
Influence of Soil Quality on the Browsing Intensity of Ungulate Herbivores on Tree Species in European Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Study Design and Plot-Level-Data
2.3. Data Generation and Soil Lab Analysis
2.4. Statistical Analysis
3. Results
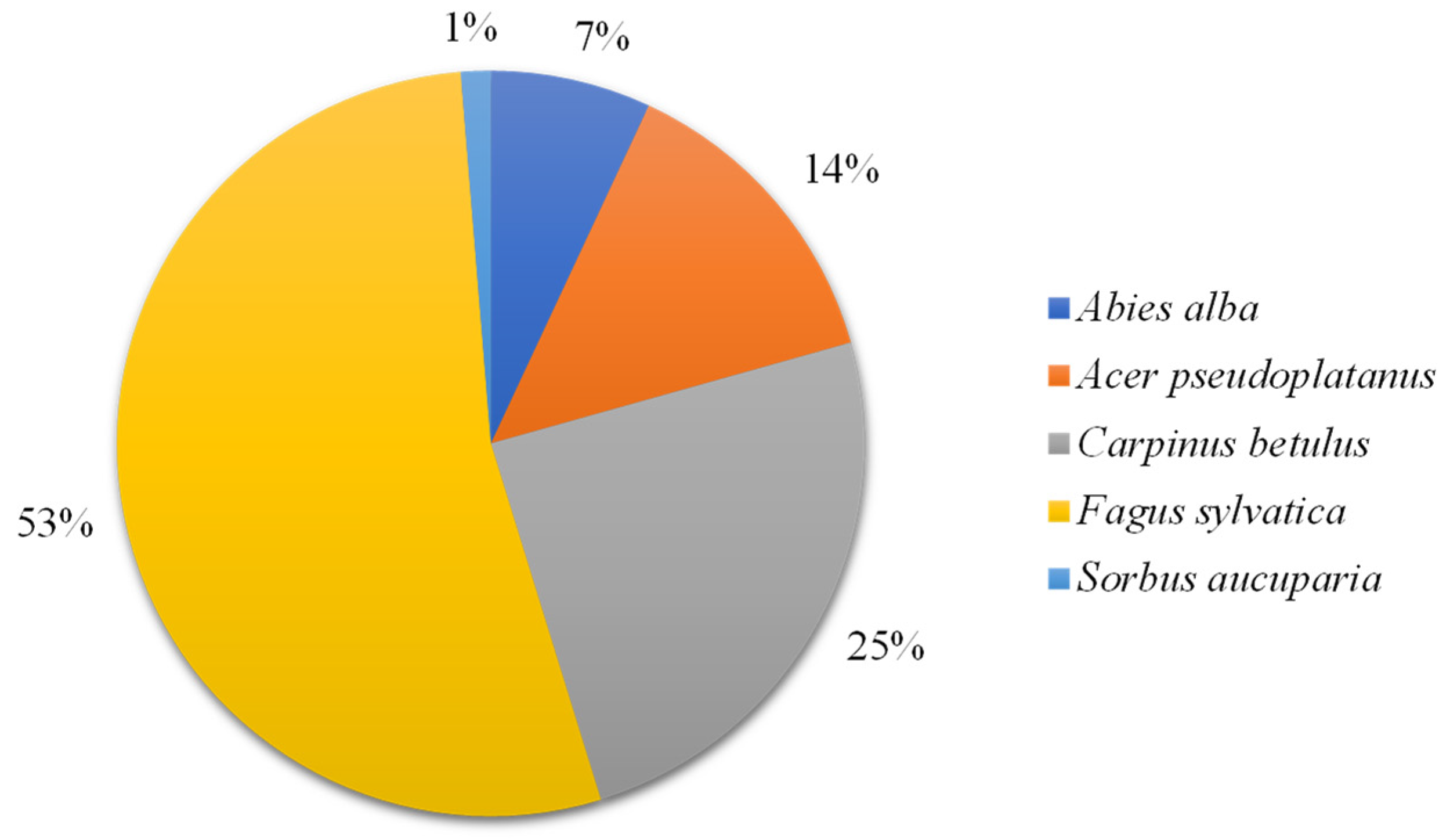
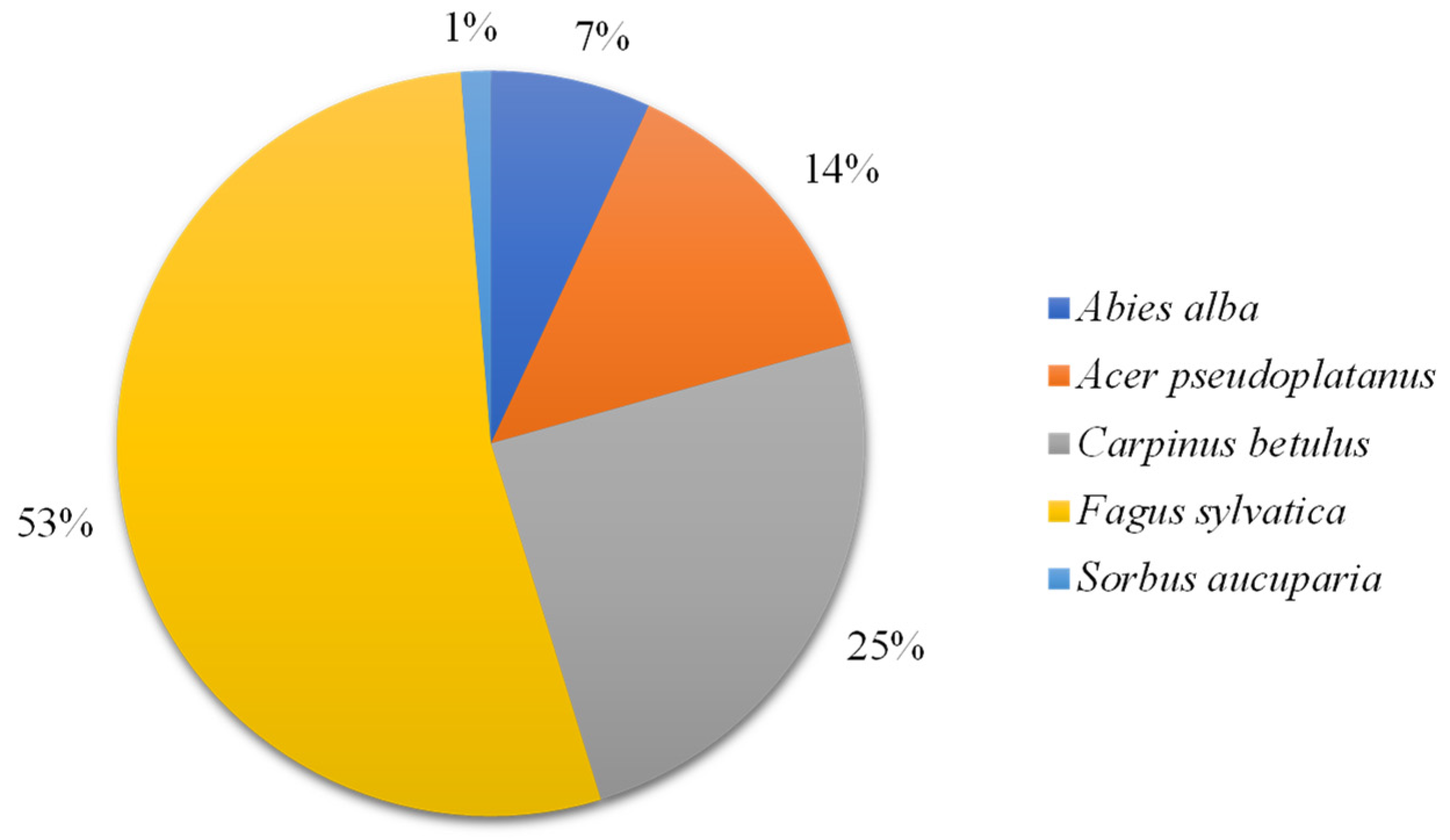
3.1. The Relative Proportion of Tree Species and Descriptive Summary of the Soil Properties
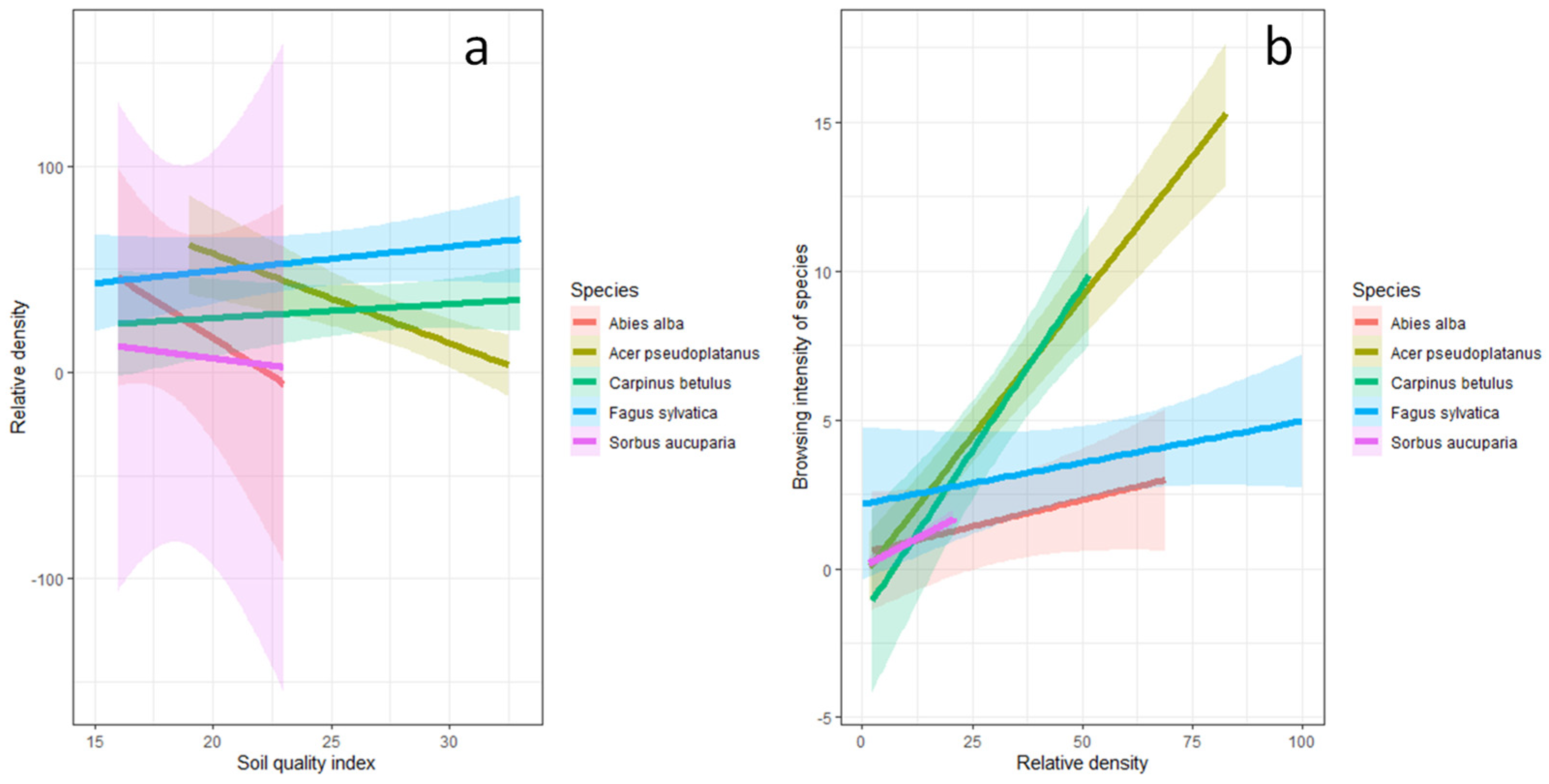
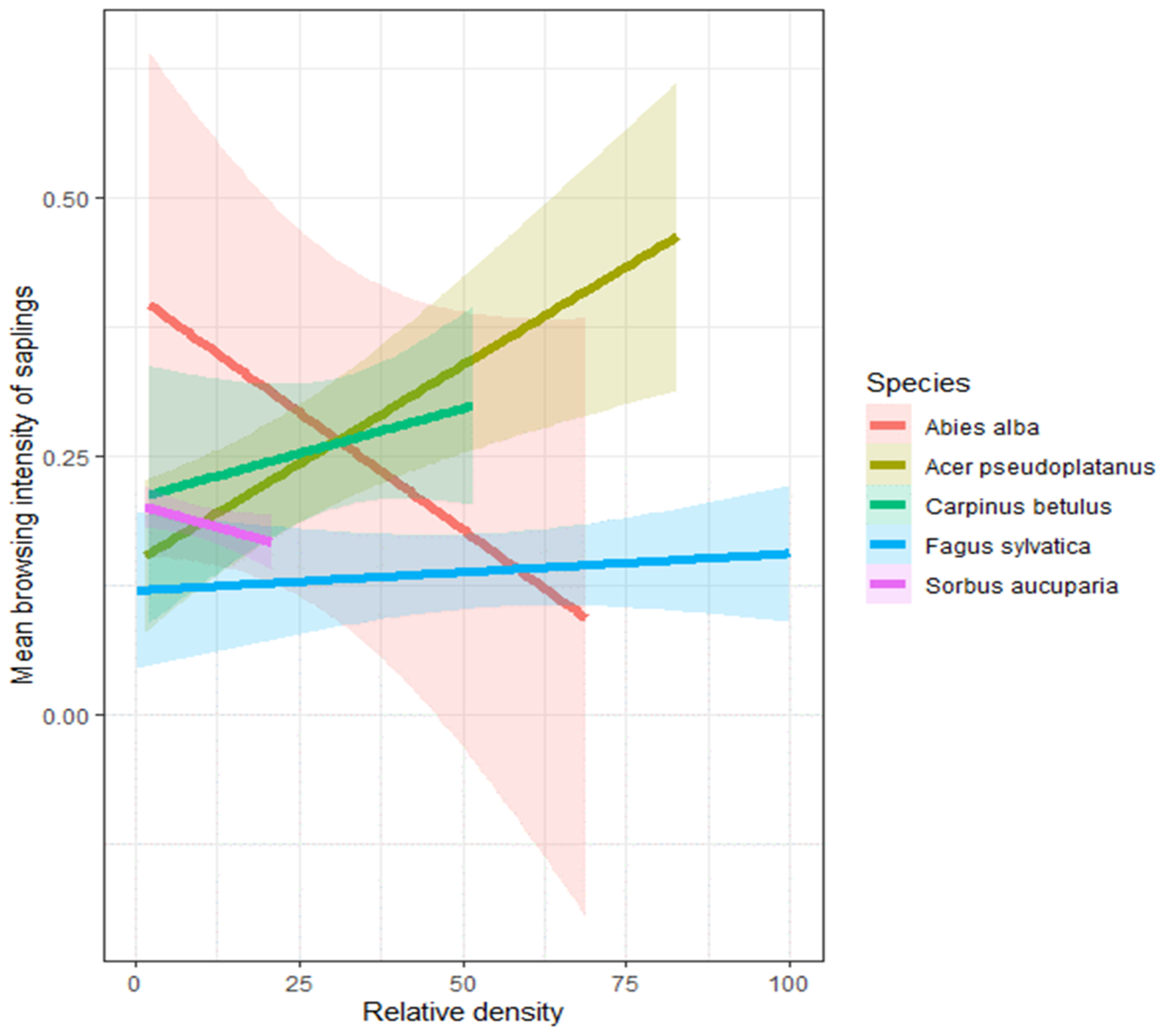
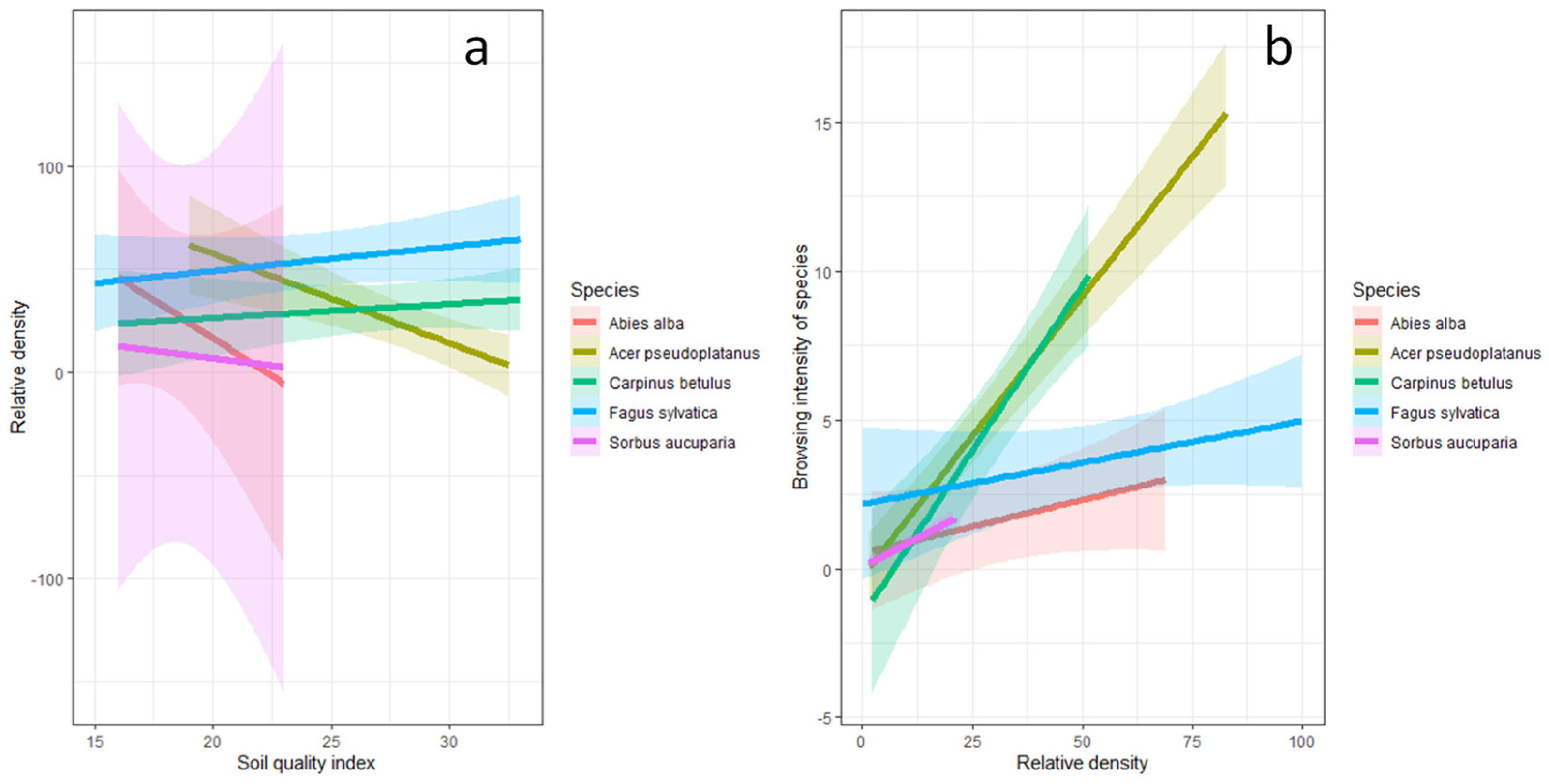
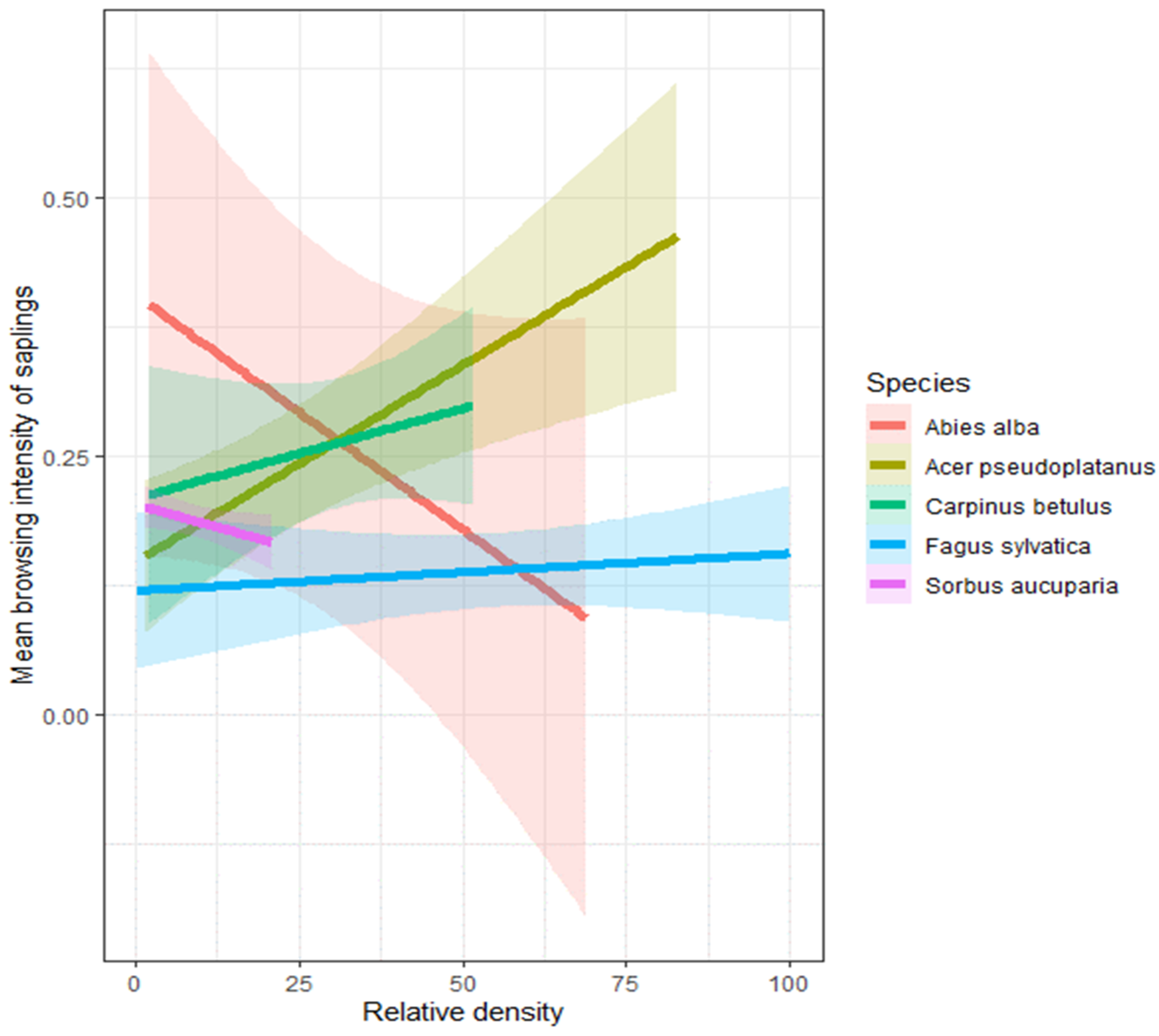
3.2. Relationship between Soil Quality Index, Tree Species Relative Density, and Browsing Intensity
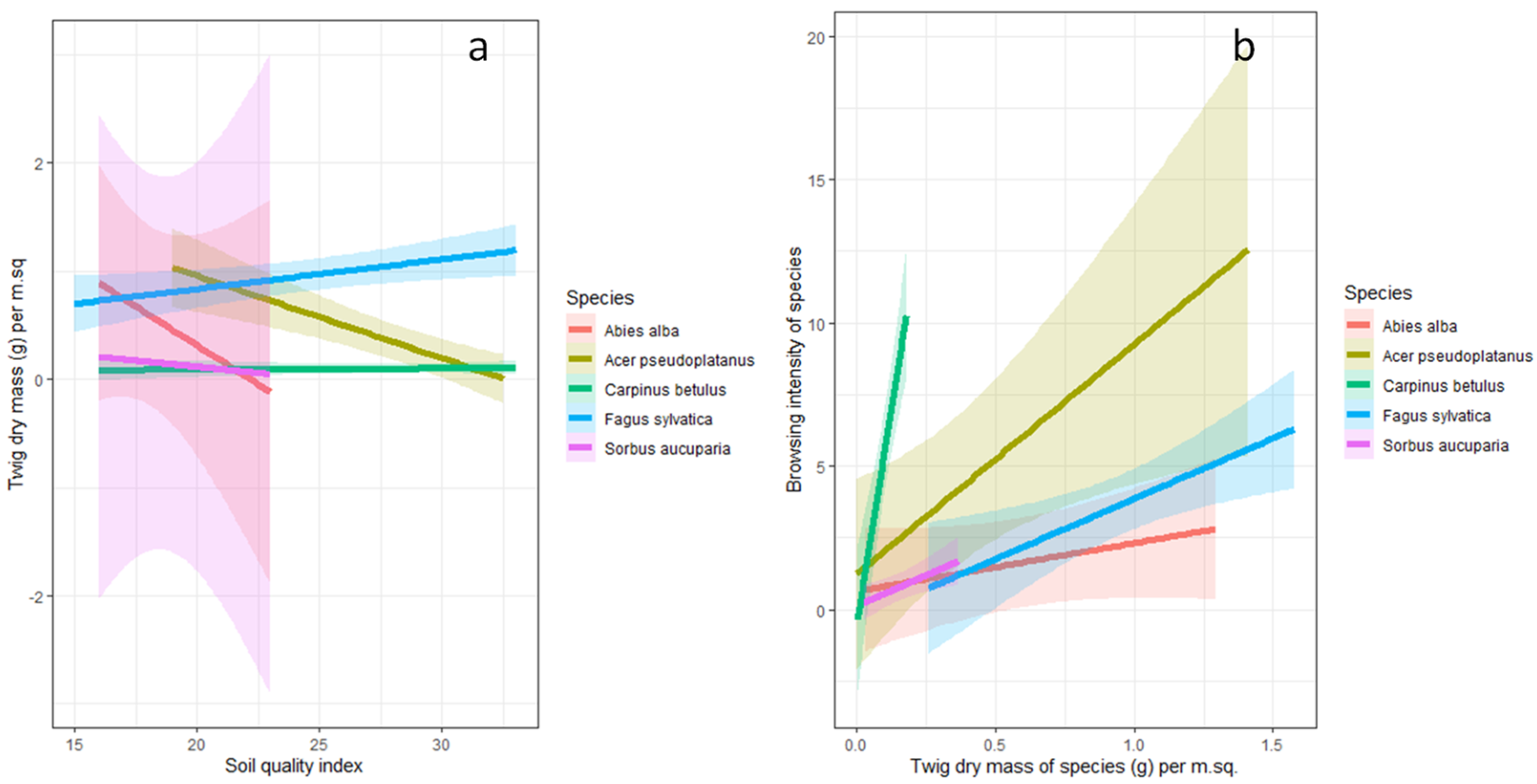
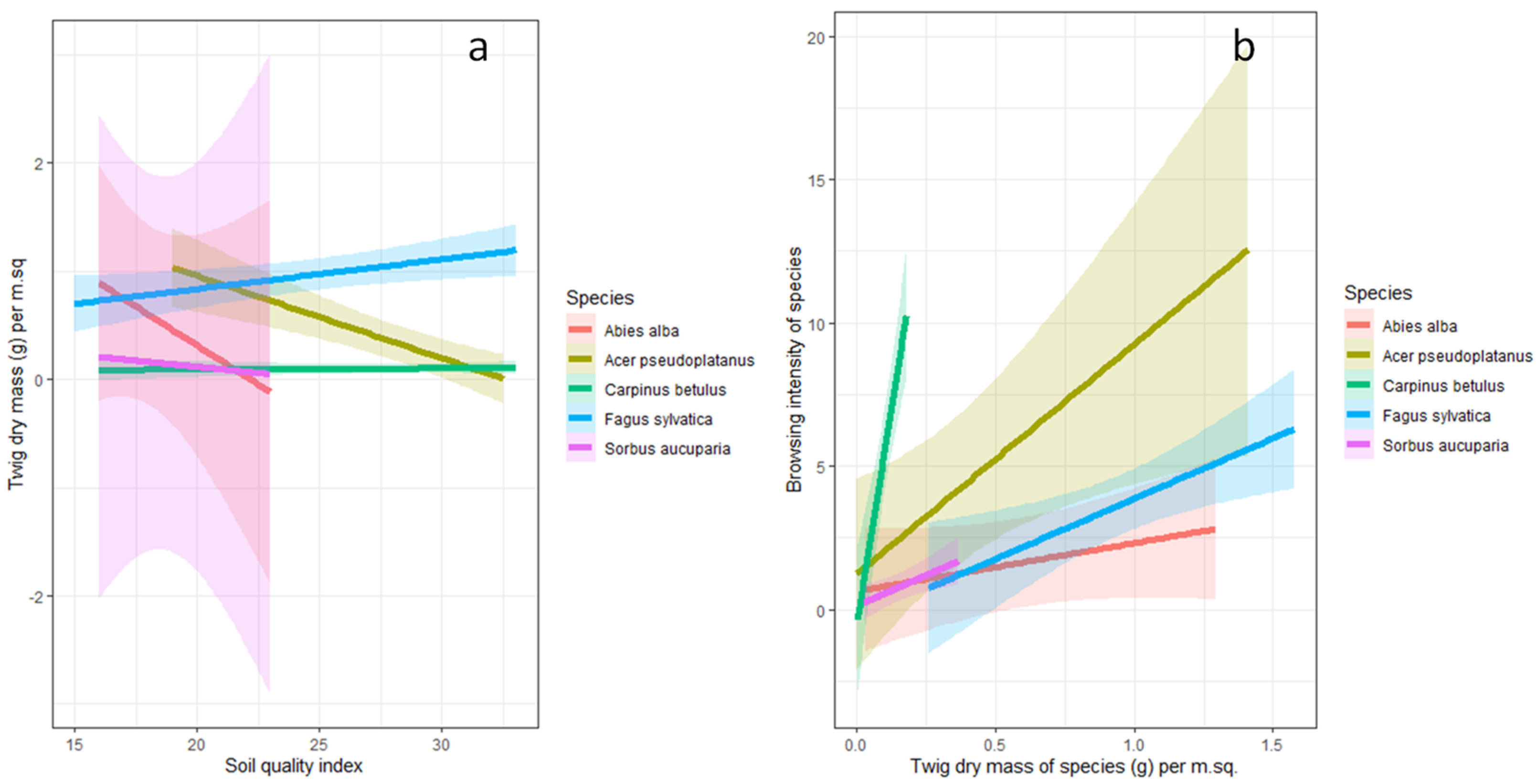
3.3. The Relationship between Soil Quality Index, Twig Dry Mass and Browsing Intensity
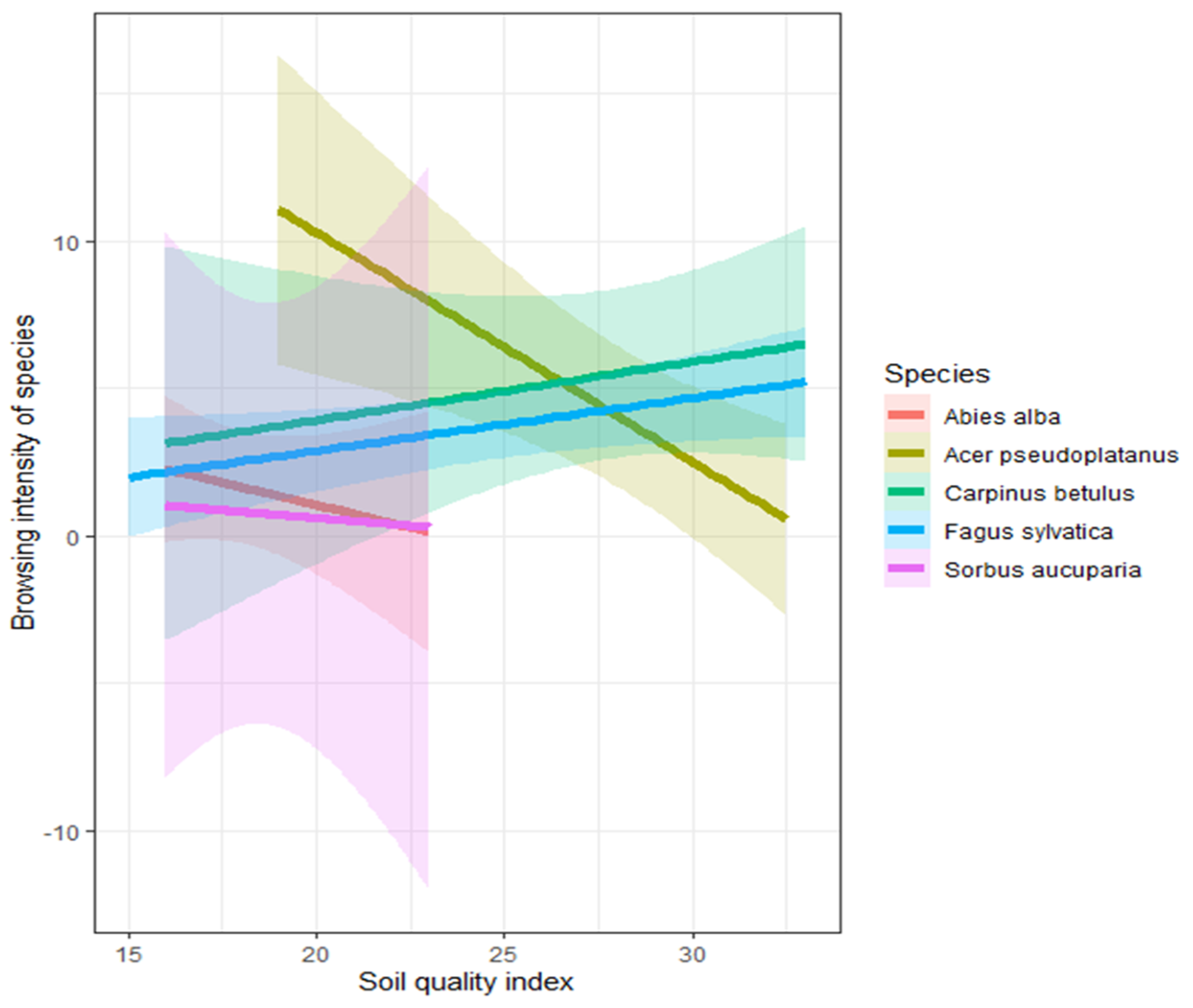
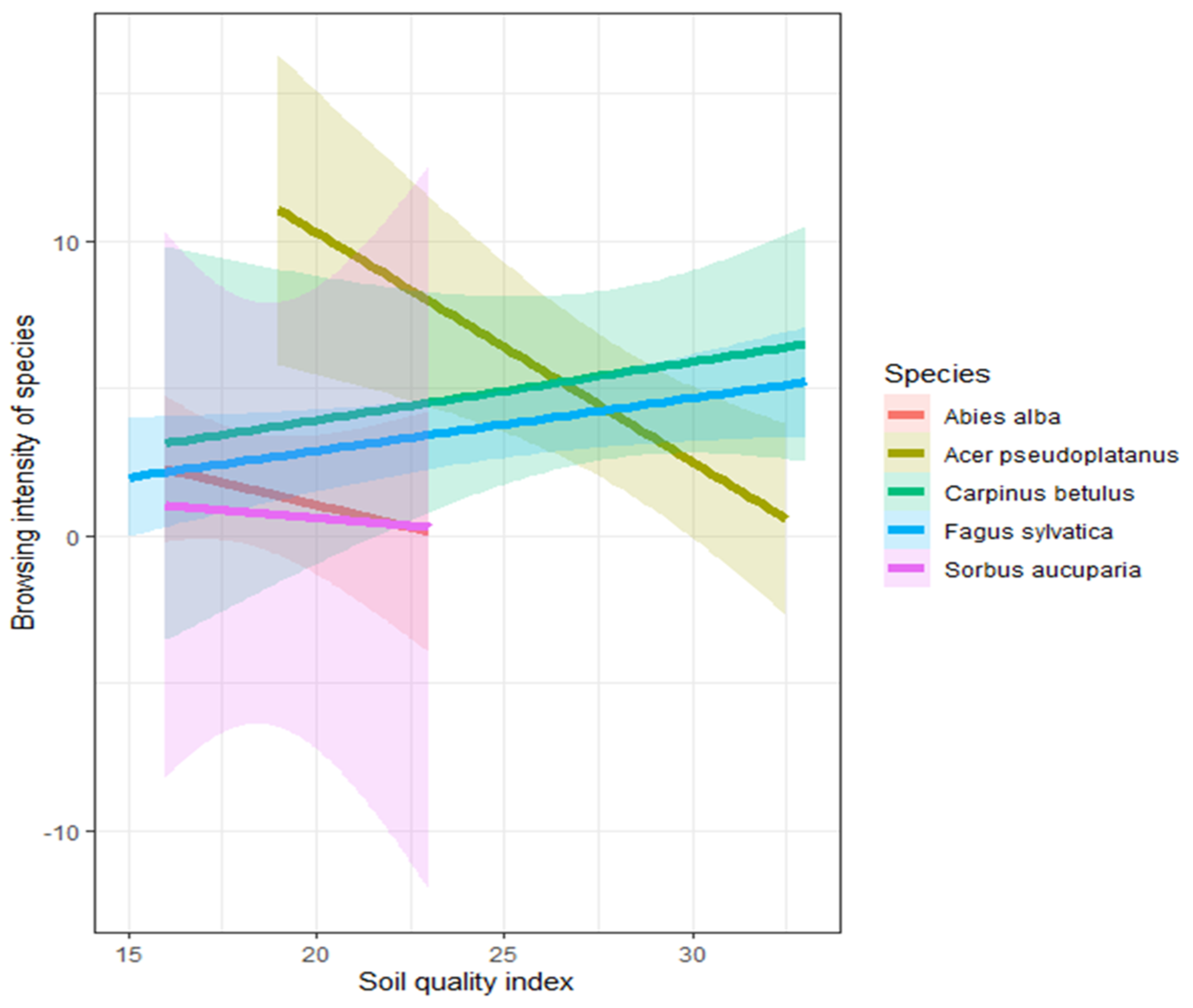
3.4. The Relationship between Soil Quality Index and Browsing Intensity
4. Discussion
4.1. Relationship between Soil Quality Index, Tree Species Relative Density, and Browsing Intensity
4.2. The Relationship between Soil Quality Index, Twig Dry Mass, and Browsing Intensity
4.3. The Relationship between Soil Quality Index and Browsing Intensity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Didion, M.; Kupferschmid, A.D.; Bugmann, H. Long-term effects of ungulate browsing on forest composition and structure. For. Ecol. Manag. 2009, 258, S44–S55. [Google Scholar] [CrossRef]
- Churski, M.; Bubnicki, J.W.; Jędrzejewska, B.; Kuijper, D.P.J.; Cromsigt, J.P.G.M. Brown world forests: Increased ungulate browsing keeps temperate trees in recruitment bottlenecks in resource hotspots. New Phytol. 2017, 214, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, T.; Müller, J. Large-scale reduction of ungulate browsing by managed sport hunting. For. Ecol. Manag. 2010, 260, 1416–1423. [Google Scholar] [CrossRef]
- Angst, J.K.; Kupferschmid, A.D. Assessing Browsing Impact in Beech Forests: The Importance of Tree Responses after Browsing. Diversity 2023, 15, 262. [Google Scholar] [CrossRef]
- Akashi, N.; Unno, A.; Uno, H. The browsing ratio as an index of the impact of deer browsing on tree seedlings. Trees For. People 2022, 8, 100276. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Gazda, A.; Muter, E.; Pielech, R.; Szewczyk, J.; Zięba, A.; Zwijacz-Kozica, T.; Wiertelorz, A.; Pachowicz, T.; Bodziarczyk, J. Effects of species and environmental factors on browsing frequency of young trees in mountain forests affected by natural disturbances. For. Ecol. Manag. 2020, 474, 118364. [Google Scholar] [CrossRef]
- Heuze, P.; Schnitzler, A.; Klein, F. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? For. Ecol. Manag. 2005, 217, 219–228. [Google Scholar] [CrossRef]
- Kupferschmid, A.D. Selective browsing behaviour of ungulates influences the growth of Abies alba differently depending on forest type. For. Ecol. Manag. 2018, 429, 317–326. [Google Scholar] [CrossRef]
- Candaele, R.; Ligot, G.; Licoppe, A.; Lievens, J.; Fichefet, V.; Jonard, M.; André, F.; Lejeune, P. Interspecific Growth Reductions Caused by Wild Ungulates on Tree Seedlings and Their Implications for Temperate Quercus-Fagus Forests. Forests 2023, 14, 1330. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. For. Ecol. Manag. 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Augustine, D.J.; McNaughton, S.J. Ungulate Effects on the Functional Species Composition of Plant Communities: Herbivore Selectivity and Plant Tolerance. J. Wildl. Manag. 1998, 62, 1165–1183. [Google Scholar] [CrossRef]
- Kuijper, D.P.J. Lack of natural control mechanisms increases wildlife-forestry conflict in managed temperate European forest systems. Eur. J. For. Res. 2011, 130, 895–909. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; Cromsigt, J.P.G.M.; Churski, M.; Adam, B.; Jedrzejewska, B.; Jedrzejewski, W. Do ungulates preferentially feed in forest gaps in European temperate forest? For. Ecol. Manag. 2009, 258, 1528–1535. [Google Scholar] [CrossRef]
- Bodziarczyk, J.; Zwijacz-Kozica, T.; Gazda, A.; Szewczyk, J.; Frączek, M.; Zięba, A.; Szwagrzyk, J. Species composition, elevation, and former management type affect browsing pressure on forest regeneration in the Tatra National Park. For. Res. Pap. 2017, 78, 238–247. [Google Scholar] [CrossRef]
- Ristok, C.; Weinhold, A.; Ciobanu, M.; Poeschl, Y.; Roscher, C.; Vergara, F.; Eisenhauer, N.; van Dam, N.M. Plant diversity effects on herbivory are related to soil biodiversity and plant chemistry. J. Ecol. 2023, 111, 412–427. [Google Scholar] [CrossRef]
- Frouz, J.; Toyota, A.; Mudrák, O.; Jílková, V.; Filipová, A.; Cajthaml, T. Effects of soil substrate quality, microbial diversity and community composition on the plant community during primary succession. Soil Biol. Biochem. 2016, 99, 75–84. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, H.; Wang, L.; Ling, F.; Wang, Z.; Li, T.; Huang, J. Topography and soil content contribute to plant community composition and structure in subtropical evergreen-deciduous broadleaved mixed forests. Plant Divers. 2021, 43, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, M.; Walthert, L.; Weber, P. Soil nutrients influence growth response of temperate tree species to drought. J. Ecol. 2016, 104, 377–387. [Google Scholar] [CrossRef]
- Holmes, P.M. Shrubland restoration following woody alien invasion and mining: Effects of topsoil depth, seed source, and fertilizer addition. Restor. Ecol. 2001, 9, 71–84. [Google Scholar] [CrossRef]
- Augusto, L.; Boča, A. Tree functional traits, forest biomass, and tree species diversity interact with site properties to drive forest soil carbon. Nat. Commun. 2022, 13, 1097. [Google Scholar] [CrossRef]
- Jiang, Z.D.; Owens, P.R.; Ashworth, A.J.; Fuentes, B.A.; Thomas, A.L.; Sauer, T.J.; Wang, Q.-B. Evaluating tree growth factors into species-specific functional soil maps for improved agroforestry system efficiency. Agrofor. Syst. 2022, 96, 479–490. [Google Scholar] [CrossRef]
- Kardol, P.; Dickie, I.A.; John, M.G.S.; Husheer, S.W.; Bonner, K.I.; Bellingham, P.J.; Wardle, D.A. Soil-mediated effects of invasive ungulates on native tree seedlings. J. Ecol. 2014, 102, 622–631. [Google Scholar] [CrossRef]
- Sardar, M.F.; Younas, F.; Farooqi, Z.U.R.; Li, Y. Soil nitrogen dynamics in natural forest ecosystem: A review. Front. For. Glob. Change 2023, 6, 1–16. [Google Scholar] [CrossRef]
- Charles, D.C.; Finzi, A.C.; Pacala, S.W.; Burbank, D.H. Causes and consequences of resource hetrogenity in forests: Interespecific variation in light transmission by canopy trees. Can. J. For. Res. 1994, 24, 7823–7830. [Google Scholar]
- Coomes, D.A.; Kunstler, G.; Canham, C.D.; Wright, E. A greater range of shade-tolerance niches in nutrient-rich forests: An explanation for positive richness-productivity relationships? J. Ecol. 2009, 97, 705–717. [Google Scholar] [CrossRef]
- Dirnböck, T.; Grandin, U.; Bernhardt-Römermann, M.; Beudert, B.; Canullo, R.; Forsius, M.; Grabner, M.; Holmberg, M.; Kleemola, S.; Lundin, L.; et al. Forest floor vegetation response to nitrogen deposition in Europe. Glob. Chang. Biol. 2014, 20, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Petren, K. Habitat and Niche, Concept of. Encycl. Biodivers. 2001, 3, 303–315. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G. Habitat suitability modelling and niche theory. J. Appl. Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Cremer, M.; Kern, N.V.; Prietzel, J. Soil organic carbon and nitrogen stocks under pure and mixed stands of European beech, Douglas fir and Norway spruce. For. Ecol. Manag. 2016, 367, 30–40. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C: N ratios in European forest soils. For. Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Gruba, P.; Mulder, J. Tree species affect cation exchange capacity (CEC) and cation binding properties of organic matter in acid forest soils. Sci. Total Environ. 2015, 511, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Dodge, V.J.; Eviner, V.T.; Cushman, J.H. Context-dependent effects of a reintroduced ungulate on soil properties are driven by soil texture, moisture, and herbivore activity. Ecol. Evol. 2020, 10, 10858–10871. [Google Scholar] [CrossRef] [PubMed]
- Fornara, D.A. Ungulate Browsing as and Ecosystem Process: Browser-Plant-Soil Interactions in a Southern African Savanna. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, March 2005. [Google Scholar]
- Liang, W.; Feng, Y.; Li, F.; Zhao, Y.; Zhu, J.; Yao, J.; Chang, C.; Wei, X. Drivers of vegetation and soil determine natural regeneration of a single plantation at different slope positions. Front. For. Glob. Chang. 2023, 6, 1167284. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Bütikofer, L.; Hothorn, T.; Schwyzer, A.; Brang, P. Ungulate species and abundance as well as environmental factors determine the probability of terminal shoot browsing on temperate forest trees. Forests 2020, 11, 764. [Google Scholar] [CrossRef]
- Grabowski, T.; Jóźwiakowski, K.; Bochniak, A.; Micek, A. Changes in the Amount of Rainwater in the Roztocze National Park (Poland) in 2001–2020 and the Possibility of Using Rainwater in the Context of Ongoing Climate Variability. Water 2022, 14, 1334. [Google Scholar] [CrossRef]
- Maciejewski, Z. The role of the Roztocze National Park in shaping and protecting the forest landscapes of the Roztocze region. Hum. Impact Upl. landscapes Lublin Reg. 2010, 109–128. [Google Scholar]
- Madalcho, A.B.; Szwagrzyk, J.; Maciejewski, Z.; Gazda, A.; Bodziarczyk, J. Allometric scaling models for tree saplings: Assessing potential fodder for ungulate herbivores. Trees For. People 2023, 13, 100424. [Google Scholar] [CrossRef]
- Brożek, S.; Zwydak, M. Atlas Gleb Leśnych Polski; State Forests Information Center (CILP): Warsaw, Poland, 2010; pp. 11–12. Available online: https://www.larix.lublin.pl/gleboznawstwo/18929-atlas-gleb-lesnych-polski.html (accessed on 12 February 2024).
- Brożek, S.; Lasota, J.; Blońska, E.; Wanic, T.; Zwydak, M. Waloryzacja siedlisk obszarów górskich na podstawie Siedliskowego Indeksu Glebowego (SIGg). Sylwan 2015, 159, 684–692. [Google Scholar]
- Shen, Y.; Li, J.; Chen, F.; Cheng, R.; Xiao, W.; Wu, L.; Zeng, L. Correlations between forest soil quality and aboveground vegetation characteristics in Hunan Province, China. Front. Plant Sci. 2022, 13, 1009109. [Google Scholar] [CrossRef]
- Diaci, J.; Adamic, T.; Fidej, G.; Rozenbergar, D. Toward a Beech-Dominated Alternative Stable State in Dinaric Mixed Montane Forests: A Long-Term Study of the Pecka Old-Growth Forest. Front. For. Glob. Chang. 2022, 5, 1–11. [Google Scholar] [CrossRef]
- Borowski, Z.; Bartoń, K.; Gil, W.; Wójcicki, A.; Pawlak, B. Factors affecting deer pressure on forest regeneration: The roles of forest roads, visibility and forage availability. Pest Manag. Sci. 2021, 77, 628–634. [Google Scholar] [CrossRef]
- Milligan, H.T.; Koricheva, J. Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: An experimental study. J. Anim. Ecol. 2013, 82, 739–748. [Google Scholar] [CrossRef]
- Knoepp, J.D.; Coleman, D.C.; Crossley, D.A., Jr.; Clark, J.S. Biological indices of soil quality: An ecosystem case study of their use. For. Ecol. Manag. 2000, 138, 357–368. [Google Scholar] [CrossRef]
- Walthert, L.; Meier, E.S. Tree species distribution in temperate forests is more influenced by soil than by climate. Ecol. Evol. 2017, 7, 9473–9484. [Google Scholar] [CrossRef] [PubMed]
- Packham, J.R.; Thomas, P.A.; Atkinson, M.D.; Degen, T. Biological Flora of the British Isles: Fagus sylvatica. J. Ecol. 2012, 100, 1557–1608. [Google Scholar] [CrossRef]
- Gottlieb, R.; Gruntman, M. Can plants integrate information on above-ground competition in their directional responses below ground? Ann. Bot. 2022, 130, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Hein, S.; Collet, C.; Ammer, C.; Le Goff, N.; Skovsgaard, J.P.; Savill, P. A review of growth and stand dynamics of Acer pseudoplatanus L. in Europe: Implications for silviculture. Forestry 2009, 82, 361–385. [Google Scholar] [CrossRef]
- Hawkes, J.C.; Pyatt, D.G.; White, I.M.S. Using Ellenberg Indicator Values to Assess Soil Quality in British Forests from Ground Vegetation: A Pilot Study Author (s): White Published by: British Ecological Society Stable. J. Appl. Ecol. 1997, 34, 375–387. [Google Scholar] [CrossRef]
- Ohse, B.; Seele, C.; Holzwarth, F.; Wirth, C. Different facets of tree sapling diversity influence browsing intensity by deer dependent on spatial scale. Ecol. Evol. 2017, 7, 6779–6789. [Google Scholar] [CrossRef]
- Kobal, M.; Grčman, H.; Zupan, M.; Levanič, T.; Simončič, P.; Kadunc, A.; Hladnik, D. Influence of soil properties on silver fir (Abies alba Mill.) growth in the Dinaric Mountains. For. Ecol. Manag. 2015, 337, 77–87. [Google Scholar] [CrossRef]
- Baranowska, M.; Behnke-Borowczyk, J.; Barzdajn, W.; Szmyt, J.; Korzeniewicz, R.; Łukowski, A.; Memišević-Hodžić, M.; Kartawik, N.; Kowalkowski, W. Effects of nursery production methods on fungal community diversity within soil and roots of Abies alba Mill. Sci. Rep. 2023, 13, 21284. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, S. Role of mycorrhiza and soil fungi in natural regeneration of fir (Abies alba Mill.) in Polish Carpathians and Sudetes. Eur. J. For. Pathol. 1982, 12, 107–112. [Google Scholar] [CrossRef]
- Paluch, J.G.; Stepniewska, H.J. Effect of microsites on the survival, density, and ectomycorrhizal status of shade-tolerant abies alba regeneration attacked by fungal pathogens. Can. J. For. Res. 2012, 42, 720–732. [Google Scholar] [CrossRef]
- Petritan, A.M.; von Lüpke, B.; Petritan, I.C. Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Quality Range | Forest Habitat Type | Trophic Variety of Soil Subtype |
|---|---|---|
| SQI of 4 to 13 | Coniferous forests | Dystrophic |
| SQI of 14 to 23 | Mixed coniferous forests | Oligotrophic |
| SQI of 24 to 33 | Mixed deciduous forests | Mesotrophic |
| SQI of 34 to 40 | Deciduous forests | Eutrophic |
| Variable | Mean | SE | Med. | SD. | Var. | Kurtosis | Skewness | Range | Minimum | Maximum |
|---|---|---|---|---|---|---|---|---|---|---|
| pH_H2O | 4.350 | 0.097 | 4.353 | 0.454 | 0.206 | −0.691 | 0.183 | 1.640 | 3.545 | 5.185 |
| pH_KCl | 3.953 | 0.102 | 4.005 | 0.479 | 0.230 | 0.102 | −0.168 | 2.005 | 2.860 | 4.865 |
| Ca | 3.424 | 0.657 | 2.409 | 3.080 | 9.485 | 1.386 | 1.476 | 10.406 | 0.383 | 10.789 |
| K | 0.300 | 0.045 | 0.243 | 0.212 | 0.045 | −0.562 | 0.673 | 0.677 | 0.061 | 0.738 |
| Mg | 0.250 | 0.037 | 0.223 | 0.172 | 0.030 | −0.070 | 0.998 | 0.555 | 0.058 | 0.613 |
| Na | 0.037 | 0.004 | 0.035 | 0.020 | 0.000 | −0.451 | 0.785 | 0.064 | 0.017 | 0.081 |
| BEC | 4.012 | 0.725 | 3.035 | 3.399 | 11.556 | 1.097 | 1.374 | 11.387 | 0.525 | 11.912 |
| EA | 12.367 | 2.598 | 9.632 | 12.184 | 148.440 | 13.581 | 3.527 | 56.656 | 4.814 | 61.470 |
| CEC | 16.378 | 2.734 | 12.959 | 12.822 | 164.395 | 9.126 | 2.748 | 57.853 | 6.283 | 64.136 |
| BS | 22.767 | 2.955 | 22.042 | 13.858 | 192.056 | −0.414 | 0.477 | 49.332 | 2.813 | 52.145 |
| N | 0.182 | 0.029 | 0.137 | 0.134 | 0.018 | 5.805 | 2.379 | 0.549 | 0.080 | 0.629 |
| OC | 3.201 | 0.770 | 2.010 | 3.613 | 13.056 | 7.982 | 2.879 | 14.561 | 1.181 | 15.742 |
| C:N | 13.852 | 0.745 | 12.987 | 3.493 | 12.202 | 3.221 | 1.822 | 13.312 | 10.206 | 23.518 |
| N2:C | 0.011 | 0.001 | 0.010 | 0.006 | 0.000 | 0.941 | 1.260 | 0.020 | 0.005 | 0.025 |
| Sand | 76.364 | 3.368 | 83.250 | 15.799 | 249.600 | −1.592 | −0.380 | 44.500 | 49.000 | 93.500 |
| Silt | 19.932 | 2.867 | 13.750 | 13.447 | 180.817 | −1.495 | 0.428 | 39.000 | 5.500 | 44.500 |
| Clay | 3.705 | 0.517 | 3.000 | 2.423 | 5.873 | −1.783 | 0.249 | 6.500 | 1.000 | 7.500 |
| (a) Species Relative Density Predicted Using the Soil Quality Index | |||||||
| Species | Explanatory Variable | Estimate | Std. Error | t Value | Pr (>|t|) | Adj. p Value | Adj. R2 |
| A. pseudoplatanus | (Intercept) | 144.13 | 29.434 | 4.897 | 0.001 *** | 0.6247 | |
| Soil quality index | −4.328 | 1.03 | −4.201 | 0.002 ** | 0.012 * | ||
| F. sylvatica | (Intercept) | 25.694 | 24.46 | 1.05 | 0.306 | 0.02297 | |
| Soil quality index | 1.178 | 0.964 | 1.222 | 0.236 | 1.000 | ||
| S. aucuparia | (Intercept) | 35.756 | 43.235 | 0.827 | 0.56 | −0.4333 | |
| Soil quality index | −1.442 | 2.293 | −0.629 | 0.643 | 1.000 | ||
| C. betulus | (Intercept) | 12.849 | 25.602 | 0.502 | 0.626 | −0.03806 | |
| Soil quality index | 0.679 | 0.908 | 0.748 | 0.47 | 1.000 | ||
| A. alba | (Intercept) | 164.358 | 91.109 | 1.804 | 0.169 | 0.2315 | |
| Soil quality index | −7.379 | 4.969 | −1.485 | 0.234 | 1.000 | ||
| (b) Browsing Intensity per Species per Transect Predicted Using Species Relative Density | |||||||
| A. pseudoplatanus | (Intercept) | −0.239 | 0.538 | −0.444 | 0.667 | ||
| Species relative density | 0.188 | 0.016 | 11.622 | 0.000 *** | 0.000 *** | 0.931 | |
| F. sylvatica | (Intercept) | 2.179 | 1.227 | 1.776 | 0.091 . | ||
| Species relative density | 0.028 | 0.02 | 1.4 | 0.177 | 0.884 | 0.044 | |
| S. aucuparia | (Intercept) | 0.065 | 0.021 | 3.093 | 0.199 | ||
| Species relative density | 0.077 | 0.002 | 45.09 | 0.014 * | 0.071 | 0.999 | |
| C. betulus | (Intercept) | −1.576 | 1.472 | −1.071 | 0.307 | ||
| Species relative density | 0.222 | 0.041 | 5.424 | 0.000 *** | 0.001 *** | 0.703 | |
| A. alba | (Intercept) | 0.521 | 0.652 | 0.799 | 0.482 | ||
| Species relative density | 0.036 | 0.015 | 2.306 | 0.104 | 0.522 | 0.519 | |
| Species | Explanatory Variable | Estimate | Std. Error | t Value | Pr (>|t|) | Adj. p Value | Adj R2 |
|---|---|---|---|---|---|---|---|
| A. pseudoplatanus | (Intercept) | 0.148 | 0.034 | 4.392 | 0.002 ** | ||
| Species relative density | 0.004 | 0.001 | 3.782 | 0.004 ** | 0.021 * | 0.571 | |
| F. sylvatica | (Intercept) | 0.12 | 0.036 | 3.293 | 0.004 ** | ||
| Species relative density | 0 | 0.001 | 0.611 | 0.548 | 1.000 | −0.031 | |
| S. aucuparia | (Intercept) | 0.203 | 0.002 | 115.76 | 0.006 ** | ||
| Species relative density | −0.002 | 0 | −12.34 | 0.052 | 0.257 | 0.987 | |
| C. betulus | (Intercept) | 0.209 | 0.06 | 3.497 | 0.005 ** | ||
| Species relative density | 0.002 | 0.002 | 1.05 | 0.316 | 1.000 | 0.008 | |
| A. alba | (Intercept) | 0.406 | 0.079 | 5.113 | 0.015 * | ||
| Species relative density | −0.005 | 0.002 | −2.406 | 0.095 | 0.476 | 0.545 |
| (a) Total Twig Dry Mass of Tree Species (g) m−2 Explained by the Soil Quality Index | |||||||
| Species | Explanatory Variable | Estimate | Std. Error | t Value | Pr (>|t|) | Adj. p Value | Adj. R2 |
| A. pseudoplatanus | (Intercept) | 2.480 | 0.449 | 5.529 | 0.000 *** | 0.6923 | |
| Soil quality index | −0.076 | 0.016 | −4.847 | 0.001 *** | 0.005 ** | ||
| F. sylvatica | (Intercept) | 0.289 | 0.271 | 1.065 | 0.300 | 0.2105 | |
| Soil quality index | 0.027 | 0.011 | 2.569 | 0.018 * | 0.091 | ||
| S. aucuparia | (Intercept) | 0.570 | 0.816 | 0.698 | 0.612 | −0.5718 | |
| Soil quality index | −0.023 | 0.043 | −0.522 | 0.694 | 1.000 | ||
| C. betulus | (Intercept) | 0.072 | 0.098 | 0.729 | 0.481 | −0.0819 | |
| Soil quality index | 0.001 | 0.003 | 0.303 | 0.768 | 1.000 | ||
| A. alba | (Intercept) | 3.197 | 1.867 | 1.712 | 0.185 | 0.2006 | |
| Soil quality index | −0.144 | 0.102 | −1.416 | 0.252 | 1.000 | ||
| (b) Browsing Intensity of Tree Species Explained by the Total Twig Dry Mass of Tree Species m−2 | |||||||
| A. pseudoplatanus | (Intercept) | 1.221 | 1.476 | 0.827 | 0.430 | 0.430 | |
| Twig dry mass (g) m−2 | 8.035 | 2.748 | 2.924 | 0.017 * | 0.084 | ||
| F. sylvatica | (Intercept) | −0.362 | 1.434 | −0.252 | 0.803 | 0.279 | |
| Twig dry mass (g) m−2 | 4.220 | 1.398 | 3.020 | 0.007 ** | 0.034 * | ||
| S. aucuparia | (Intercept) | 0.114 | 0.053 | 2.149 | 0.277 | 0.993 | |
| Twig dry mass (g) m−2 | 4.272 | 0.249 | 17.185 | 0.037 * | 0.185 | ||
| C. betulus | (Intercept) | −0.646 | 1.215 | −0.532 | 0.605 | 0.741 | |
| Twig dry mass (g) m−2 | 60.334 | 10.141 | 5.949 | 0.000 *** | 0.004 *** | ||
| A. alba | (Intercept) | 0.631 | 0.692 | 0.912 | 0.429 | 0.429 | |
| Twig dry mass (g) m−2 | 1.682 | 0.840 | 2.002 | 0.139 | 0.695 | ||
| Species | Explanatory Variable | Estimate | Std. Error | t Value | Pr (>|t|) | Adj. p Values | Adj. R2 |
|---|---|---|---|---|---|---|---|
| A. pseudoplatanus | (Intercept) | 25.845 | 6.429 | 4.02 | 0.003 ** | ||
| Soil quality index | −0.778 | 0.225 | −3.458 | 0.007 ** | 0.036 * | 0.523 | |
| F. sylvatica | (Intercept) | −0.707 | 2.125 | −0.332 | 0.743 | ||
| Soil quality index | 0.179 | 0.084 | 2.143 | 0.045 * | 0.027 * | 0.146 | |
| S. aucuparia | (Intercept) | 2.75 | 3.381 | 0.813 | 0.565 | ||
| Soil quality index | −0.107 | 0.179 | −0.598 | 0.657 | 1.000 | −0.473 | |
| C. betulus | (Intercept) | −0.024 | 6.611 | −0.004 | 0.997 | ||
| Soil quality index | 0.198 | 0.234 | 0.843 | 0.417 | 1.000 | −0.025 | |
| A. alba | (Intercept) | 7.138 | 4.284 | 1.666 | 0.194 | ||
| Soil quality index | −0.305 | 0.234 | −1.304 | 0.283 | 1.000 | 0.149 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madalcho, A.B.; Gazda, A.; Wanic, T.; Szwagrzyk, J. Influence of Soil Quality on the Browsing Intensity of Ungulate Herbivores on Tree Species in European Forests. Forests 2024, 15, 708. https://doi.org/10.3390/f15040708
Madalcho AB, Gazda A, Wanic T, Szwagrzyk J. Influence of Soil Quality on the Browsing Intensity of Ungulate Herbivores on Tree Species in European Forests. Forests. 2024; 15(4):708. https://doi.org/10.3390/f15040708
Chicago/Turabian StyleMadalcho, Aklilu Bajigo, Anna Gazda, Tomasz Wanic, and Jerzy Szwagrzyk. 2024. "Influence of Soil Quality on the Browsing Intensity of Ungulate Herbivores on Tree Species in European Forests" Forests 15, no. 4: 708. https://doi.org/10.3390/f15040708
APA StyleMadalcho, A. B., Gazda, A., Wanic, T., & Szwagrzyk, J. (2024). Influence of Soil Quality on the Browsing Intensity of Ungulate Herbivores on Tree Species in European Forests. Forests, 15(4), 708. https://doi.org/10.3390/f15040708