1. Introduction
Non-native plants can profoundly impact ecological communities by reducing native biodiversity and altering ecosystem function [
1,
2,
3]. By modifying the biotic (e.g., prey availability) and abiotic characteristics (e.g., structure, light availability, and soil chemistry) of a native species’ habitat, non-native plants may affect individual fitness and ultimately population or species persistence [
4,
5,
6]. Because impacts on individual species can radiate through food webs, non-native plants can also alter the availability, distribution, and quality of resources for consumers at other trophic levels [
7,
8,
9]. For example, non-native plants compete with and suppress the growth of native plants, altering the pre-existing vegetative community [
10,
11]. Such changes can directly and indirectly reduce the survival and reproductive success of some native herbivorous insects, which subsequently impact insectivores and their predators [
12]. By potentially reducing native biodiversity, non-native plants can also diminish the ecosystem functions that native species perform, such as pollination and tree regeneration, which may consequently alter the succession dynamics of a landscape [
13,
14,
15,
16].
Forests, particularly those in the eastern temperate zone, are highly susceptible to invasions of non-native plants due to such human-induced stressors as forest fragmentation and soil disturbance [
17,
18]. Non-native shrubs typically increase understory clutter [
17], which could affect how forest-dwelling wildlife use the landscape. Although non-native plants in some cases have been shown to have little impact on native fauna [
19], non-native plants often force animals to adapt or suffer negative fitness consequences [
16]. Indeed, native eastern North American butterflies, birds, and snakes have been shown to alter their habitat use in response to non-native plants, often resulting in increased mortality. The West Virginia white (
Pieris virginiensis, Edwards), an imperiled native butterfly, preferentially oviposits on non-native garlic mustard (
Alliaria petiolate, Cavara & Grande), despite the plant being toxic to its caterpillars [
20,
21]. Neotropical migrant birds, such as Kentucky Warblers (
Geothlypis Formosa, Wilson) and Hooded Warblers (
Setophaga citrina, Boddaert), will not nest in areas dominated by non-native plants [
22], leading to decreased reproductive success. Further, nonmigratory birds that do build nests in non-native shrubs experience higher daily mortality and nest failure due to predation than birds using native shrubs [
22,
23]. Ectotherms also avoid non-native vegetation at multiple spatial scales, suggesting that non-native plants do not provide the required habitat components [
24]. Consequently, non-native plants can negatively impact native fauna by increasing mortality, reducing reproductive success, and diminishing habitat quality. Thus, understanding how and to what extent non-native plants impact native fauna requires studying multiple animal taxa.
Despite the growing body of research on the effects of non-native plants on native wildlife, temperate insectivorous bats are understudied in the invasion literature [
25]. This is problematic because North American bats rely heavily on forested landscapes and are experiencing significant population declines resulting from multiple threats (e.g., disease, wind energy production, etc.) [
26,
27]. All bat species native to eastern North America are insectivorous and require forests for roosting, reproduction, and/or foraging [
28,
29,
30]. When selecting foraging habitats in forested landscapes, bats value both the high abundance of arthropod prey as well as the appropriate vegetative structure. However, studies show that neither of these factors alone can reliably predict bat presence or activity [
31,
32]. Non-native plants further obfuscate these dynamics by influencing both arthropod abundance and vegetative structure [
5,
12]. Although most of the literature suggests that non-native vegetation negatively affects habitat structure for bats by acting as clutter that obstructs flight and impedes echolocation, non-native plants often suppress native tree and shrub seedlings and create forest structures with open midstories and canopies. Such open habitats may facilitate flight and predator avoidance [
25,
33], ultimately increasing their use by bats.
Because many bats, including all temperate North American species, echolocate [
34], passive acoustic monitoring has become a valuable tool for evaluating how bats are affected by factors such as non-native plants [
35,
36]. Bats produce echolocation pulses at a consistent rate to navigate and hunt [
37,
38]; thus, the number of pulses recorded can be a proxy for bat activity [
39]. Moreover, when a bat detects a prey item, it deviates from its consistent echolocation pattern and produces a terminal buzz, whereby the bat directs pulses toward the target prey at an increasingly rapid rate until it is found and consumed [
40]. Therefore, the number of terminal buzzes recorded can be a proxy for foraging activity [
41]. Acoustic monitoring studies have implicated low prey abundance caused by non-native plants in reduced bat activity [
42]. Acoustic monitoring has also revealed that non-native structural clutter impacts open-space foraging bats but not clutter-loving foragers [
43]. Bat guilds or species may be impacted by non-native plants differently due to variations in foraging strategy, physiology, and prey preferences, which are all often correlated [
37,
40]. Insectivorous bats can be characterized into two foraging guilds based on their echolocation pulses. Species that produce low-frequency pulses (~16–33 kHz) typically forage in open habitats, such as above the forest canopy or in openings and corridors. Such low-frequency echolocation is ideal for long-range prey detection [
44]. Oppositely, species that forage in cluttered habitats, such as in or below the forest canopy, generally produce high-frequency pulses (~34–50 kHz) that facilitate short-range object detection [
44,
45]. Thus, it is possible that non-native plants that mediate changes in overall vegetative structure, either by creating clutter or more open landscapes, may have opposite effects on bats based on their preferences in foraging habitats.
Here, we explored the relationship between non-native plants and habitat use by bats in eastern temperate forests. Using bat activity as an indicator of habitat suitability [
46,
47], we combined acoustic monitoring, conventional insect trapping, and vegetation surveys to investigate if bat activity is correlated with plot non-native percentage, which we defined as the average non-native cover of the ground and midstory vegetative layers. We predicted that survey plots with a high non-native percentage would also have open midstories and canopies, suggesting that non-native plants positively influence bat habitat by facilitating flight. Despite increased levels of overall bat activity, however, we also predicted that plots with high non-native percentages would exhibit reduced bat foraging activity relative to plots with low non-native percentages due to lower arthropod abundance and diversity. However, we hypothesized that non-native plants would affect bats differently depending on their foraging strategy: bats that prefer to forage in open areas (which produce low-frequency pulses) would likely be more active in survey plots with a high percent non-native, which we predicted would be more structurally open, while the opposite would be true for clutter-loving bats (which produce high-frequency pulses).
4. Discussion and Conclusions
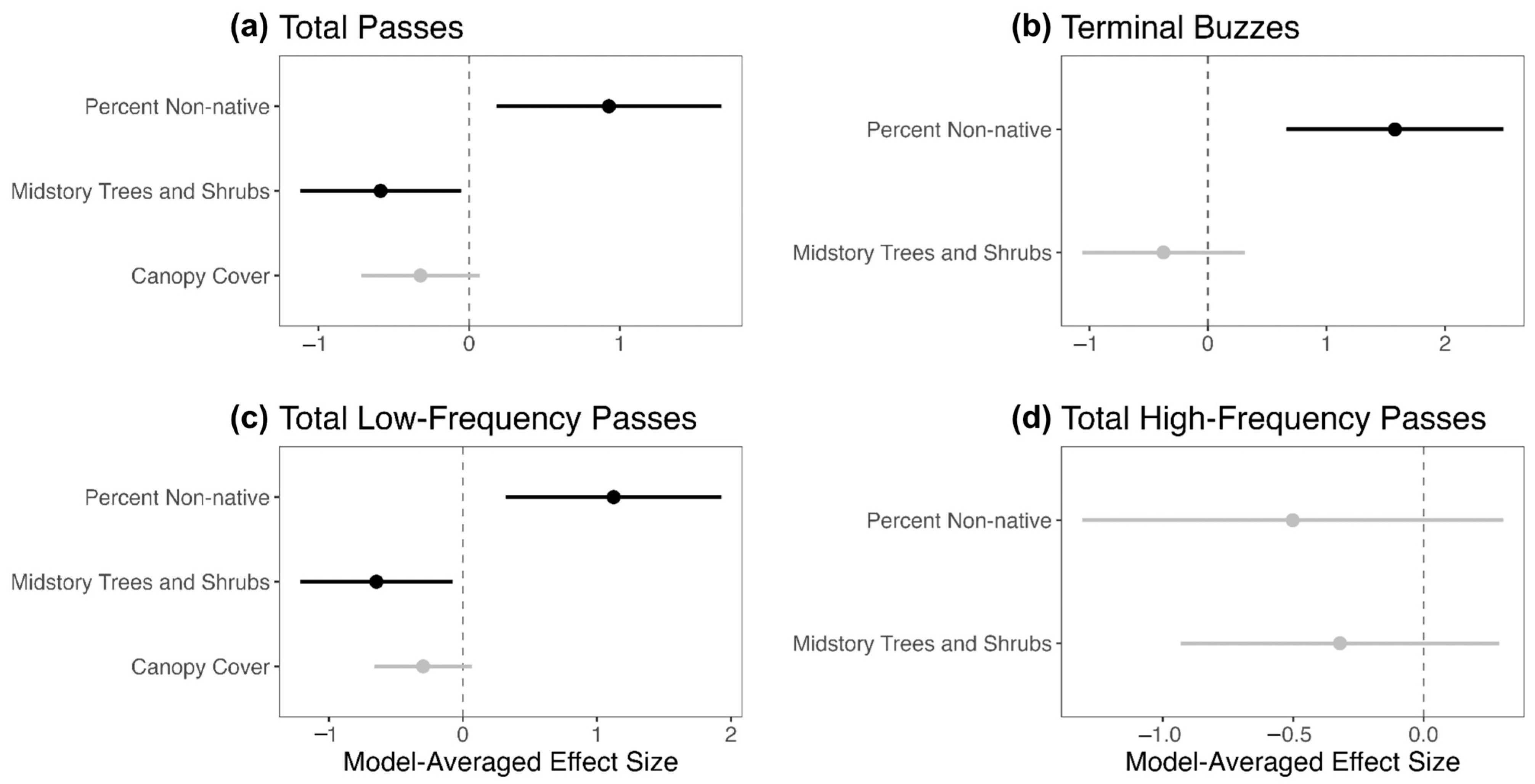
Overall, our research suggests that non-native vegetation does not have a completely negative impact on forest habitat use by bats. This conclusion is driven by two important findings. First, the percentage of non-native vegetation at a plot had the largest and most positive effect on both total passes, a proxy for overall bat activity, as well as terminal buzzes, a proxy for foraging activity, indicating that bats were more active in plots with higher percentages of non-native plants. Second, non-native percentage was not significantly correlated with putative prey abundance, which itself did not have a significant effect on overall bat activity or foraging activity. Together, such results suggest that bats value habitat structure more than prey availability when using foraging habitats and that non-native plants may benefit bats by creating habitat structures more conducive to foraging.
Contrary to our predictions and the findings of previous studies [
69,
70], putative prey abundance and total arthropod order diversity and richness were not correlated with plot percent non-native. This could suggest that either non-native plants altered arthropod species composition without changing diversity and richness, or that arthropod communities were similar amongst all plots regardless of percent non-native. The former could occur if a diverse and species-rich community of non-native arthropods replaced the pre-existing native community. It is well documented that non-native plants can be toxic to and reduce the abundance of native arthropod species [
12,
71]. Because most herbivorous insects are specialists that have coevolved with one or a few plant lineages, native arthropods are likely unequipped to combat the chemical and physical defenses of non-native plants [
72,
73]. Therefore, non-native insects, especially those that have coevolved with a specific non-native plant, may fill niches opened by the loss of native insects, leading to no net reductions in overall arthropod abundance.
Alternatively, it is possible that the introduction of non-native plants to our study area had little to no effect on the pre-existing arthropod community. Some native insects demonstrate plasticity in host plant preference and could prefer to use non-native vegetation [
74,
75]. Non-native plants can also be highly suitable for native insects and actually increase arthropod fitness [
76]. In such cases, native insect abundance could even increase post-invasion, although this was not detected in our study. In addition, many plants introduced to North America are closely related to native species (e.g., the non-native
Lonicera japonica is a congener of
L. sempervirens, L., an eastern North American native), increasing the likelihood that an herbivore that specializes in a native plant can exploit a closely related plant that produces similar chemical compounds [
73]. We did not classify arthropods into species; therefore, the mechanisms driving our results are not known.
A non-significant change in arthropod abundance, regardless of the insect community composition, logically results in few impacts on bat use of forest habitats. First, North American bats are generalist insectivores, feeding on a wide array of arthropods [
59,
77], suggesting that bats can tolerate small changes in arthropod availability. Thus, changes in insect diversity are likely inconsequential to bats, so long as there are ample numbers of prey items to make foraging activities beneficial. Furthermore, bats may not discriminate between native and non-native insects. Indeed, bats have been shown to consistently consume such non-native insects as brown marmorated stink bugs (
Halyomorpha halys, Stål) [
78], spotted lanternflies (
Lycorma delicatula, White; McHale et al., unpublished data), and emerald ash borers (
Agrilus planipennis, Waterhouse) [
79]. Thus, bats may cope with decreased abundances of native prey by consuming non-native species, which often occur in high abundances [
80,
81]. It is important to note that we caught arthropods in every survey plot, indicating that even plots with the lowest recorded arthropod abundances may have had ample prey to sustain bat foraging.
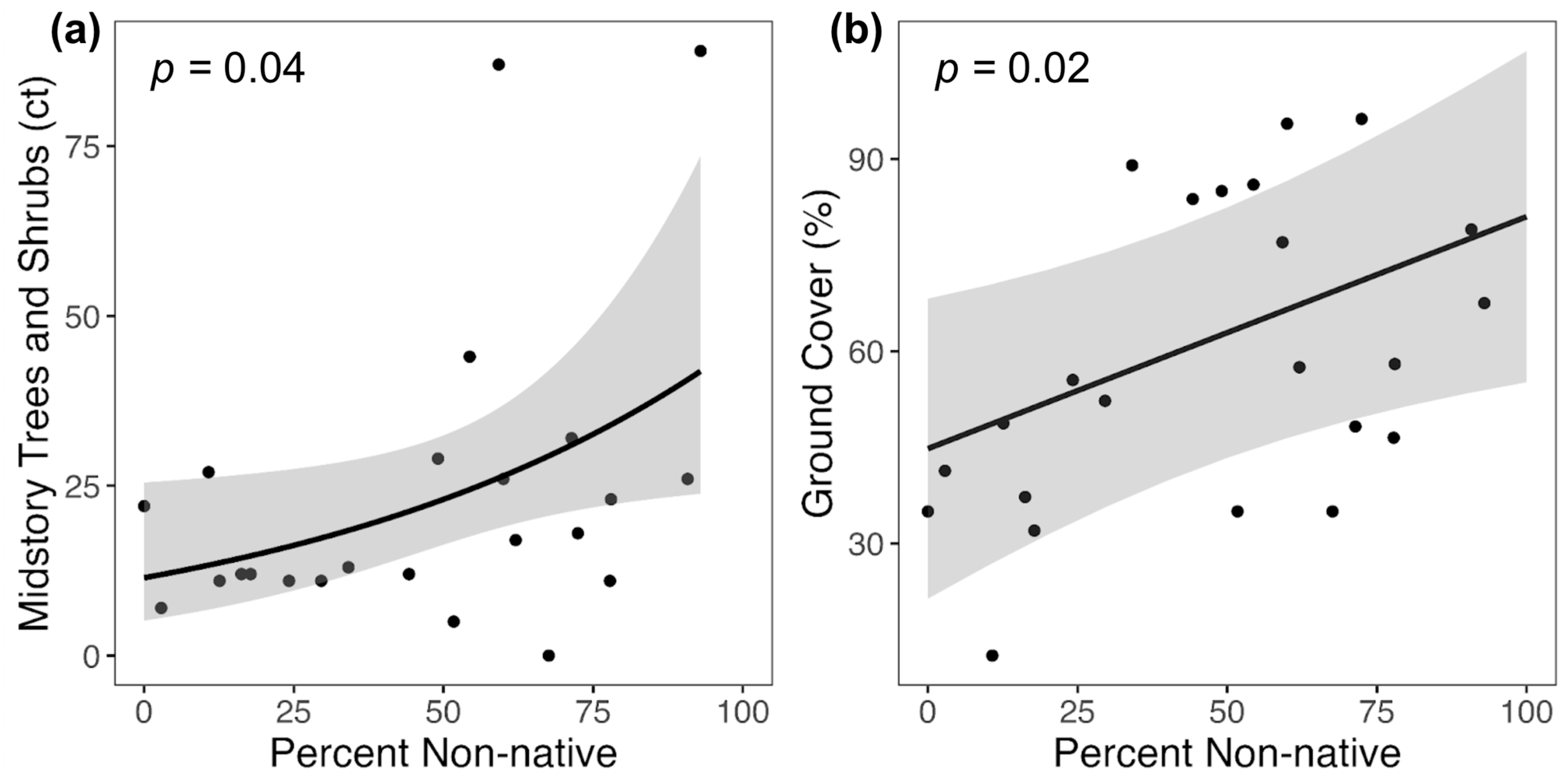
We found that the percent of non-native vegetation was also significantly and positively correlated with ground cover and the number of midstory trees and shrubs, but not correlated with canopy cover. Such findings suggest that non-native plants may proliferate in and alter the vegetative structure of the ground and midstory layers. Because North American insectivorous bats primarily use the midstory and canopy strata of forests, changes in ground-level structure likely have few direct impacts on bat activity. On the other hand, the increased number of midstory trees and shrubs, and therefore midstory clutter, associated with a higher degree of vegetative invasion, could impact bat maneuverability and foraging.
In addition to percent non-native, the number of midstory trees and shrubs also had a significant, albeit negative, effect on overall bat activity. Such results indicate that bats were more active in plots with higher percent non-native and lower midstory clutter, which is interesting given our finding that percent non-native was positively associated with midstory trees and shrubs. Logically, bats should be less active in plots with high percent non-native due to the association with increased midstory clutter. However, our model results may indicate that bats were either (1) partitioning their time between sites with only high percent non-native or only a few midstory trees and shrubs; or (2) using sites that had high percent non-native and low midstory clutter. Regarding the latter, we did survey heavily invaded plots that had fewer than 25 midstory trees and shrubs. In such plots, it is possible that non-native vegetation was concentrated at the ground layer, resulting in a high calculated plot percent non-native despite there being few, mostly native, midstory trees and shrubs. Additionally, midstory clutter did not have a significant effect on terminal buzzes, which could suggest that the habitat structure affects bat locomotion more so than foraging. While foraging bats likely compromise their own habitat preferences for the habitat preferences of their prey, commuting bats may have fewer tradeoffs to consider and more often travel down paths of least resistance [
82]. The discrepancy in the relationships between percent non-native, number of midstory trees and shrubs, and bat activity may also suggest that non-native plants may facilitate bat activity in ways that do not alter habitat structure, such as by improving habitat quality (e.g., providing roost sites). Further research should be conducted to determine the mechanisms by which non-native plants affect bats.
Although none of our measured predictors significantly affected total high-frequency passes, both percent non-native and the number of midstory trees and shrubs significantly affected total low-frequency passes, much like the model for total passes. This similarity is likely explained by the dominance within our dataset of bats that produce low-frequency pulses. The low-frequency echolocating big brown bat, responsible for over half of our recorded passes, is the most common bat in New Jersey [
83]. We attribute the model results for total low-frequency passes, which comprised less than 15% of all passes recorded, to an inadequate sample size. There are several explanations for why we recorded so few low-frequency passes. First, white-nose syndrome has significantly reduced populations of previously common species that produce high-frequency pulses, such as the little brown and northern long-eared bats [
84,
85]. Alternatively, many areas within Jockey Hollow are dominated by non-native vegetation and have low densities of midstory trees and shrubs, both of which seem to facilitate the activity of bats with low-frequency pulses. Therefore, the noticeably low number of high-frequency passes recorded is not surprising but does prevent us from concluding if non-native plants or habitat structure affect bats at Jockey Hollow differently depending on their foraging preferences.
Implications for Forest Management
Managers often recommend removing non-native plants in both forested and urban landscapes because it can significantly increase insectivorous bat activity [
43,
86,
87]. However, our results suggest that in the short term, non-native plants might not be detrimental to bats and may even benefit them by maintaining open flight space beneath the forest canopy. However, we are uncertain about how bats are impacted in the long term by non-native shrubs, especially in conjunction with deer browse. In the United Kingdom, the presence of a non-native rhododendron and red deer (
Cervus elaphus, L.) negatively affected insectivorous bats that prefer to forage in open spaces [
43]. Both non-native plants and deer browse can hinder the survival of native seedlings, which could eventually decrease plant richness and woody species recruitment [
88,
89]. As native trees die, these habitats may transition to having cluttered midstories and little to no canopy, which could negatively impact bats.
Our findings demonstrate that there are complex relationships between native and non-native species that warrant further study. Because bats seem to highly value habitat structure when choosing where to travel and forage, management activities that remove non-native plants while maintaining structural attributes preferred by bats are likely to sustain bat activity. Therefore, reducing midstory clutter and removing non-native shrubs could create travel corridors that facilitate foraging for bats with low-frequency pulses, the most common species detected at Jockey Hollow. Unfortunately, we collected insufficient data to draw conclusions about what structural habitat attributes significantly influenced high-frequency bat activity. Such an evidence base is needed to facilitate habitat-based bat conservation, particularly for bats impacted by other threats such as white-nose syndrome and wind energy infrastructure [
90,
91]. Because our model results were largely driven by big brown bats, we cannot advocate for a single management solution for increasing the activity of all bats. It is likely that a variety of management techniques that promote or maintain structurally heterogeneous habitats will support the greatest bat diversity. Further studies targeting bats with high-frequency pulses can reduce the uncertainty surrounding the impacts of non-native plants on bats of the highest conservation concern.







{kind=link}
{kind=link}
{kind=link}


