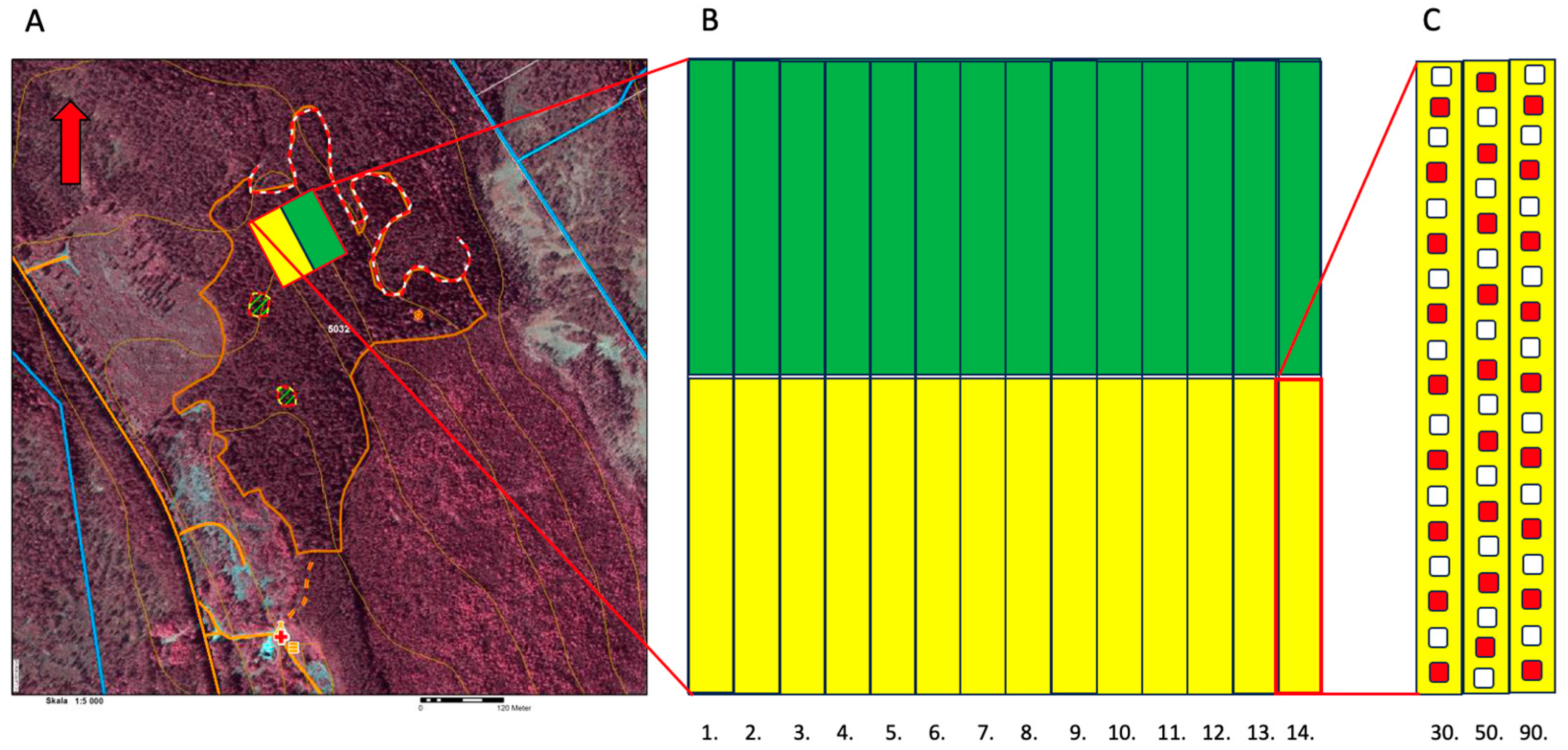
The study site was 100 × 100 m and situated on a clear-cut area with a total size of 12.3 ha in northern Sweden at 63.720954° N, 19.927771° E, ~50 m.a.s.l. (
Figure 2A). The trial area sloped gently towards the east–northeast. The ground vegetation was dominated by bilberry plants (
Vaccinium myrtillus L.) upslope, where the harvested stand was dominated by pine, grading to grass downslope where the harvested stand was dominated by spruce. The soil comprised a thin podzol with underlying mineral soil fractions grading from coarse wave-washed gravel upslope to fine sand downslope, with intermediate mixtures in between. The stand was harvested in June 2018 and mechanical soil preparation by mounding was undertaken in October 2019. The coarser-grained and relatively drier upper part was planted with Scots pine (yellow in
Figure 2) and the lower, more fine-grained, moister part was planted with Norway spruce (green in
Figure 2) during May 2020. The mechanical soil preparation method used was the mounding method. The trial area comprised 14 adjacent plots for each species (
Figure 2B). Each plot comprised three rows with one seedling size in each row (
Figure 2C), the seedlings having been grown in containers with cell sizes of 30, 50, and 90 cm
3; the three respective rows are hereafter referred to by these volumes (
Figure 1a). For each mound, a group of three seedlings was planted: one in the mound, one in the hinge, and one in the mineral soil, i.e., the exposed patch resulting from the removal of the material making up the mound (
Figure 1b,d).
Mounds of insufficient quality, i.e., with insufficient contact with the underlying ground and/or without soil cover, and their accompanying hinge and mineral soil positions were not used in the trial. The variation in numbers of suitable mounds per row resulted in different numbers of seedling groups being planted in each row (7–24 seedlings per row). Every second group of seedlings was given a dose of arginine–phosphate (AP), i.e., arGrow
® Granulat (Arevo AB, Umeå, Sweden), at planting. One dose of arGrow
® Granulat contains 40 mg N and 22 mg P, in the form of L-arginine phosphate (C
6H
17N
4O
6P). All seedlings were marked with numbered plastic sticks to facilitate individual identification, with different colors for each seedling size and with red sticks marking the AP-treated groups (
Figure 1d). All seedlings were planted with commercial Pottiputki (BCC AB, Landskrona, Sweden) planting pipes by experienced workers, deep enough for the substrate to be covered by soil, which was gently compacted by pushing the soil next to the seedling with one foot (i.e., following common commercial practice). For the AP treatment, a Pottiputki with an attached arGrow
® dispenser (designed for this purpose and used in commercial applications) was used (
Figure 1e). One dose was dropped into the pipe when it was inserted into the ground and prior to putting the plant in the pipe, so that the dose ended up directly below the seedlings’ roots. The seedlings were planted during the period 18–29 May 2020.
All seedlings were sown in 2019 and grown in the nursery at Skogforsk, Sävar, Sweden. Seedlings were grown in containers with cell sizes of 30, 50, and 90 cm
3 for both Scots pine and Norway spruce, then stored frozen during the winter, following common practice. Forty seedlings of each species and size were randomly chosen for initial size measurements: the roots and shoots were separated and dried at 70 °C for ~60 h and then weighed separately (
Table 1). For Scots pine, the percentage of seedlings with primary needles, i.e., only one needle per bundle, was recorded in a control inventory carried out immediately after planting (
Table 1).
After planting, a control inventory was compiled with a more detailed quality classification assessment of the planting positions, identifying whether the seedlings were planted surrounded by mineral soil or in the vicinity of organic material. Being in the vicinity of organic material represents an increased risk of pine weevil damage by offering shelter to the weevils and thus can be considered a low-quality position at sites where pine weevils are common. Following examination of the data, we found that seedlings planted in proximity to organic material had suffered from more pine weevil damage, and these were all grouped as low-quality positions regardless of the original intended planting position. This group included seedlings in the mineral soil area planted with peat or decomposing humus in the patch closer than 10 cm to the humus edge, seedlings planted in the hinge adjacent to a mound that was not covered by mineral soil, and seedlings planted in mounds that were not covered by mineral soil. The remaining seedlings were re-defined as planted in high-quality (HQ) mineral soil, HQ hinge positions, and HQ capped mounds. Due to different patterns regarding pine weevil damage for pine and spruce in positions on capped mounds where there was humus mixed into the mineral soil around the seedlings, such positions were grouped as low-quality positions due to the high pine weevil damage for spruce, but were not grouped as such for pine. Since high-quality positions were prioritized during planting, they outnumbered the lower-quality positions for each main position. The grouping of all low-quality positions as a separate group allowed us to obtain a clearer definition of the main planting position characteristics and define the potential effects of planting position with less interference from the effects of pine weevil damage.
Seedling development records were compiled at the end of each of the first three growing seasons following planting: on 24–26 August 2020, 6–10 September 2021, and 2–8 September 2022. The inventories included recorded scores for vitality (all years, 0 = dead or dying, 1 = severely low vitality, 2 = moderate decrease in vitality from damage and/or damaged leading shoot, 3 = none or minor decrease in vitality, undamaged leading shoot); leading shoot length (all years, in cm); height (all years, measured from ground level to top bud in cm); stem basal diameter (in 2022, measured close to ground level in mm); level of pine weevil damage (all years, not including healed damage from previous years; 1 = minor, <25% of the stem circumference damaged; 2 = moderate, 25%–50% of the stem circumference damaged; 3 = severe, >50% stem of the circumference damaged; 4 = seedling killed by pine weevil damage); other damage and, when applicable, the causes of other damage; and mortality.
A subset of seedlings was harvested (starting from plot 1, seedling groups 1 and 2, and then every second pair of AP-treated and untreated seedling groups in every second plot for each species up to plot 11), including the roots, in the period 10–12 October in 2022. In total, 524 pine and 510 spruce seedlings were harvested. For these seedlings, stem base, seedling height, and leading shoot length were measured. The roots and shoots were separated at the base of the stem, then the roots were rinsed to remove soil and other foreign material under running water. The roots and shoots were dried at 70 °C for ~60 h and weighed. Comparisons of their dry weights to the field measurements were used as the basis for choosing which growth parameter to use in the analysis of field data for each tree species.
Scots pine and Norway spruce were analyzed separately. All analyses were carried out using R-studio software version 1.4.1106 [
26]. We applied generalized mixed-effect models using the glmer function in the [lme4] package version 1.1-27.1 [
27] with a binomial distribution for the response variables for survival, mortality, and damage by pine weevils from the field inventory data. Planting position (i.e., HQ mineral soil, HQ hinge, HQ capped-mound, and LQ positions), seedling size, and AP addition were used as fixed factors, and the plot was set as a random factor to account for the varying number of seedlings in each row as well as other potential within-site variability. ANOVA tables from these analyses are presented with the results for survival, damage, and mortality caused by pine weevils. To enable interpretable visualizations of the results, the seedling sizes were analyzed separately when effect of size was significant, following the same procedure as that used in the described analyses but omitting the size factor.
For the measured values, i.e., the diameter of pine and height of spruce from the field measurements and the dry weights and shoot–root ratio for the harvested seedlings, we applied linear mixed models using the lmer function in the [lmerTest] package version 3.1-3 [
28], with AP treatment and planting position used as fixed factors and the plot used as a random factor. For growth, each seedling size was analyzed separately without first analyzing them together, since the effect of seedling size on seedling size would be superfluous.
The analysis of variance (ANOVA) type III function in the [car] package version 3.0-12 [
29] was used to test each model for interactions. Where no significant interactions were found, the model was reduced to include the main effects only and was tested again with ANOVA type II in the [car] package. From all models made for visualization of the results, estimated marginal means were calculated using the [emmeans] package [
30] and presented graphically using the [ggplot2] package version 3.3.6 [
31].
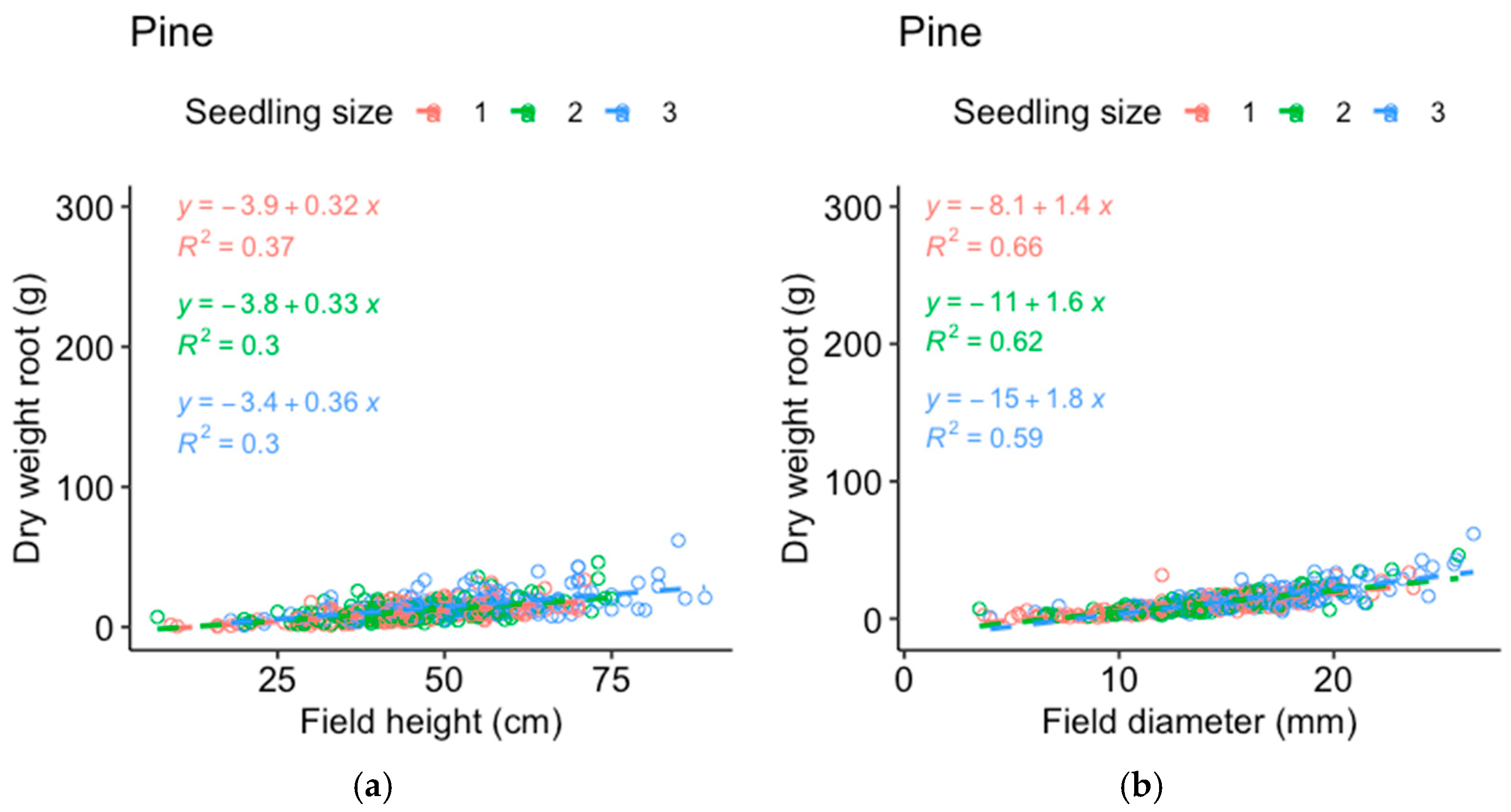
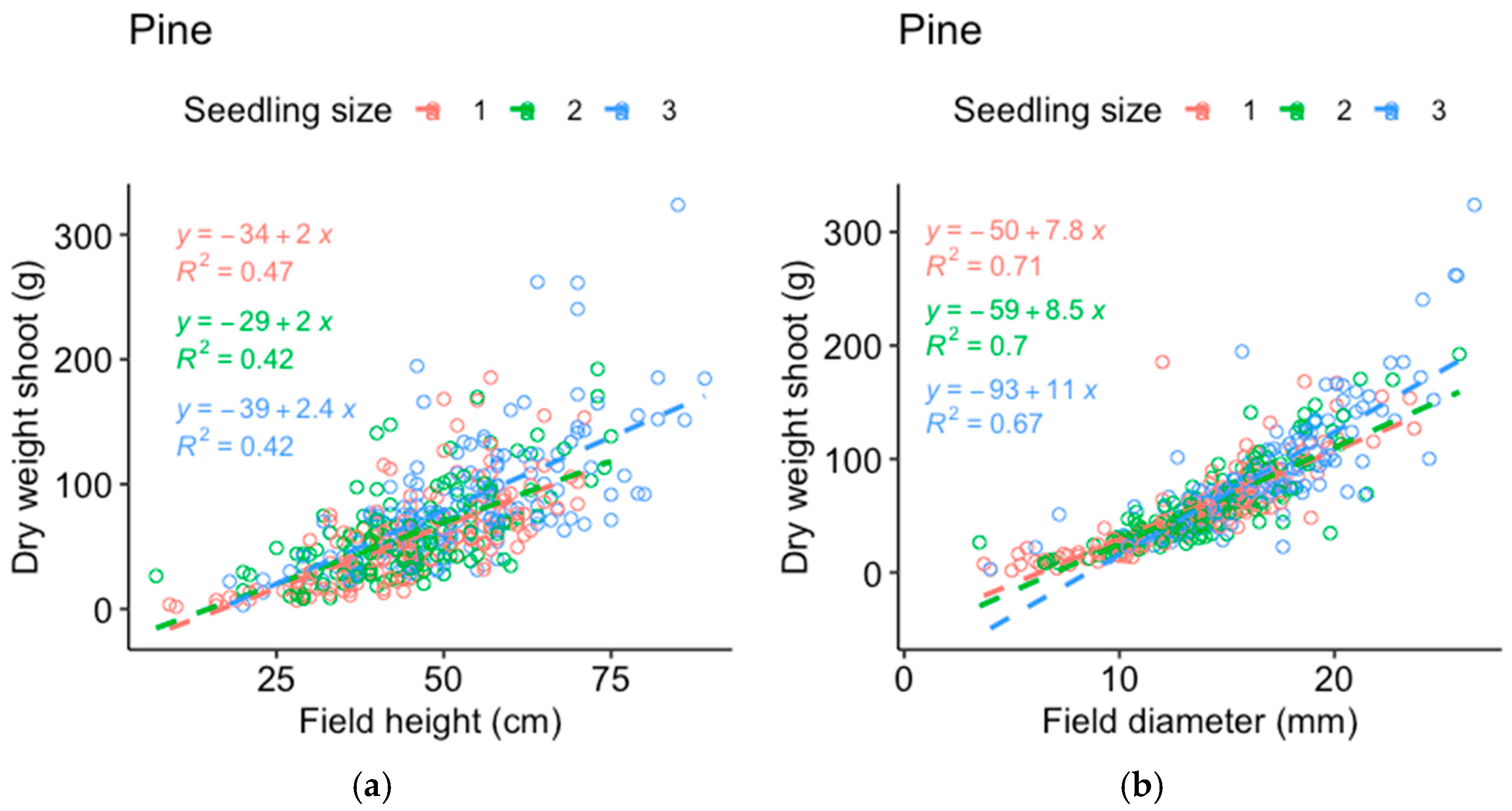
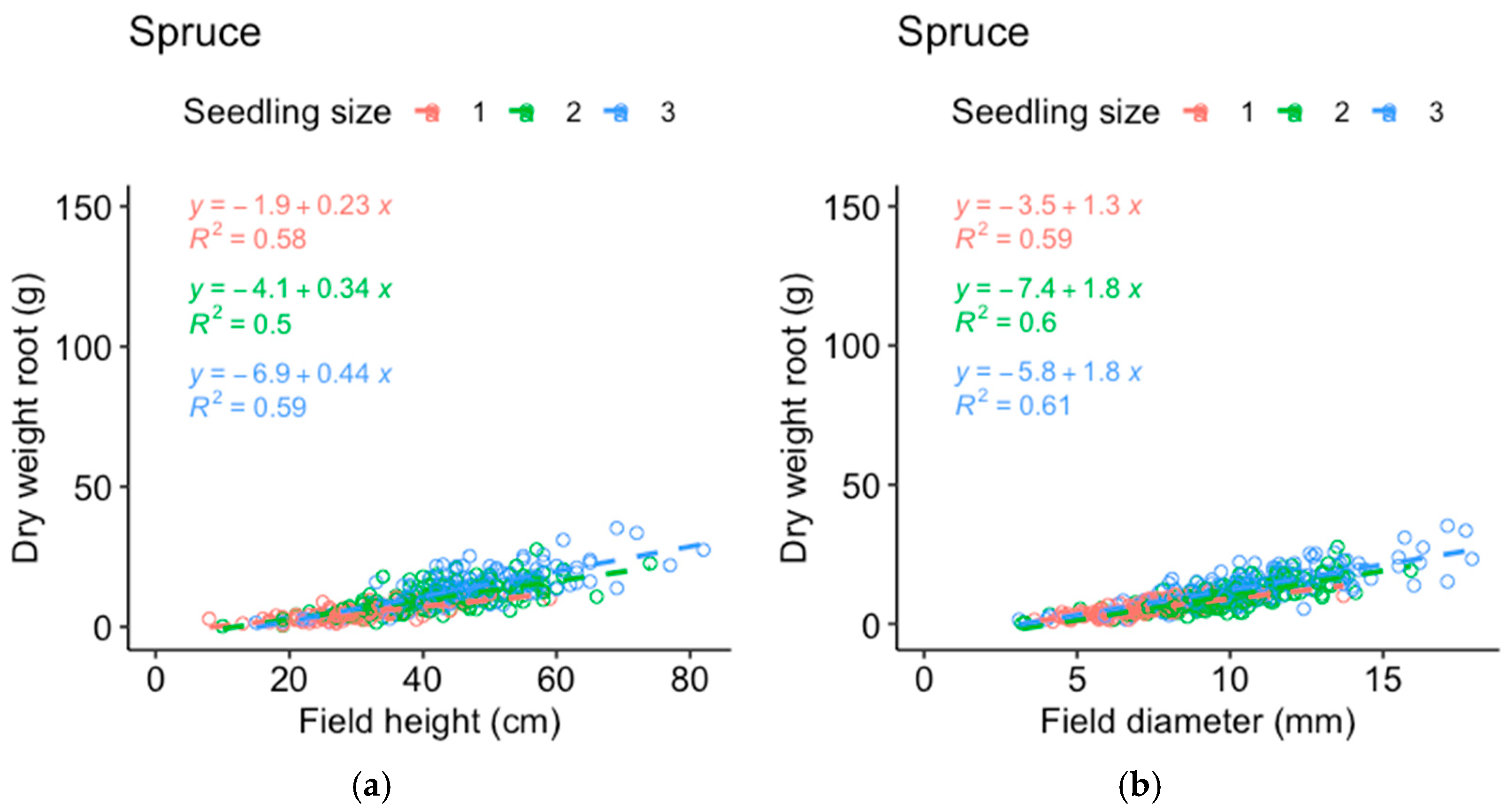
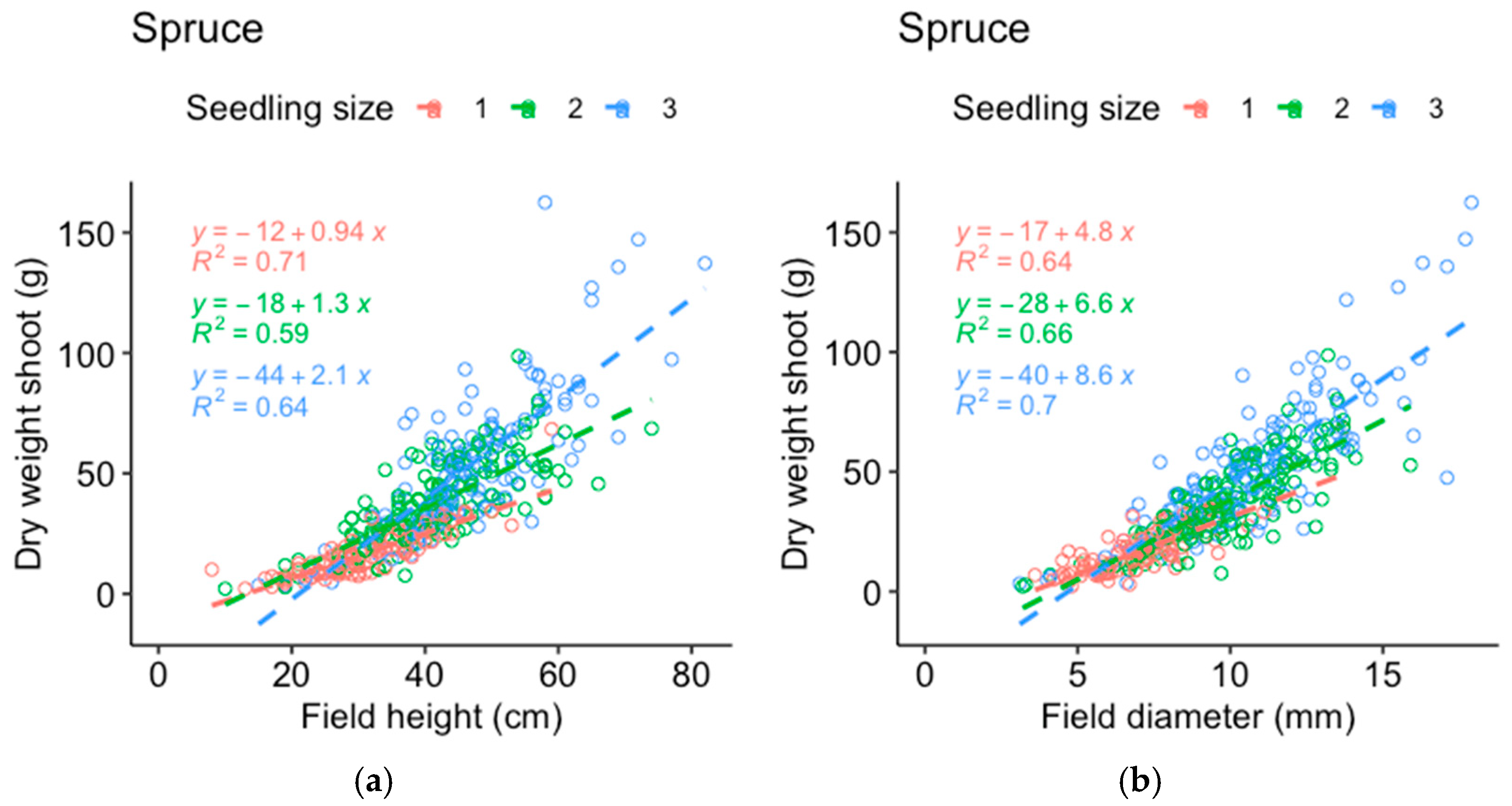
Comparisons between the dry weight data from the harvested seedlings and those from the field measurements of the same seedlings were made to determine which of the measured parameters best represented the seedling mass (
Appendix A). The stem basal diameter correlated best with the dry weight of the pine roots (
Figure A1) and shoots (
Figure A2) and was used as the response variable for pine, while height corresponded almost as well or better compared to the stem basal diameter (depending on seedling size) for the spruce roots (
Figure A3) and shoots (
Figure A4). In addition, field measurements of the spruce stem basal diameter were missing for half of the plots and height was therefore used as the response variable for spruce growth to include as much of the available data as possible. The data on the dry weights of shoots and roots from the harvested seedlings were analyzed following a procedure similar to that followed for the field measurements.
4. Discussion
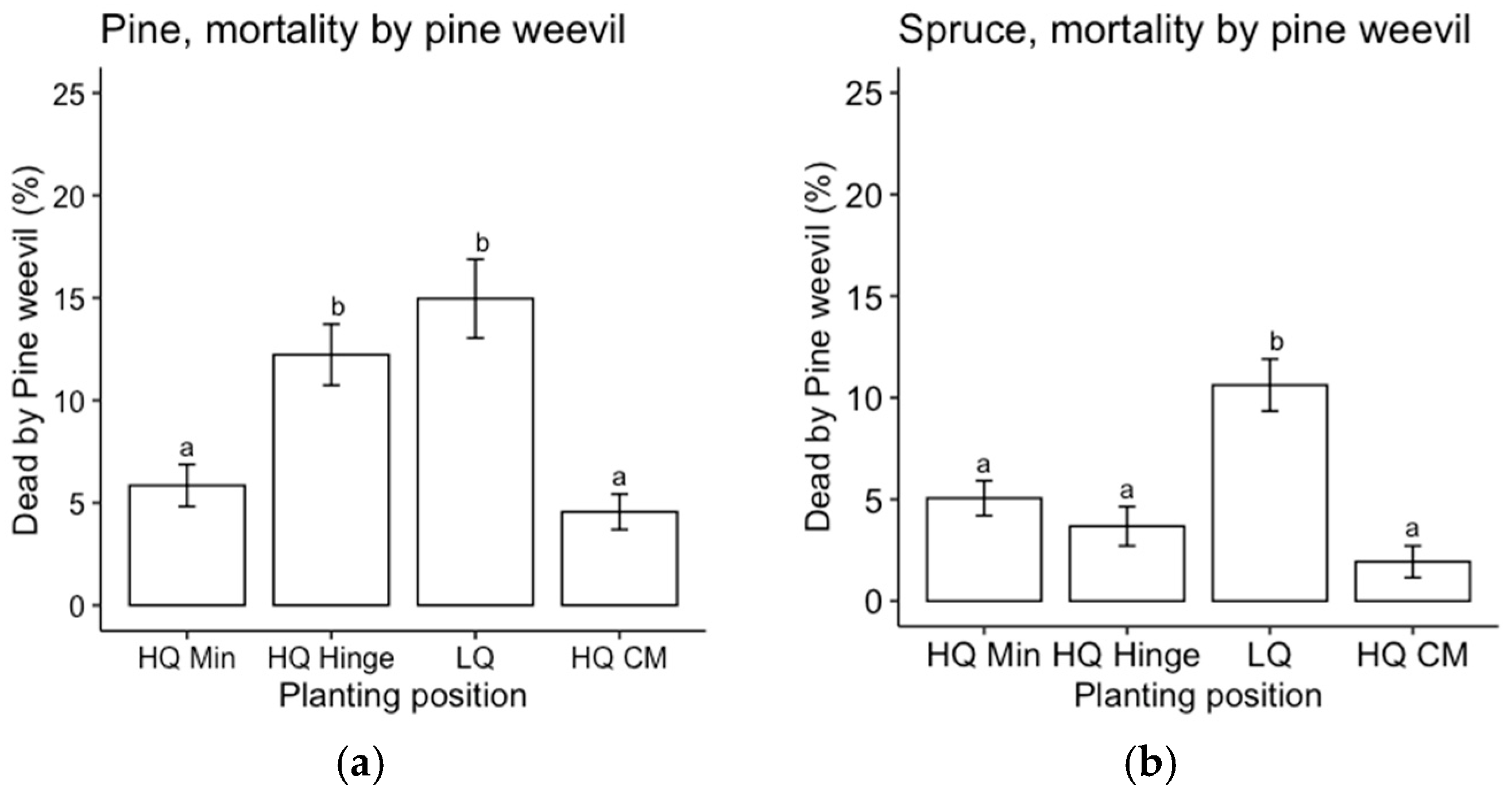
Three years after planting, seedling survival was in the range of 74%–96% for pine and 83%–98% for spruce. The main cause of mortality was pine weevil damage for both pine and spruce. Most deaths caused by pine weevils occurred in 2020 (the year of planting), which is an expected result due to the high risk of thin-stemmed seedlings being girdled [
18,
19,
20].
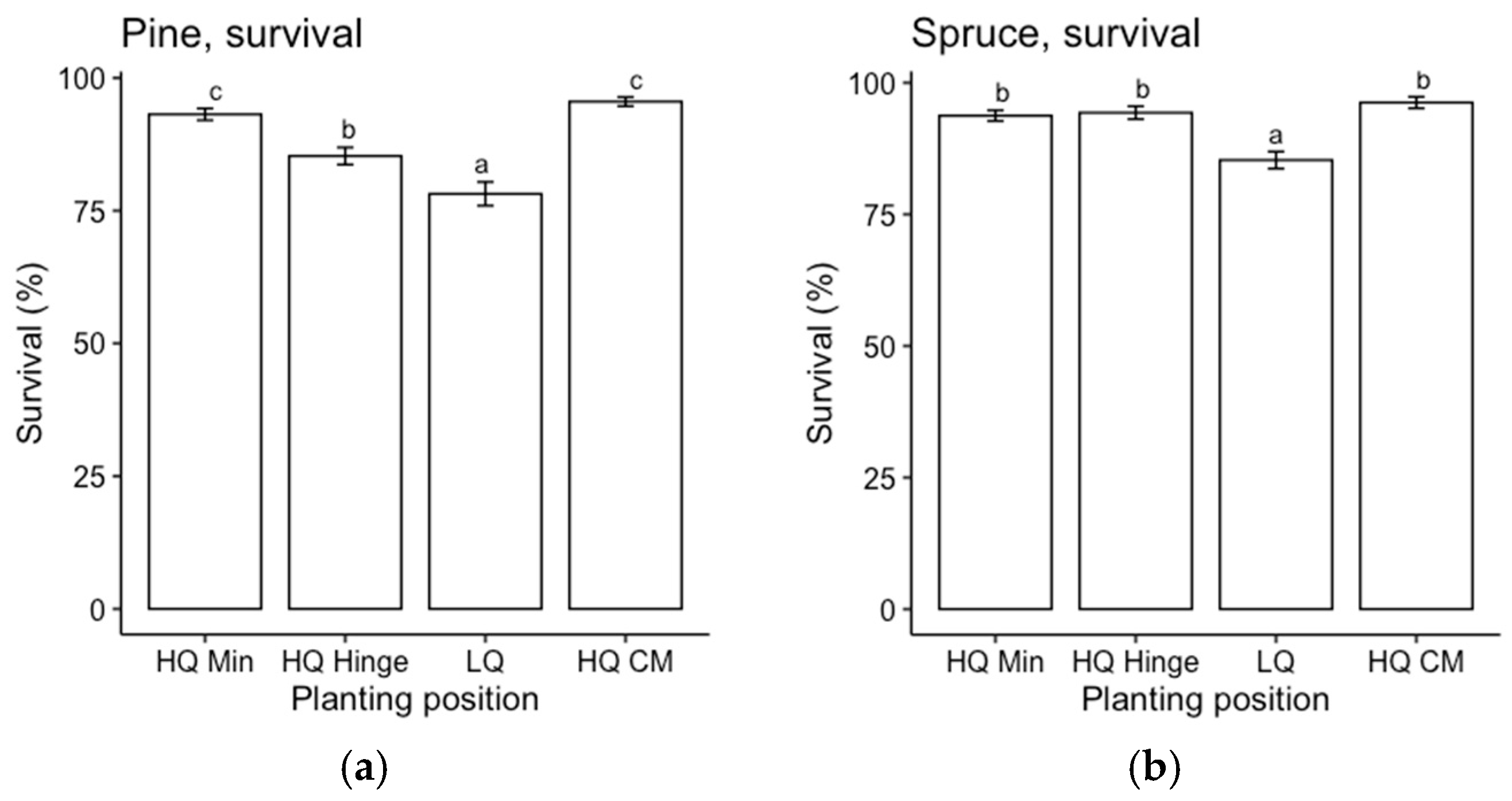
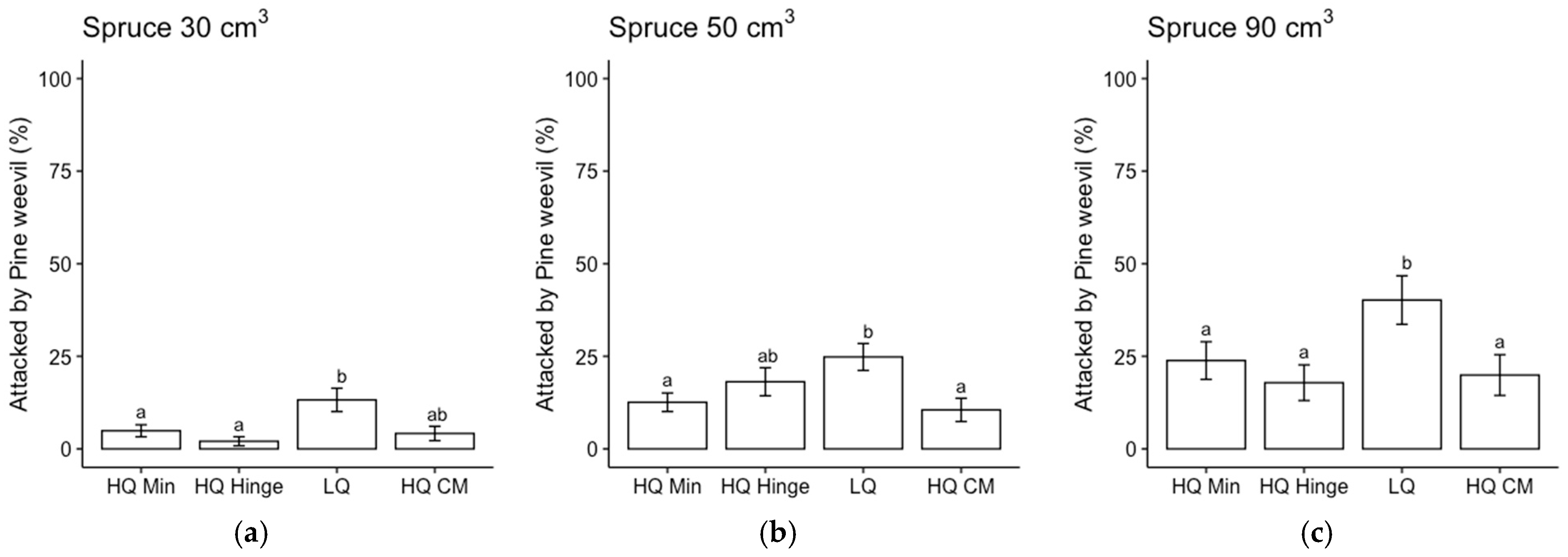
The choice of planting position affected the outcomes regarding survival, in accordance with our first hypothesis. However, it appeared to be related more to the quality of the position rather than the choice of mineral, hinge, or capped-mound position. Mortality was highest in the low-quality planting positions for both pine and spruce, and it was mainly related to pine weevil damage. This was expected since the definition of a low-quality position was based on the occurrence of organic material (i.e., humus and vegetation) in the vicinity of the seedling, which has been found to increase the frequency and level of damage by pine weevils compared to seedlings surrounded by mineral soil [
15,
16,
17,
18,
19,
20,
21]. However, mortality due to pine weevil damage was almost as high in the high-quality hinge positions for pine. Hence, survival in the high-quality hinge positions was lower than in the capped mounds for pine, while there was no significant difference in survival between the high-quality mineral soil and capped-mound positions. For spruce, there was no significant difference in survival between any of the three main positions as long as they were of high quality. This survival pattern was reflected by a higher attack frequency for seedlings planted in low-quality positions.
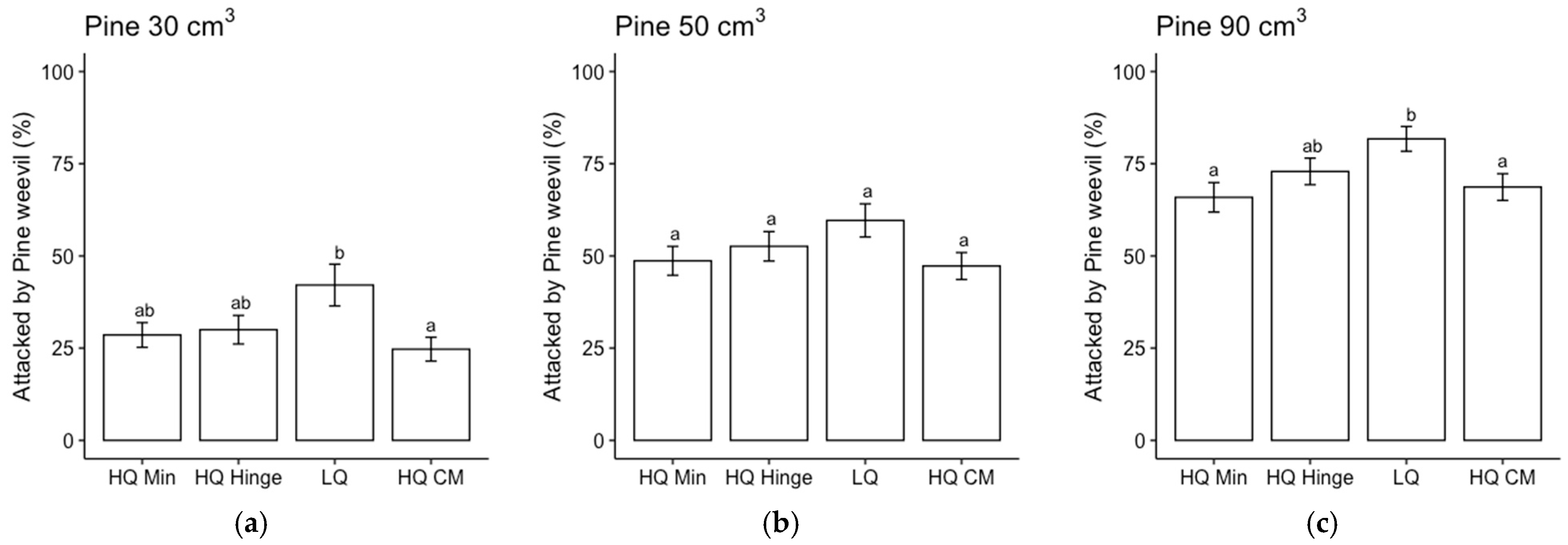
The frequency of pine weevil attacks increased with the seedling size for both species, in accordance with our second hypothesis. Despite a higher attack frequency, the larger seedlings did not exhibit any higher mortality rates compared to the smaller seedlings, indicating that they were able to cope with pine weevil attacks better than the smaller seedlings. This result is in line with previous research, in which larger seedlings attracted more pine weevil damage but exhibited no increase in mortality compared to smaller seedlings [
5]. Pine weevil pressure increases with increasing temperature sum during the growing season and, in general, the harsher northern inland part of Sweden has less problems with pine weevil damage [
5,
32]. However, climate change is increasing the length of growing seasons over the whole country, so pine weevil pressure could potentially increase in more areas.
There was overall a higher attack frequency and higher mortality due to pine weevils for pine than for spruce, which might potentially alter trends associated with the positions in which the seedlings are more vulnerable (e.g., the number of pine weevils competing for the available seedlings may increase their attraction to otherwise less attractive feed sources). The difference in attack frequency between the tree species may, in part, be explained by the larger size of pine, since the 30 cm
3 pine seedlings were approximately equal in dry weight to the 90 cm
3 spruce seedlings. It could also be an effect of species preferences, since pine weevils prefer pine over spruce if both conifers are present [
12,
18]. However, there were also differences in environmental factors between the pine and spruce areas that may have had a direct or indirect effect on the pine weevil attack frequency in different planting positions. The pine area comprised more coarse soil (i.e., fractions larger than sand, including pebbles and stones of various sizes) than the spruce area, there were more mounds containing slash in the pine area, and there was a slightly higher soil moisture level in the spruce area. Soil microtopography has previously been found to affect pine weevils’ feeding behavior [
33], and the occurrence of pine weevils has been found to correlate positively with the amount of slash (harvest residues) [
34]. Hence, the pine area presented a coarser topography, which could potentially explain the higher mortality in the high-quality hinge positions for pine within the subset of plots for which these factors were assessed. Other biotic agents, such as red ants, can also decrease pine weevil damage [
35]. Consequently, factors that affect red ants can have an extended effect on pine weevil damage. The spruce area had, overall, more plots closer to the adjacent forest than the pine area, which may have effects both on the pine weevils’ feeding behavior and red ant presence. The presence of ants was not recorded in our inventories but could be of interest in further studies.
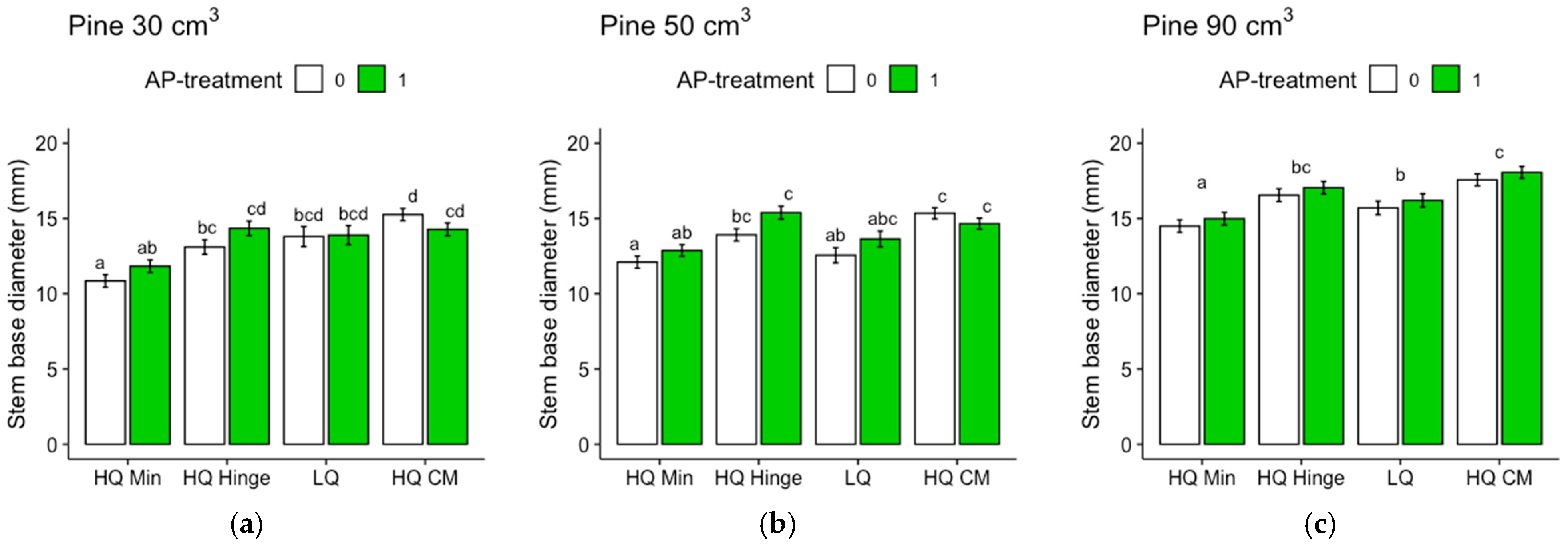
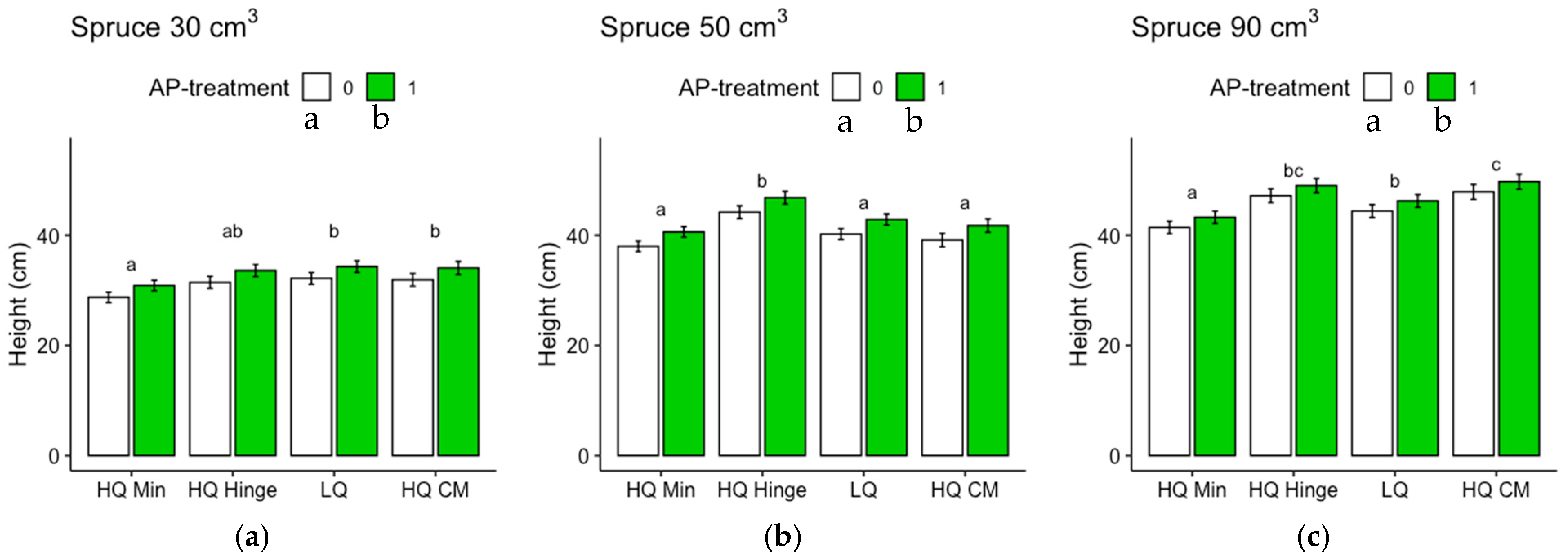
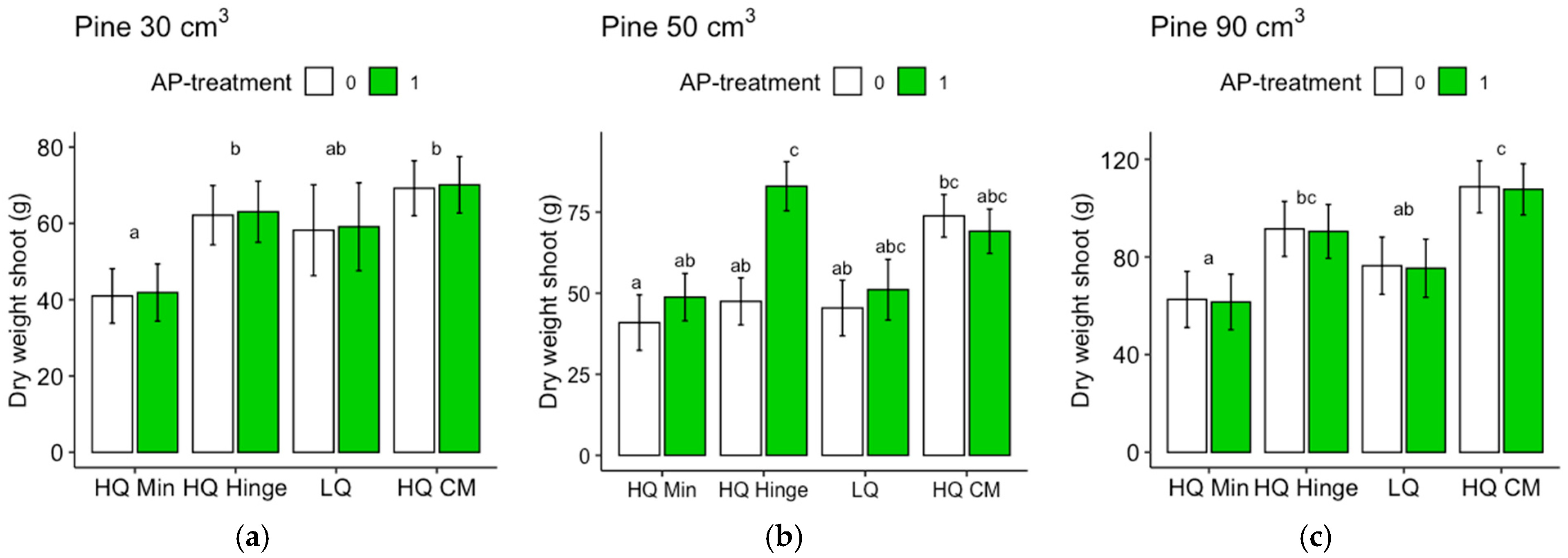
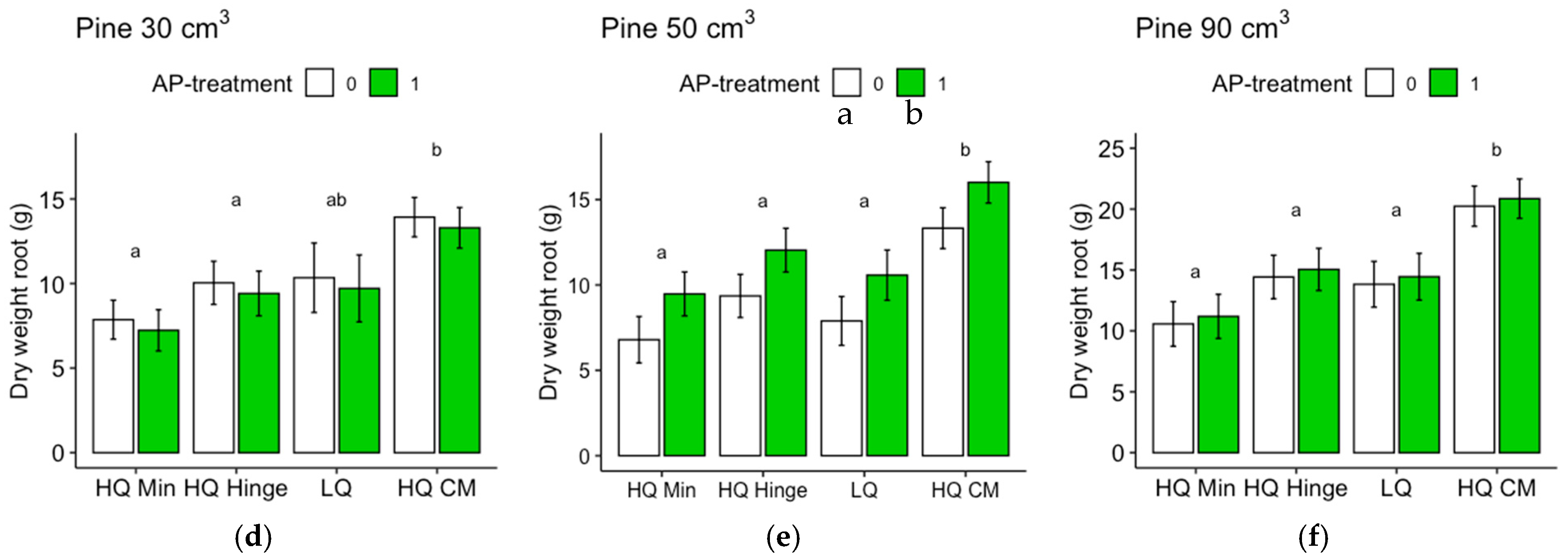
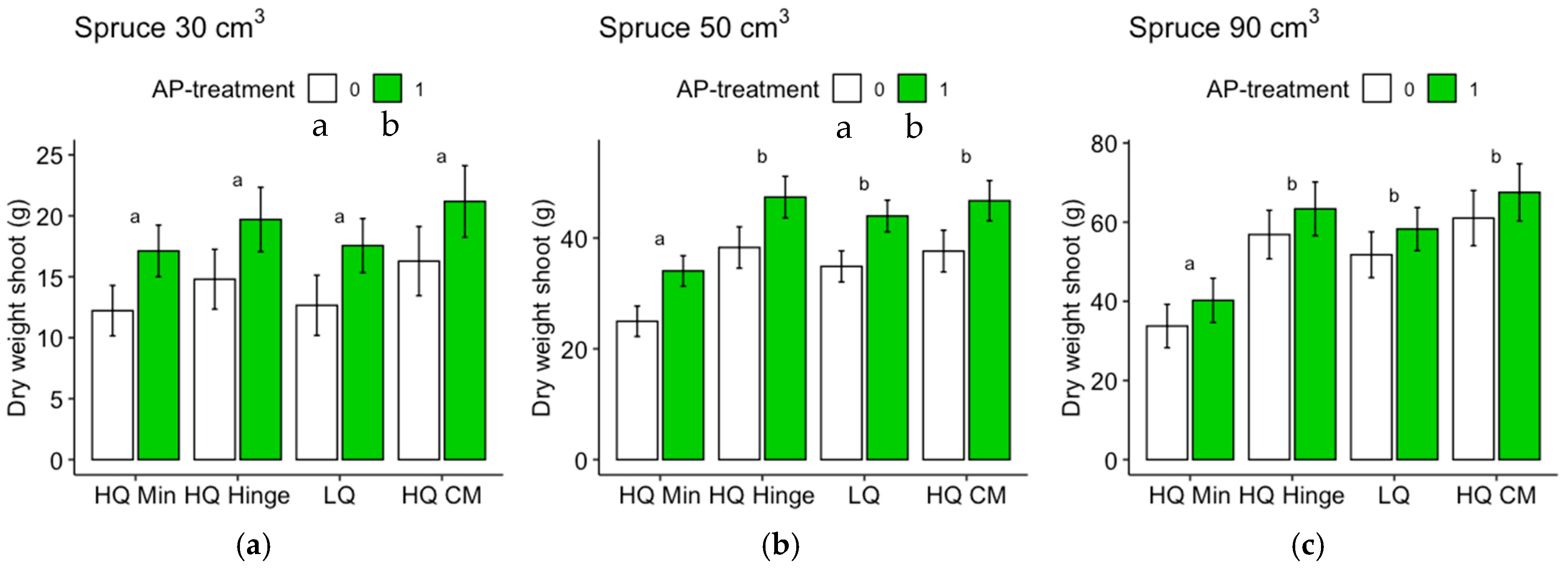
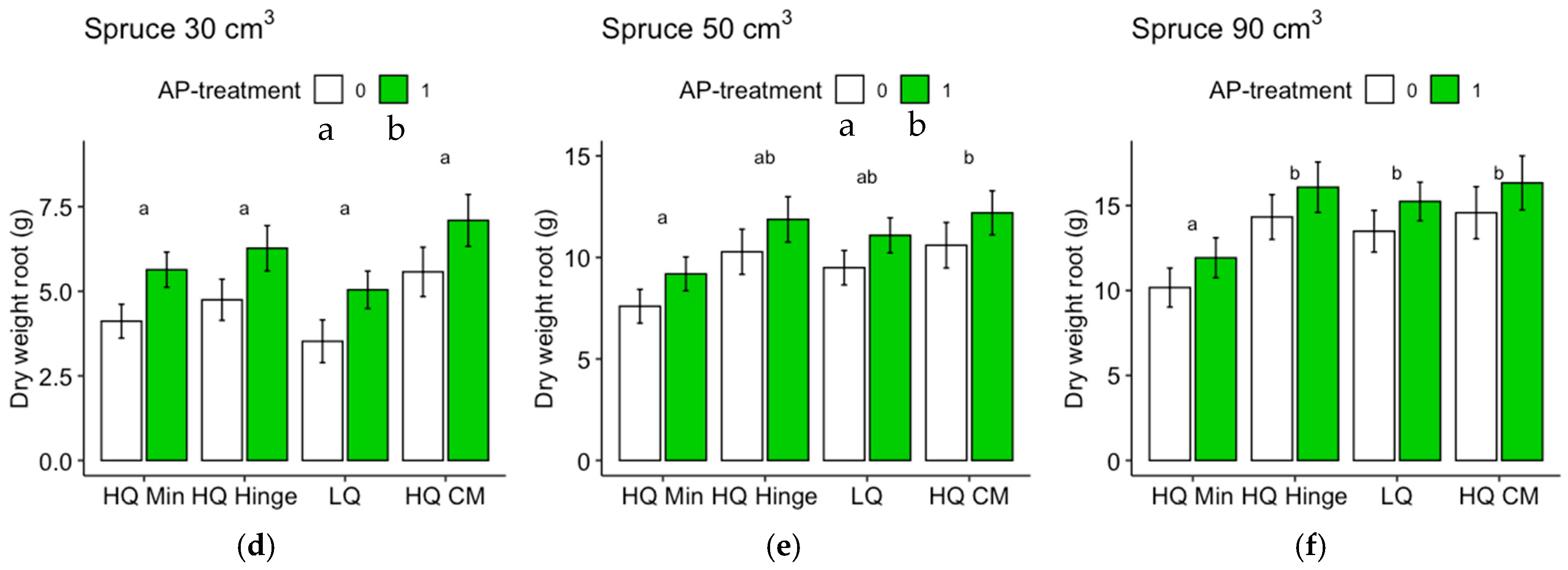
Seedlings, in general, grew less well in the mineral soil positions than in the capped-mound positions, except for the 50 cm3 spruce seedlings, in part corroborating our third hypothesis. However, growth in the high-quality hinge positions was similar to growth in the capped mounds, except for the untreated 30 cm3 pine seedlings. Growth in the low-quality positions was, in general, lower than in the mound positions, but only significantly for the 90 cm3 seedlings of both tree species and the untreated 50 cm3 pine. The effect of planting position on growth appeared greater for pine (up to 40% greater stem basal diameters for the smallest seedling size) than for spruce (up to 16% taller seedlings).
The increase in growth resulting from the AP treatment was marginal and depended on planting position for the 30 and 50 cm3 pine but was not significant for the 90 cm3 pine. However, for spruce, there was a small but significant effect on growth, irrespective of position or seedling size. The addition of AP improved growth so that the seedlings planted in mineral soil achieved similar growth to those planted in the hinge positions for the 30 and 50 cm3 pine and 30 cm3 spruce seedlings, but their growth was not equivalent to that of seedlings in the capped-mound positions, contrary to our fourth hypothesis. The small effect of AP treatment on growth did not fully compensate for the growth loss observed in the mineral soil compared to the seedlings in capped mounds, perhaps representing an insufficient increase in growth compared to the cost and extra effort required in the planting process. However, our results are only short-term, and this study does not show how the effects develop over subsequent years. Considering the potentially lower cost and ecological impact of mechanical soil preparation when patch scarification is used instead of mounding on dry sites, AP treatment could also be seen as a tool to increase growth in mineral soil positions to compensate for potential growth loss compared to capped mounds. Whether this would be economically beneficial or not is a topic for further studies.
Regardless of AP treatment, the option to plant in mineral soil in dry conditions could guarantee more water to seedlings. This would be true for the hinge positions as well, where the seedlings, in general, also grew better than in the mineral soil and equally well as those in the capped mounds (with the exception of the 30 cm3 pine). The relatively high risk of pine weevil damage in the hinge positions for pine found in this study may be different with other site conditions, but should also be considered if the site is within a pine-weevil-dense area. However, if the chosen method was a mechanical soil preparation method that does not aim to make capped mounds, hinges would not be present either.
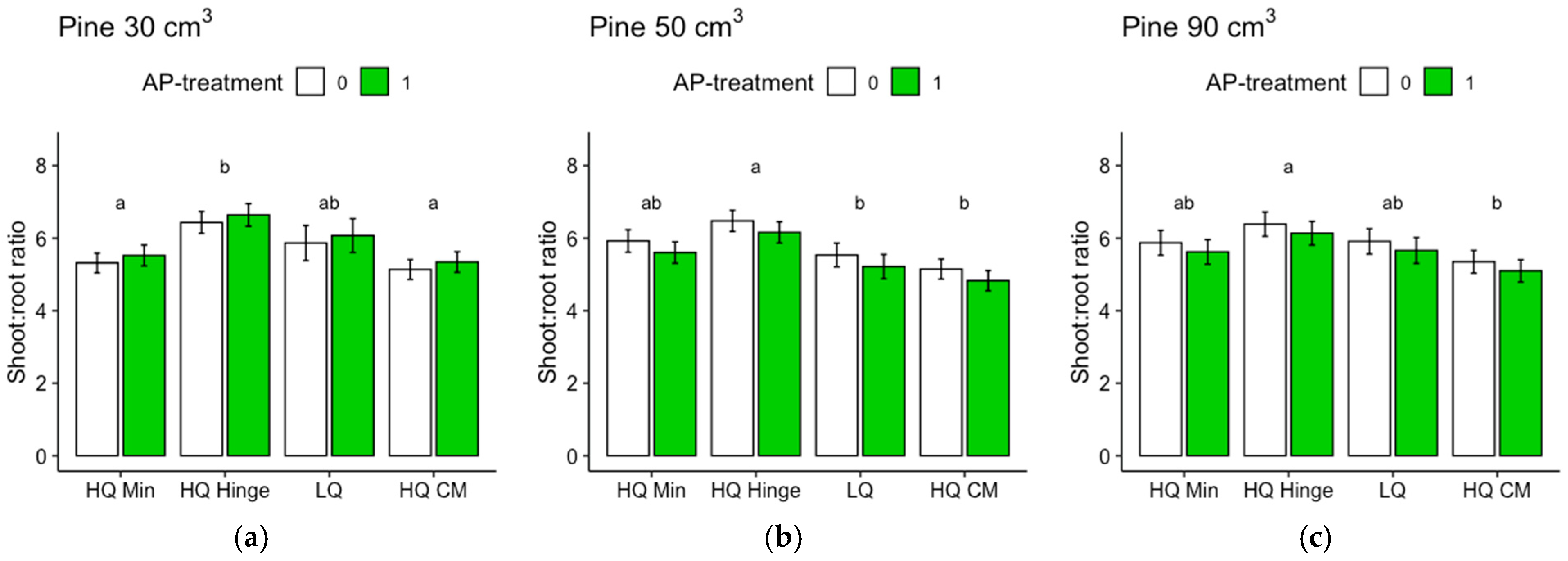
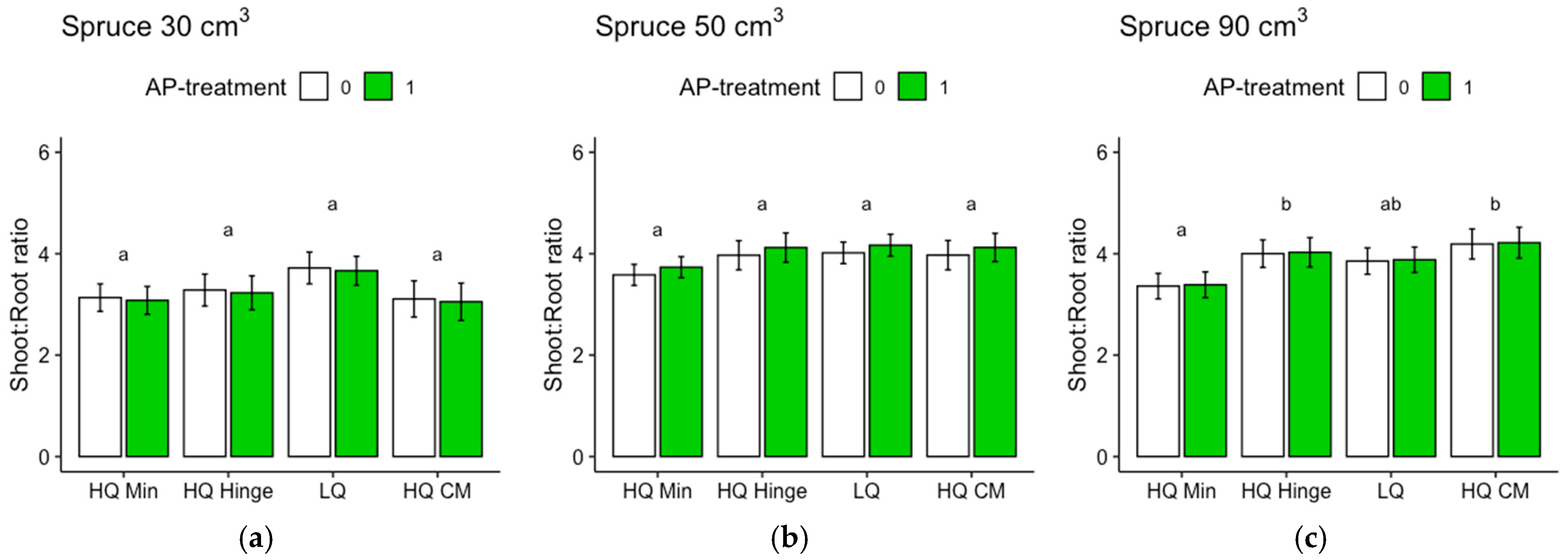
Regarding the analyses of the shoot–root ratios based on the dry weights, it should be noted that the root weights of the harvested seedlings were underestimated, since roots that grew far away from the seedlings were too difficult to dig up. Therefore, the difference between pine and spruce could, in part, be due to different root morphologies. Nevertheless, the analyses give an indication of the growth distribution above and below ground, which is seldom assessed in field studies. The harvested spruce seedlings had lower shoot–root ratios than pine, suggesting that the spruce grew relatively more below ground compared to pine, as shown by the shoot–root ratio of pine increasing by approximately three times the original ratio, while for spruce, the ratio only almost doubled. The shoot–root ratio of pine was highest in the hinge positions for all three sizes. This may potentially indicate that in this position, there is a balance between the water and nutrient availability that allows the seedlings to spend comparatively more resources on shoot growth than on root growth.
The capped-mound position has, in Swedish forest regeneration, long been considered the optimal planting position. Based on the survival and growth results in this study and other studies in which seedlings performed well in capped mounds, it may seem like this is indeed the optimal planting position. It should, in this context, also be noted that the total precipitation during the first month following planting was ~39 mm at the closest SMHI weather station, Torrböle D, ~20 km from the study site (SMHI 2023), which is sufficient for water supply from above. If, instead, there had been a dry month, mortality would probably have been higher for the capped-mound positions, since seedlings planted in mounds are more dependent on precipitation [
9,
36].
Mounding is a method that is most appropriate for moist and/or flat sites with fine soil where the seedlings have the advantage of elevation, especially to improve soil aeration. At such sites, lower-elevation planting positions would be at risk of oxygen deficiency in their roots [
8,
9]. It is also important to remember that the ideal positioning of the seedling root substrate, as presented in
Figure 1b, is rarely achieved. In practice, capped mounds are highly heterogenous due to the high variation in site conditions. In practice, it is often difficult to plant the seedling deep enough due to stoniness or other obstacles. Furthermore, it not possible to assess the interior of each capped mound to ensure the seedling root substrate position. On dry to mesic sites with permeable soil, patch scarification may be sufficient, considering that the effort expended in making capped mounds does not always result in sufficient suitable planting spots in the actual mounds. In addition, the risk of capped mounds being too dry when there is limited precipitation during the establishment phase would be avoided.
Furthermore, instructing planters not to prioritize capped mounds on dry sites would logically decrease the risk of planting in low-quality positions, considering our results in which there was a lower risk of choosing low-quality positions in mineral soil than in hinge and capped-mound positions. The proportion of low-quality positions based on the total percentage for each main planting position in our study was approximately 15% for the mineral soil positions, 28% for the hinge positions, and 31% for the capped-mound positions (averaged over the classifications for pine and spruce).
As a final remark, the results in our study, as well as other trial case studies, are likely biased towards a better outcome than in a commercial regeneration situation, where the variability of planting position quality might be higher. Within-site variation, including the number of suitable planting spots and water availability, affects the survival of seedlings [
37]. We actively avoided planting positions where the mounds were of insufficient quality (e.g., not compacted due to the interference of branches, logs, or other obstacles). The maximum potential number of planting positions for each tree species in this study would, in theory, be approximately 1000, based on the maximum number of seedlings in a row. In reality, 633 triplets of pine and 643 of spruce were planted, i.e., only 60% in the pine area and 64% in the spruce area of potential planting positions were used in this trial. The remaining potential planting positions were either of poor quality (e.g., not compacted due to the interference of branches, logs, or other obstacles) or absent (e.g., where the mechanical soil preparation failed due to obstacles).
Where a seedling ends up being planted depends on the planting performance as well as the availability of high-quality planting positions. Planters are generally paid per planted seedling, and to consider the planting positions as carefully as in this trial would be too time-consuming for them. The availability of high-quality positions also depends on the MSP method and performance, which, in turn, is highly dependent on local site conditions.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











