
The Relationships between Biomass and Soil Respiration across Different Forest Management Practices


Abstract
:1. Introduction
2. Materials and Methods
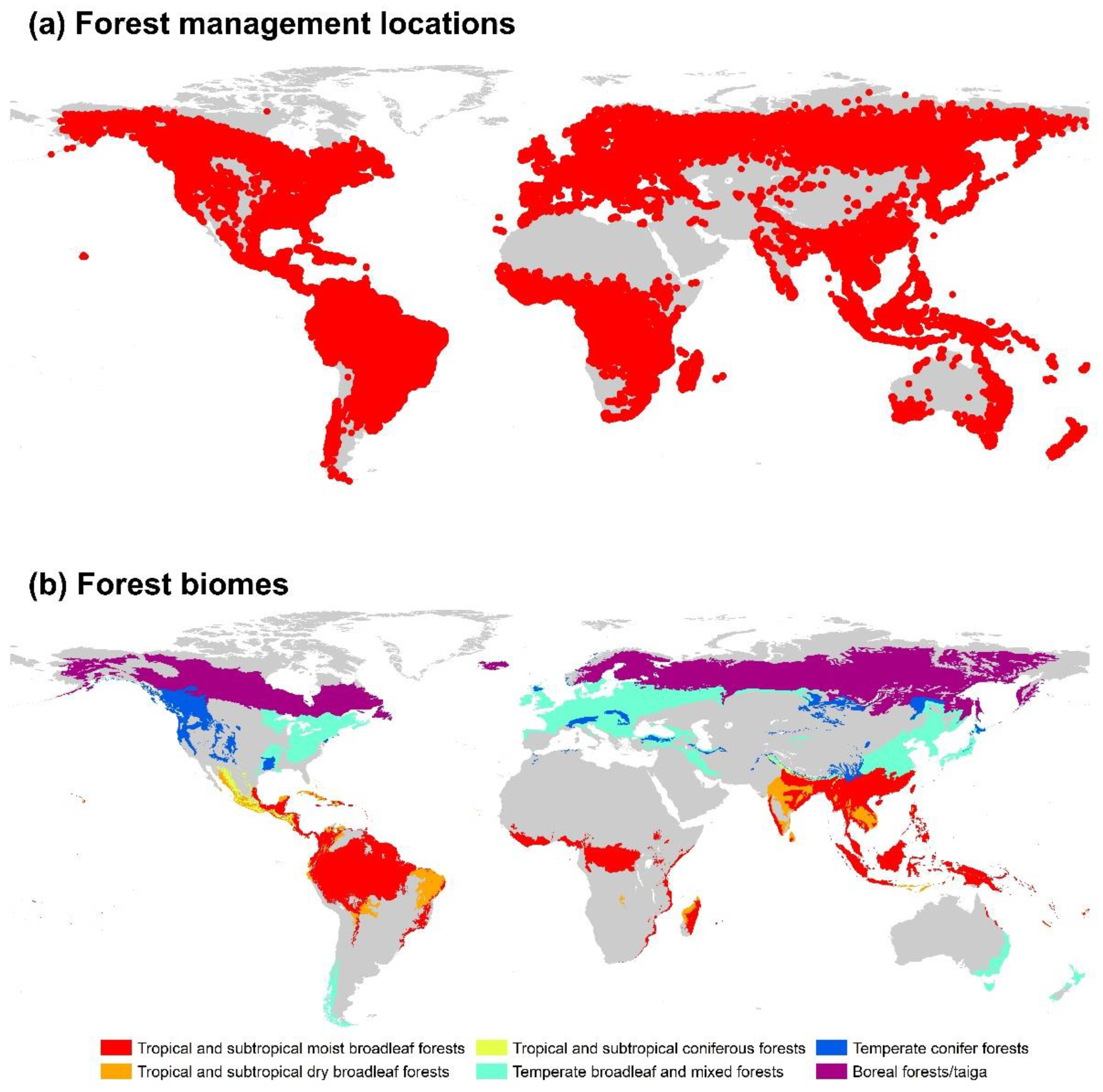
2.1. Data on Forest Management Practices
2.2. Data on Soil Respiration
2.3. Data on Forest Biomass
2.4. Data on Soil Temperature and Moisture
2.5. Analyses
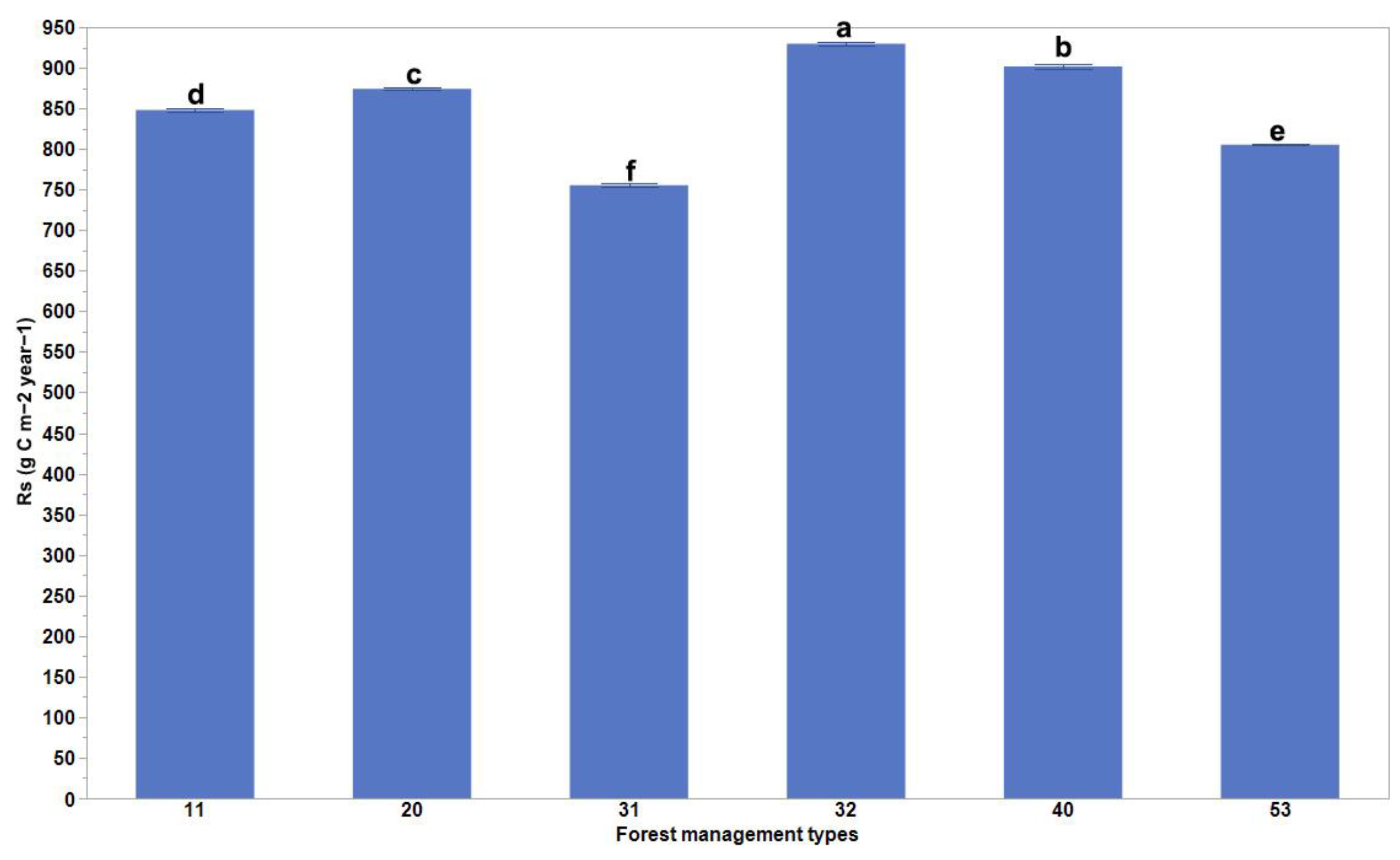
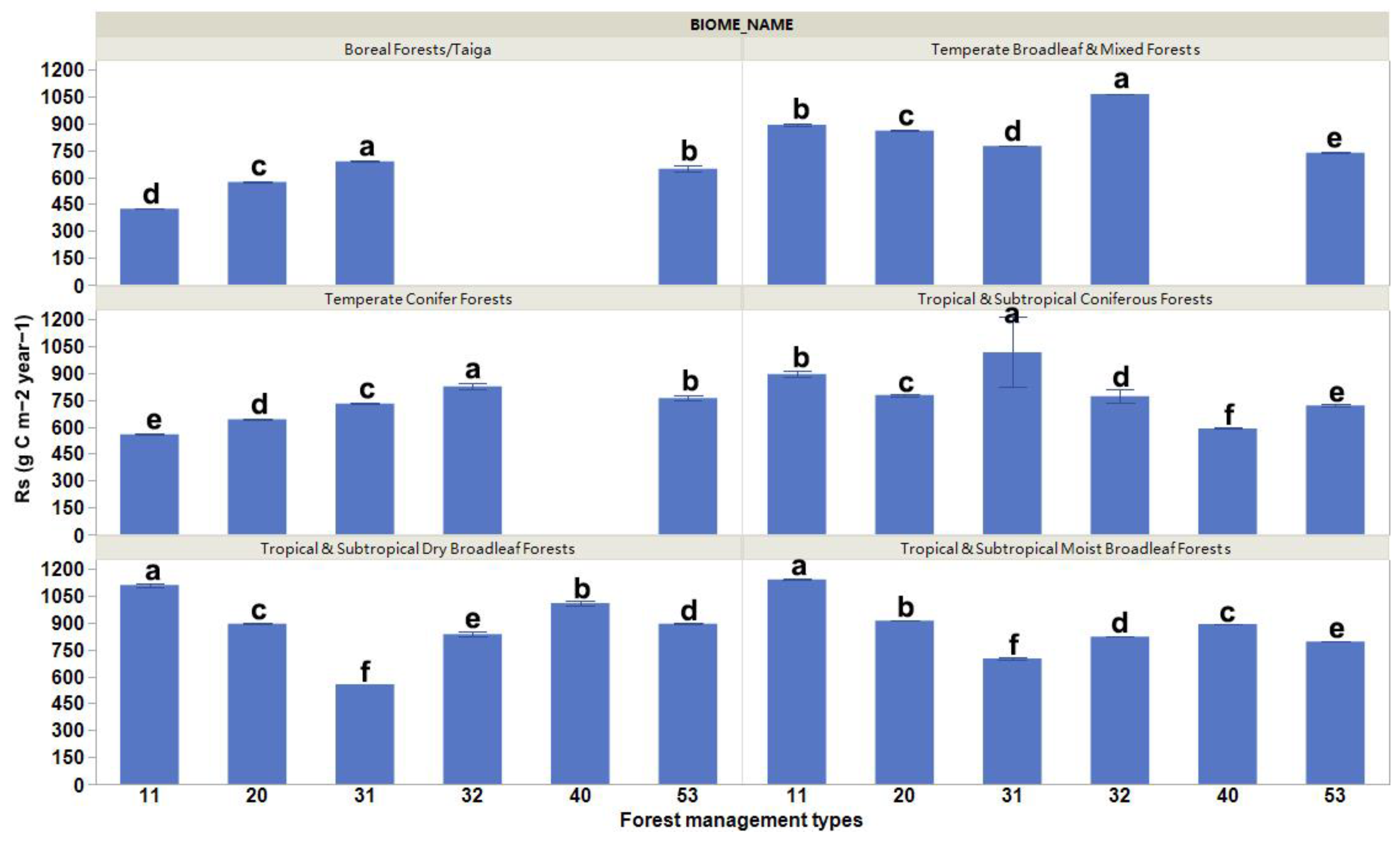
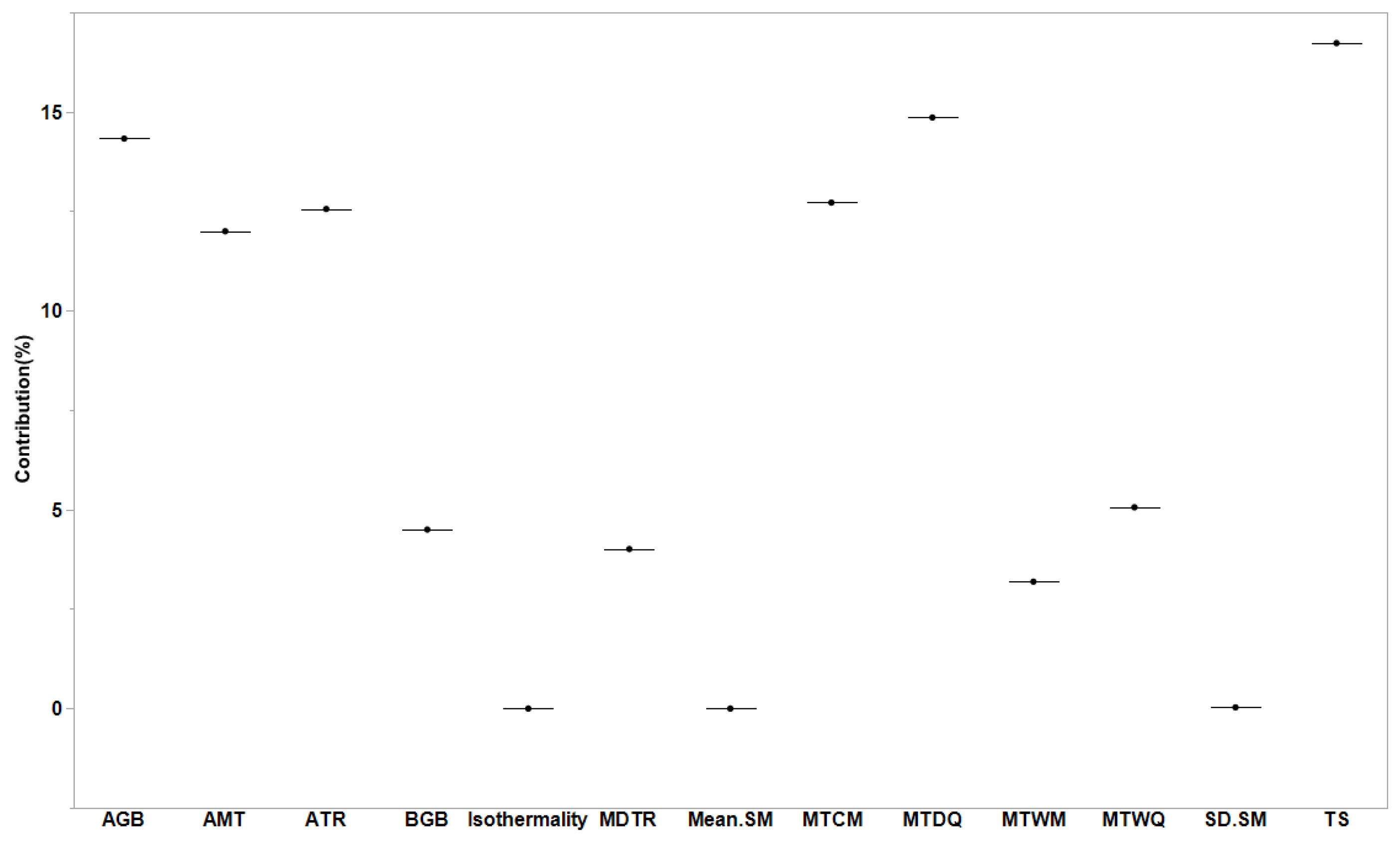
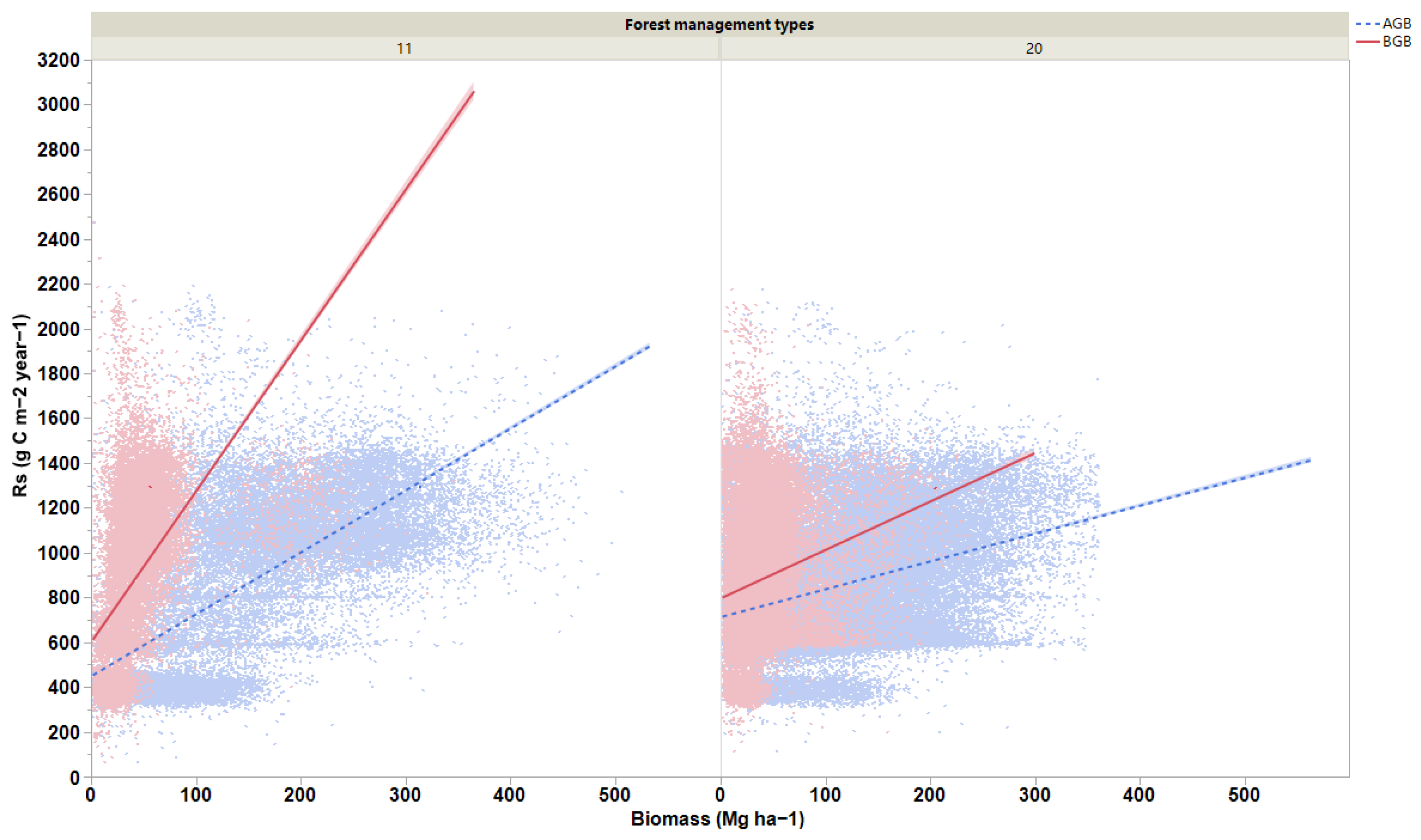
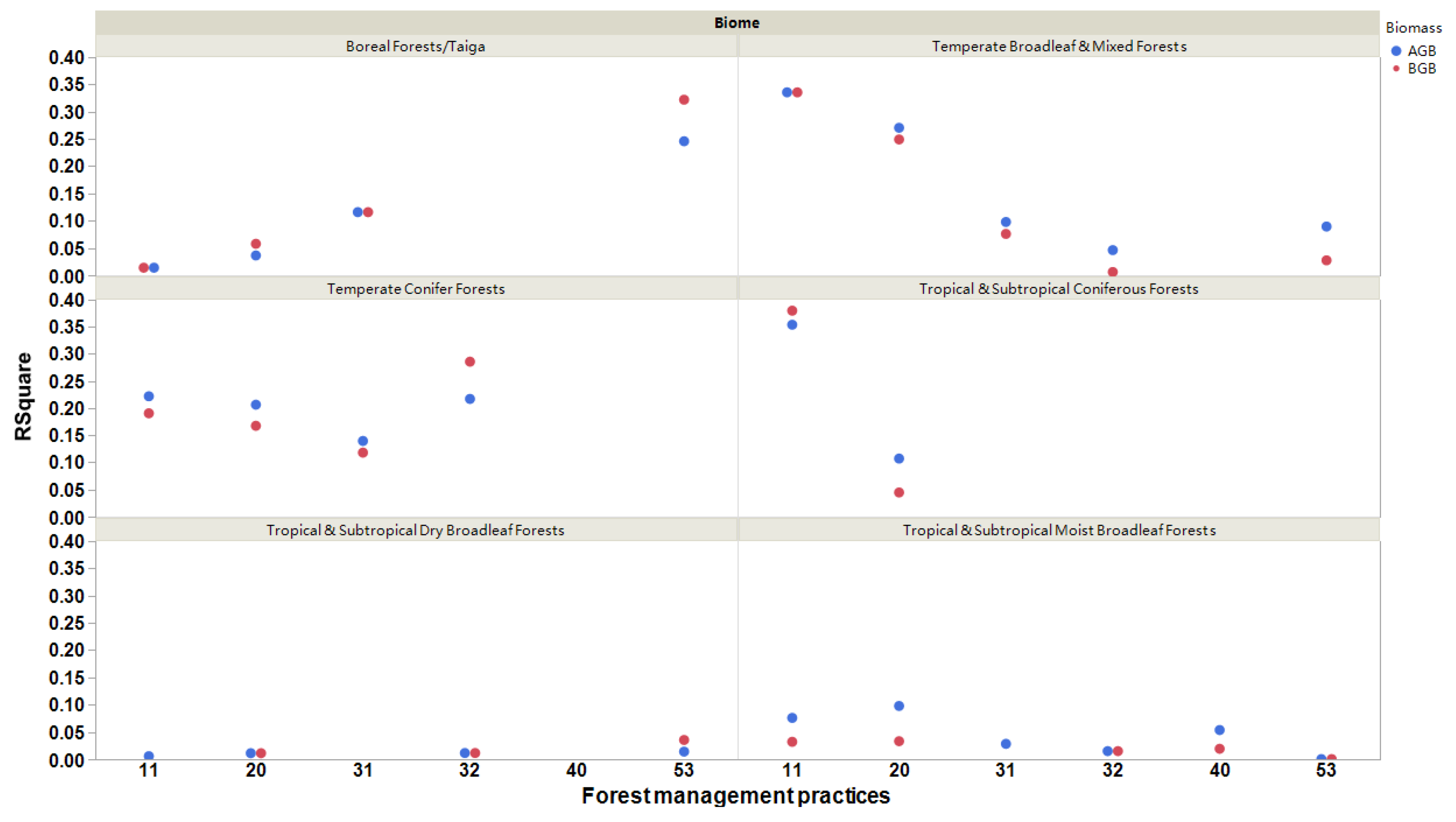
3. Results
4. Discussion
4.1. Relationships between Biomass and Soil Respiration
4.2. Relationships between Soil Temperature and Moisture and Soil Respiration
4.3. Biome Effects
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lesiv, M.; Schepaschenko, D.; Buchhorn, M.; See, L.; Dürauer, M.; Georgieva, I.; Jung, M.; Hofhansl, F.; Schulze, K.; Bilous, A.; et al. Global forest management data for 2015 at a 100 m resolution. Sci. Data 2022, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Peng, C. Understanding the role of forest simulation models in sustainable forest management. Environ. Impact Assess. Rev. 2000, 20, 481–501. [Google Scholar] [CrossRef]
- Varma, V.K.; Ferguson, I.; Wild, I. Decision support system for the sustainable forest management. For. Ecol. Manag. 2000, 128, 49–55. [Google Scholar] [CrossRef]
- Colombo, F.; Macdonald, C.A.; Jeffries, T.C.; Powell, J.R.; Singh, B.K. Impact of forest management practices on soil bacterial diversity and consequences for soil processes. Soil Biol. Biochem. 2016, 94, 200–210. [Google Scholar] [CrossRef]
- Muchane, M.N.; Sileshi, G.W.; Gripenberg, S.; Jonsson, M.; Pumariño, L.; Barrios, E. Agroforestry boosts soil health in the humid and sub-humid tropics: A meta-analysis. Agric. Ecosyst. Environ. 2020, 295, 106899. [Google Scholar] [CrossRef]
- Fahad, S.; Chavan, S.B.; Chichaghare, A.R.; Uthappa, A.R.; Kumar, M.; Kakade, V.; Pradhan, A.; Jinger, D.; Rawale, G.; Yadav, D.K.; et al. Agroforestry systems for soil health improvement and maintenance. Sustainability 2022, 14, 14877. [Google Scholar] [CrossRef]
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Du, X.; Stewart, R.D. A database for global soil health assessment. Sci. Data 2020, 7, 16. [Google Scholar] [CrossRef]
- Carbone, M.S.; Still, C.J.; Ambrose, A.R.; Dawson, T.E.; Williams, A.P.; Boot, C.M.; Schaeffer, S.M.; Schimel, J.P. Seasonal and episodic moisture controls on plant and microbial contributions to soil respiration. Oecologia 2011, 167, 265–278. [Google Scholar] [CrossRef]
- Phillips, C.L.; Bond-Lamberty, B.; Desai, A.R.; Lavoie, M.; Risk, D.; Tang, J.; Todd-Brown, K.; Vargas, R. The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling. Plant Soil 2017, 413, 1–25. [Google Scholar] [CrossRef]
- Peng, Y.; Thomas, S.C.; Tian, D. Forest management and soil respiration: Implications for carbon sequestration. Environ. Rev. 2008, 16, 93–111. [Google Scholar] [CrossRef]
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. Soil respiration within riparian buffers and adjacent crop fields. Plant Soil 2001, 229, 117–124. [Google Scholar] [CrossRef]
- Hynynen, J.; Salminen, H.; Ahtikoski, A.; Huuskonen, S.; Ojansuu, R.; Siipilehto, J.; Lehtonen, M.; Eerikäinen, K. Long-term impacts of forest management on biomass supply and forest resource development: A scenario analysis for Finland. Eur. J. For. Res. 2015, 134, 415–431. [Google Scholar] [CrossRef]
- Erb, K.-H.; Kastner, T.; Plutzar, C.; Bais, A.L.S.; Carvalhais, N.; Fetzel, T.; Gingrich, S.; Haberl, H.; Lauk, C.; Niedertscheider, M.; et al. Unexpectedly large impact of forest management and grazing on global vegetation biomass. Nature 2018, 553, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, P.J.; Fitzgerald, J.B.; Datta, P.; Dees, M.; Hengeveld, G.M.; Lindner, M.; Zudin, S. Spatial distribution of the potential forest biomass availability in Europe. For. Ecosyst. 2019, 6, 5. [Google Scholar] [CrossRef]
- Dornbush, M.E.; Raich, J.W. Soil temperature, not aboveground plant productivity, best predicts intra-annual variations of soil respiration in central Iowa grasslands. Ecosystems 2006, 9, 909–920. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Fisher, R.A.; Wardle, D.A. Plant communities as drivers of soil respiration: Pathways, mechanisms, and significance for global change. Biogeosciences 2011, 8, 2047–2061. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y. Plant diversity loss reduces soil respiration across terrestrial ecosystems. Glob. Chang. Biol. 2019, 25, 1482–1492. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Liu, L.; Wang, X.; Lajeunesse, M.J.; Miao, G.; Piao, S.; Wan, S.; Wu, Y.; Wang, Z.; Yang, S.; Li, P.; et al. A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes. Glob. Chang. Biol. 2016, 22, 1394–1405. [Google Scholar] [CrossRef] [PubMed]
- Major, J.; Lehmann, J.; Rondon, M.; Goodale, C. Fate of soil-applied black carbon: Downward migration, leaching and soil respiration. Glob. Chang. Biol. 2010, 16, 1366–1379. [Google Scholar] [CrossRef]
- Zhang, H.; Ying, B.; Hu, Y.; Wang, Y.; Yu, X.; Tang, C. Response of soil respiration to thinning is altered by thinning residue treatment in Cunninghamia lanceolata plantations. Agric. For. Meteorol. 2022, 324, 109089. [Google Scholar] [CrossRef]
- Zhao, B.; Ballantyne, A.P.; Meng, S.; Zhao, G.; Zheng, Z.; Zhu, J. Understory plant removal counteracts tree thinning effect on soil respiration in a temperate forest. Glob. Chang. Biol. 2022, 28, 6102–6113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Han, H.; Kang, F.; Liu, K.; Song, Y.; Zhou, B.; Li, Y. Short-term effects of thinning on soil respiration in a pine (Pinus tabulaeformis) plantation. Biol. Fertil. Soils 2014, 50, 357–367. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Ren, C.; Chen, J.; Deng, J.; Zhao, F.; Han, X.; Yang, G.; Tong, X.; Feng, Y.; Shelton, S.; Ren, G. Response of microbial diversity to C: N: P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fertil. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y.; et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef]
- Lavigne, M.B.; Foster, R.J.; Goodine, G. Seasonal and annual changes in soil respiration in relation to soil temperature, water potential and trenching. Tree Physiol. 2004, 24, 415–424. [Google Scholar] [CrossRef]
- Crous, K.Y.; Uddling, J.; De Kauwe, M.G. Temperature responses of photosynthesis and respiration in evergreen trees from boreal to tropical latitudes. New Phytol. 2022, 234, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fu, B.; Zhang, L.; Xu, Z. Soil moisture–plant interactions: An ecohydrological review. J. Soils Sediments 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Lobell, D.B.; Asner, G.P. Moisture effects on soil reflectance. Soil Sci. Soc. Am. J. 2002, 66, 722–727. [Google Scholar] [CrossRef]
- Ainiwaer, M.; Ding, J.; Kasim, N.; Wang, J.; Wang, J. Regional scale soil moisture content estimation based on multi-source remote sensing parameters. Int. J. Remote Sens. 2020, 41, 3346–3367. [Google Scholar] [CrossRef]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Marquet, P.A. Environmental heterogeneity as a driver of terrestrial biodiversity on a global scale. Prog. Phys. Geogr. 2023, 47, 912–930. [Google Scholar] [CrossRef]
- Conradi, T.; Slingsby, J.A.; Midgley, G.F.; Nottebrock, H.; Schweiger, A.H.; Higgins, S.I. An operational definition of the biome for global change research. New Phytol. 2020, 227, 1294–1306. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Qi, X.; Zhang, R.; He, M.; Li, J.; Xu, R.; Li, Y.; Sarmah, S.; Wang, H.; Zhao, J. A Novel Approach for Ecosystem Respiration Simulation in Drylands. Front. Ecol. Evol. 2023, 11, 1186272. [Google Scholar] [CrossRef]
- Huang, N.; Wang, L.; Song, X.P.; Black, T.A.; Jassal, R.S.; Myneni, R.B.; Wu, C.; Song, W.; Ji, D.; Yu, S.; et al. Spatial and temporal variations in global soil respiration and their relationships with climate and land cover. Sci. Adv. 2020, 6, eabb8508. [Google Scholar] [CrossRef]
- Santoro, M.; Cartus, O.; Carvalhais, N.; Rozendaal, D.; Avitabilie, V.; Araza, A.; de Bruin, S.; Herold, M.; Quegan, S.; Rodríguez-Veiga, P.; et al. The global forest above-ground biomass pool for 2010 estimated from high-resolution satellite observations. Earth Syst. Sci. Data Discuss. 2020, 13, 3927–3950. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Hoogen, J.v.D.; Aalto, J.; Ashcroft, M.B.; De Frenne, P.; Kemppinen, J.; Kopecký, M.; Luoto, M.; Maclean, I.M.D.; Crowther, T.W.; et al. Global maps of soil temperature. Glob. Chang. Biol. 2021, 28, 3110–3144. [Google Scholar] [CrossRef]
- Guevara, M.; Taufer, M.; Vargas, R. Gap-free global annual soil moisture: 15 km grids for 1991–2018. Earth Syst. Sci. Data 2021, 13, 1711–1735. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, S.; Zhang, X.; Mao, L. glmm. hp: An R package for computing individual effect of predictors in generalized linear mixed models. J. Plant Ecol. 2022, 15, 1302–1307. [Google Scholar] [CrossRef]
- Akinremi, O.O.; McGinn, S.M.; McLean, H.D.J. Effects of soil temperature and moisture on soil respiration in barley and fallow plots. Can. J. Soil Sci. 1999, 79, 5–13. [Google Scholar] [CrossRef]
- Curiel Yuste, J.; Baldocchi, D.D.; Gershenson, A.; Goldstein, A.; Misson, L.; Wong, S. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Glob. Chang. Biol. 2007, 13, 2018–2035. [Google Scholar] [CrossRef]
- Tang, X.L.; Zhou, G.Y.; Liu, S.G.; Zhang, D.Q.; Liu, S.Z.; Li, J.; Zhou, C.Y. Dependence of soil respiration on soil temperature and soil moisture in successional forests in southern China. J. Integr. Plant Biol. 2006, 48, 654–663. [Google Scholar] [CrossRef]
- Von Arx, G.; Graf Pannatier, E.; Thimonier, A.; Rebetez, M. Microclimate in forests with varying leaf area index and soil moisture: Potential implications for seedling establishment in a changing climate. J. Ecol. 2013, 101, 1201–1213. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Yan, Y.; Quan, Q.; Meng, C.; Wang, J.; Tian, D.; Wang, B.; Zhang, R.; Niu, S. Varying soil respiration under long-term warming and clipping due to shifting carbon allocation toward below-ground. Agric. For. Meteorol. 2021, 304, 108408. [Google Scholar] [CrossRef]
- Heinen, R.; Thakur, M.P.; Hiddes De Fries, J.R.; Steinauer, K.; Vandenbrande, S.; Jongen, R.; Bezemer, T.M. Foliar herbivory on plants creates soil legacy effects that impact future insect herbivore growth via changes in plant community biomass allocation. Funct. Ecol. 2022, 36, 1047–1062. [Google Scholar] [CrossRef]
- Cisneros-Dozal, L.M.; Trumbore, S.E.; Hanson, P.J. Effect of moisture on leaf litter decomposition and its contribution to soil respiration in a temperate forest. J. Geophys. Res. 2007, 112, G01013. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Zhao, R.; Guo, Y.; Hao, L. Aboveground net primary productivity and soil respiration display different responses to precipitation changes in desert grassland. J. Plant Ecol. 2021, 15, 57–70. [Google Scholar] [CrossRef]
- Valentini, C.M.A.; Sanches, L.; de Paula, S.R.; Vourlitis, G.L.; de Souza Nogueira, J.; Pinto, O.B., Jr.; de Almeida Lobo, F. Soil respiration and aboveground litter dynamics of a tropical transitional forest in northwest Mato Grosso, Brazil. J. Geophys. Res. 2008, 113, G00B10. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Dacal, M.; García-Palacios, P.; Asensio, S.; Cano-Díaz, C.; Gozalo, B.; Ochoa, V.; Maestre, F.T. Contrasting mechanisms underlie short-and longer-term soil respiration responses to experimental warming in a dryland ecosystem. Glob. Chang. Biol. 2020, 26, 5254–5266. [Google Scholar] [CrossRef]
- Bae, K.; Lee, D.K.; Fahey, T.J.; Woo, S.Y.; Quaye, A.K.; Lee, Y.K. Seasonal variation of soil respiration rates in a secondary forest and agroforestry systems. Agrofor. Syst. 2013, 87, 131–139. [Google Scholar] [CrossRef]
- Pang, X.; Bao, W.; Zhu, B.; Cheng, W. Responses of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agric. For. Meteorol. 2013, 171, 57–64. [Google Scholar] [CrossRef]
- Wang, C.-J.; Wang, R.; Yu, C.-M.; Pubu, Y.; Sun, W.-G.; Dang, X.-F.; Li, Q.-F.; Wan, J.-Z. Determinants of species assemblages of insect pests in alpine forest ecosystems of western China. For. Ecosyst. 2021, 8, 71. [Google Scholar] [CrossRef]
- Epron, D.; Nouvellon, Y.; Roupsard, O.; Mouvondy, W.; Mabiala, A.; Saint-André, L.; Joffre, R.; Jourdan, C.; Bonnefond, J.-M.; Berbigier, P.; et al. Spatial and temporal variations of soil respiration in a Eucalyptus plantation in Congo. For. Ecol. Manag. 2004, 202, 149–160. [Google Scholar] [CrossRef]
- Yang, L.; Qin, J.; Geng, Y.; Zhang, C.; Pan, J.; Niu, S.; Tian, D.; Zhao, X.; Wang, J. Long-term effects of forest thinning on soil respiration and its components in a pine plantation. For. Ecol. Manag. 2022, 513, 120189. [Google Scholar] [CrossRef]
- Hua, F.; Bruijnzeel, L.A.; Meli, P.; Martin, P.A.; Zhang, J.; Nakagawa, S.; Miao, X.; Wang, W.; McEvoy, C.; Peña-Arancibia, J.L.; et al. The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022, 376, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Messier, C.; Bauhus, J.; Sousa-Silva, R.; Auge, H.; Baeten, L.; Barsoum, N.; Bruelheide, H.; Caldwell, B.; Cavender-Bares, J.; Dhiedt, E.; et al. For the sake of resilience and multifunctionality, let’s diversify planted forests! Conserv. Lett. 2022, 15, e12829. [Google Scholar] [CrossRef]
- Ahlström, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Schaeffer, S.M.; Katul, G.; Porporato, A.; Schimel, J.P. A theoretical analysis of microbial eco-physiological and diffusion limitations to carbon cycling in drying soils. Soil Biol. Biochem. 2014, 73, 69–83. [Google Scholar] [CrossRef]
- Misiak, M.; Goodall-Copestake, W.P.; Sparks, T.H.; Worland, M.R.; Boddy, L.; Magan, N.; Convey, P.; Hopkins, D.W.; Newsham, K.K. Inhibitory effects of climate change on the growth and extracellular enzyme activities of a widespread Antarctic soil fungus. Glob. Chang. Biol. 2021, 27, 1111–1125. [Google Scholar] [CrossRef] [PubMed]
- Concilio, A.; Ma, S.; Ryu, S.R.; North, M.; Chen, J. Soil respiration response to experimental disturbances over 3 years. For. Ecol. Manag. 2006, 228, 82–90. [Google Scholar] [CrossRef]
- Hagemann, U.; Moroni, M.T.; Gleißner, J.; Makeschin, F. Disturbance history influences downed woody debris and soil respiration. For. Ecol. Manag. 2010, 260, 1762–1772. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Jia, X.; Mu, Y.; Zha, T.; Wang, B.; Qin, S.; Tian, Y. Seasonal and interannual variations in ecosystem respiration in relation to temperature, moisture, and productivity in a temperate semi-arid shrubland. Sci. Total Environ. 2020, 709, 136210. [Google Scholar] [CrossRef]
- Jian, J.; Bahn, M.; Wang, C.; Bailey, V.L.; Bond-Lamberty, B. Prediction of annual soil respiration from its flux at mean annual temperature. Agric. For. Meteorol. 2020, 287, 107961. [Google Scholar] [CrossRef]
- Tang, J.; Qi, Y.; Xu, M.; Misson, L.; Goldstein, A.H. Forest thinning and soil respiration in a ponderosa pine plantation in the Sierra Nevada. Tree Physiol. 2005, 25, 57–66. [Google Scholar] [CrossRef]
- Wang, Z.; Ji, L.; Hou, X.; Schellenberg, M.P. Soil respiration in semiarid temperate grasslands under various land management. PLoS ONE 2016, 11, e0147987. [Google Scholar]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Salomón, R.; Barba, J.; Gordaliza, G.G.; Curiel Yuste, J.; Magro, C.; Gil, L. Regeneration in the understory of declining overstory trees contributes to soil respiration homeostasis along succession in a sub-Mediterranean beech forest. Forests 2019, 10, 727. [Google Scholar] [CrossRef]
- Jose, S.; Bardhan, S. Agroforestry for biomass production and carbon sequestration: An overview. Agrofor. Syst. 2012, 86, 105–111. [Google Scholar] [CrossRef]
- Kukumägi, M.; Ostonen, I.; Kupper, P.; Truu, M.; Tulva, I.; Varik, M.; Aosaar, J.; Sõber, J.; Lõhmus, K. The effects of elevated atmospheric humidity on soil respiration components in a young silver birch forest. Agric. For. Meteorol. 2014, 194, 167–174. [Google Scholar] [CrossRef]
- Ma, Y.Q.; Wang, C.J.; Chen, Z.; Yu, F.H.; Wan, J.Z. Linking Forest Management Practices to the Functional Composition of Plant Communities. Forests 2023, 14, 1939. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 11 | 20 | 31 | 32 | 40 | 53 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | Slope | R2 | Slope | R2 | Slope | R2 | Slope | R2 | Slope | R2 | Slope | |
| AMT | 0.72 | 29.20 | 0.06 | 8.21 | 0.04 | 7.27 | 0.26 | −19.81 | 0.00 | −4.69 | 0.01 | 6.59 |
| MDTR | 0.34 | −84.33 | 0.04 | −20.45 | 0.06 | −22.11 | 0.00 | 7.41 | 0.02 | −24.31 | 0.00 | −1.73 |
| Isothermality | 0.00 | 0.00 | 0.000 | 0.00 | 0.02 | −3.01 | 0.00 | 0.010 | 0.00 | −0.01 | 0.00 | 0.00 |
| TS | 0.72 | −0.89 | 0.12 | −0.41 | 0.09 | −0.30 | 0.02 | 0.22 | 0.03 | −1.04 | 0.01 | −0.12 |
| MTWM | 0.34 | 34.87 | 0.06 | 3.26 | 0.00 | −1.72 | 0.27 | −20.83 | 0.00 | −2.79 | 0.01 | 4.56 |
| MTCM | 0.73 | 19.44 | 0.07 | 6.65 | 0.11 | 12.27 | 0.17 | −12.60 | 0.01 | 16.42 | 0.02 | 5.30 |
| ATR | 0.66 | −23.74 | 0.09 | −10.11 | 0.10 | −9.75 | 0.00 | 1.74 | 0.00 | −4.87 | 0.00 | −1.20 |
| MTWQ | 0.49 | 38.21 | 0.02 | 6.10 | 0.00 | −0.54 | 0.25 | −15.59 | 0.00 | −9.38 | 0.00 | 3.05 |
| MTDQ | 0.72 | 21.12 | 0.07 | 7.19 | 0.05 | 4.562 | 0.17 | −20.56 | 0.00 | −11.24 | 0.01 | 5.91 |
| Mean.SM | 0.41 | 5501.12 | 0.00 | 0.00 | 0.03 | 741.47 | 0.00 | −0.06 | 0.02 | 1190.86 | 0.00 | 0.00 |
| SD.SM | 0.00 | −0.06 | 0.00 | 0.00 | 0.00 | 522.05 | 0.00 | 0.02 | 0.00 | 0.05 | 0.00 | −0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Lei, J.-P.; Wan, J.-Z. The Relationships between Biomass and Soil Respiration across Different Forest Management Practices. Forests 2024, 15, 712. https://doi.org/10.3390/f15040712
Hu C, Lei J-P, Wan J-Z. The Relationships between Biomass and Soil Respiration across Different Forest Management Practices. Forests. 2024; 15(4):712. https://doi.org/10.3390/f15040712
Chicago/Turabian StyleHu, Chen, Jing-Pin Lei, and Ji-Zhong Wan. 2024. "The Relationships between Biomass and Soil Respiration across Different Forest Management Practices" Forests 15, no. 4: 712. https://doi.org/10.3390/f15040712
APA StyleHu, C., Lei, J.-P., & Wan, J.-Z. (2024). The Relationships between Biomass and Soil Respiration across Different Forest Management Practices. Forests, 15(4), 712. https://doi.org/10.3390/f15040712