
Confocal Microscopy and Molecular Analyses Reveal Anal Secretory Apparatus in Immatures and Recover Transcontinental Clade of Gall Mites (Eriophyoidea) from Tamarisks

 , , ,
, , ,
Abstract
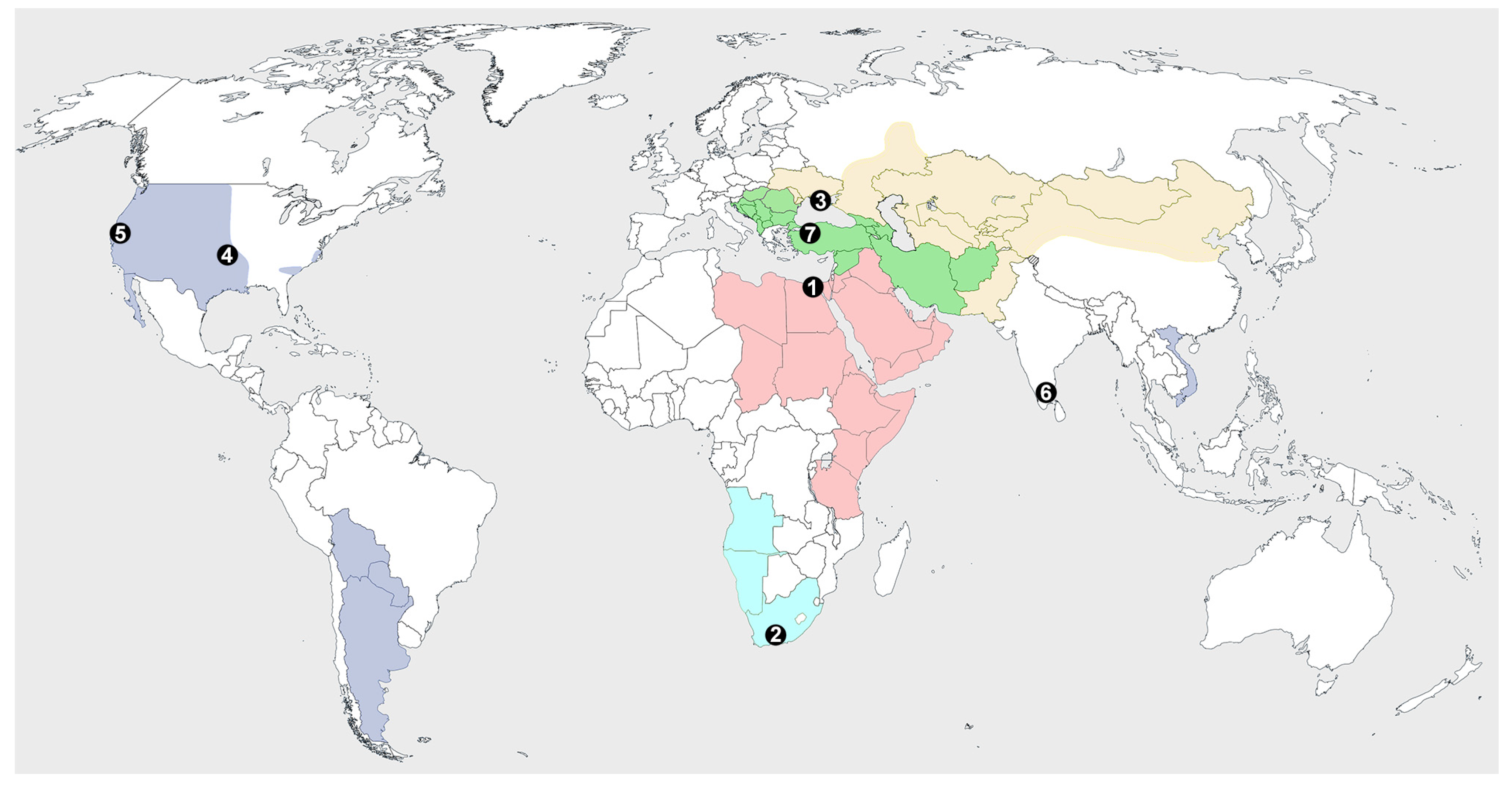
1. Introduction
2. Materials and Methods
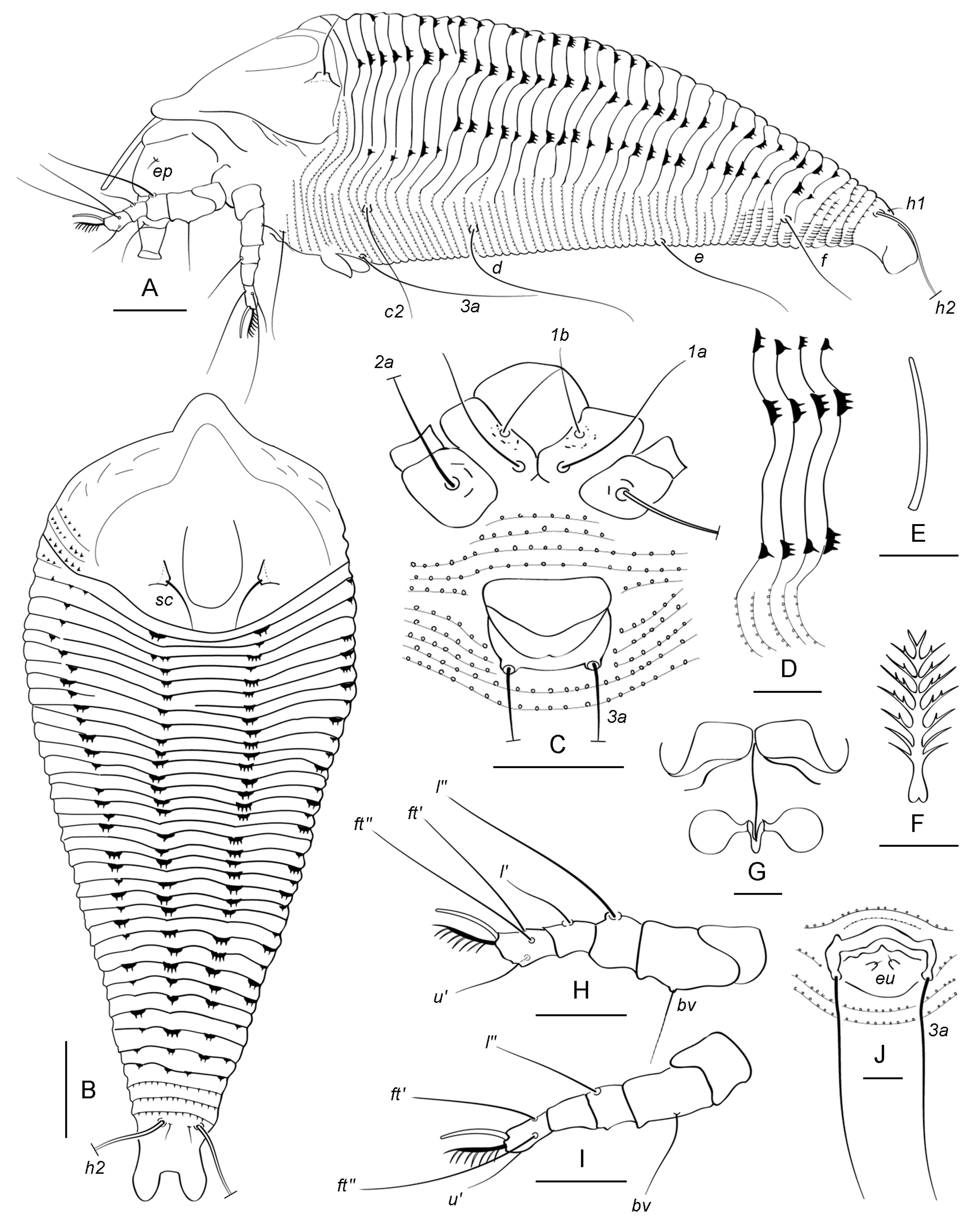
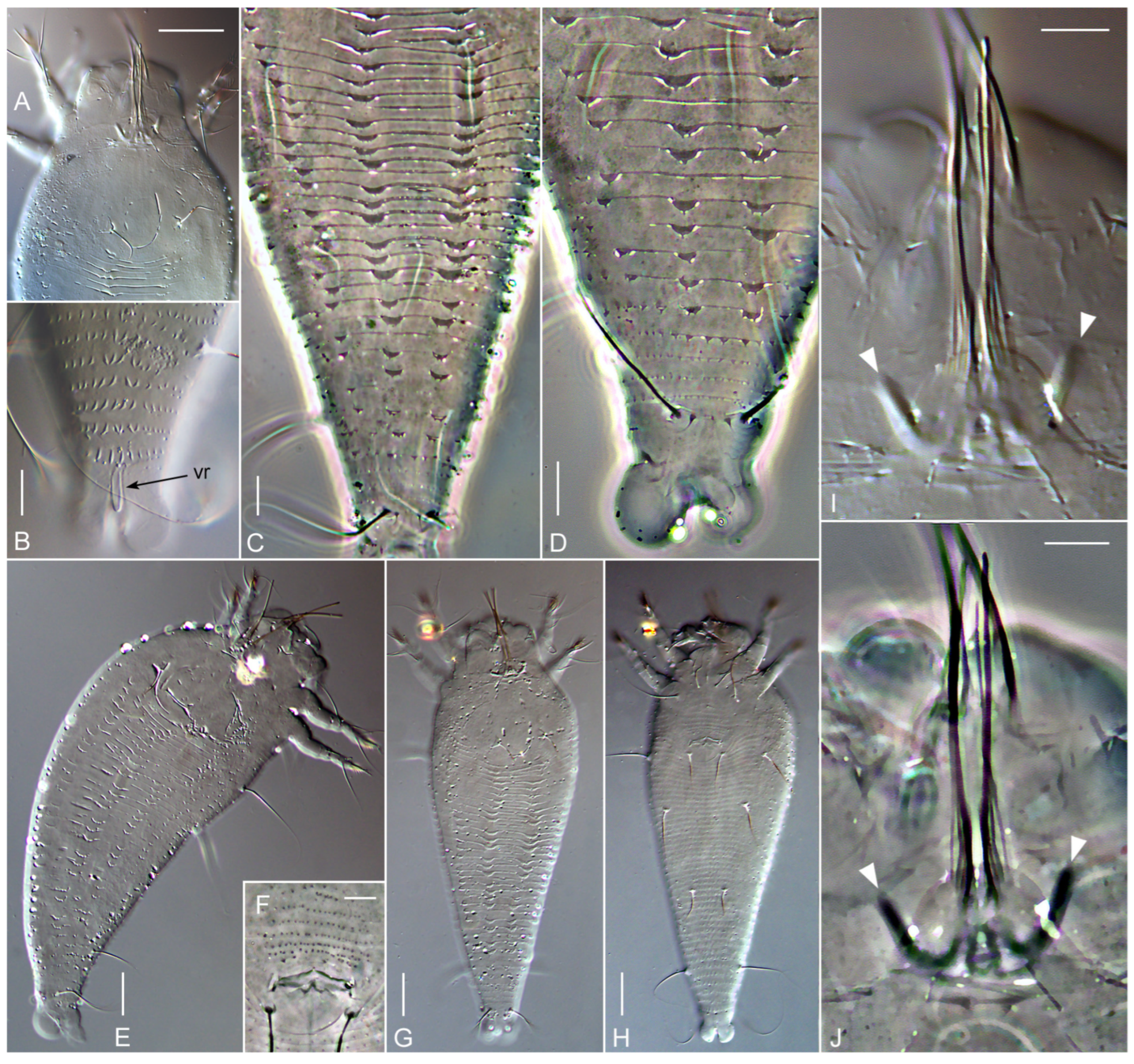
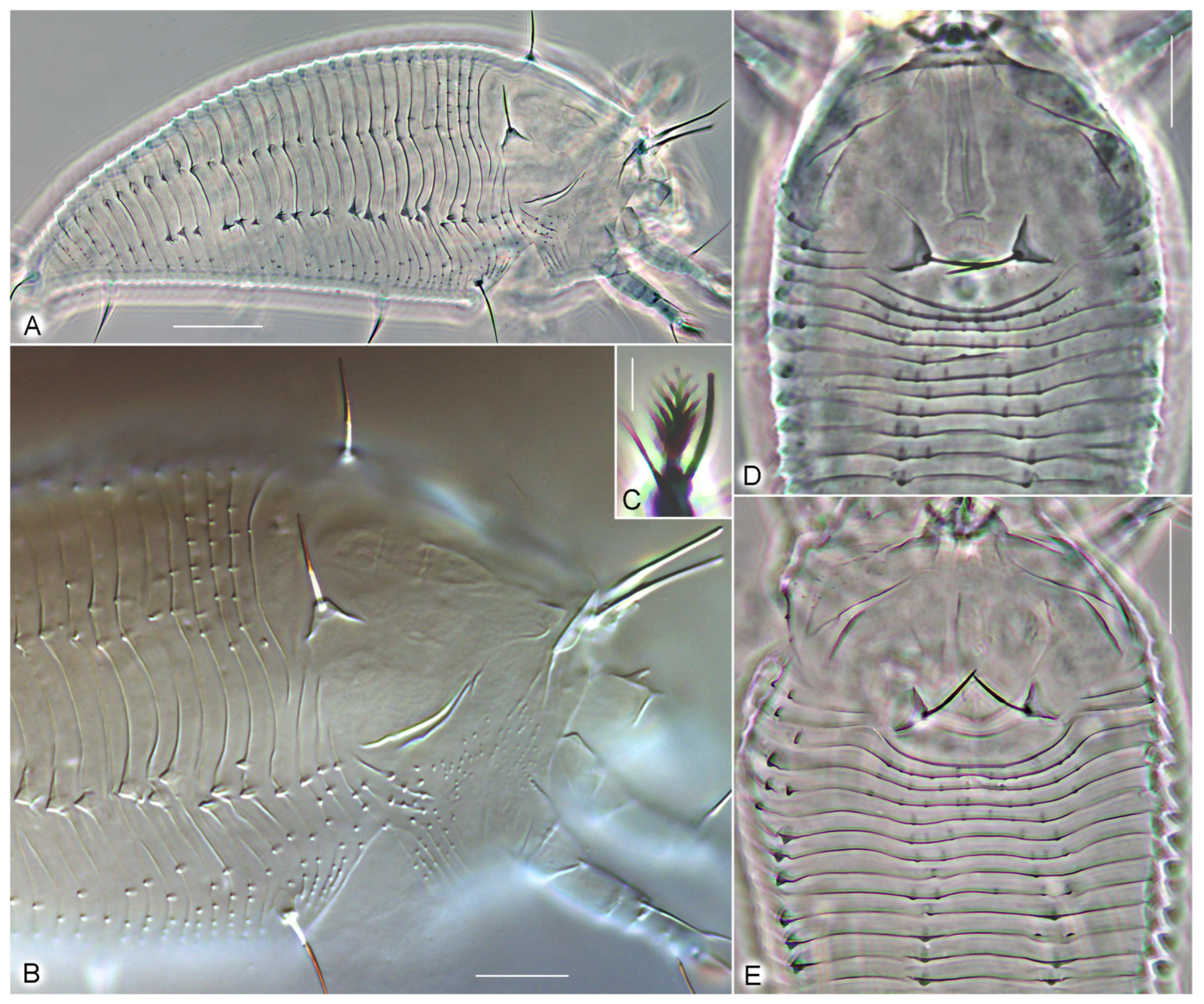
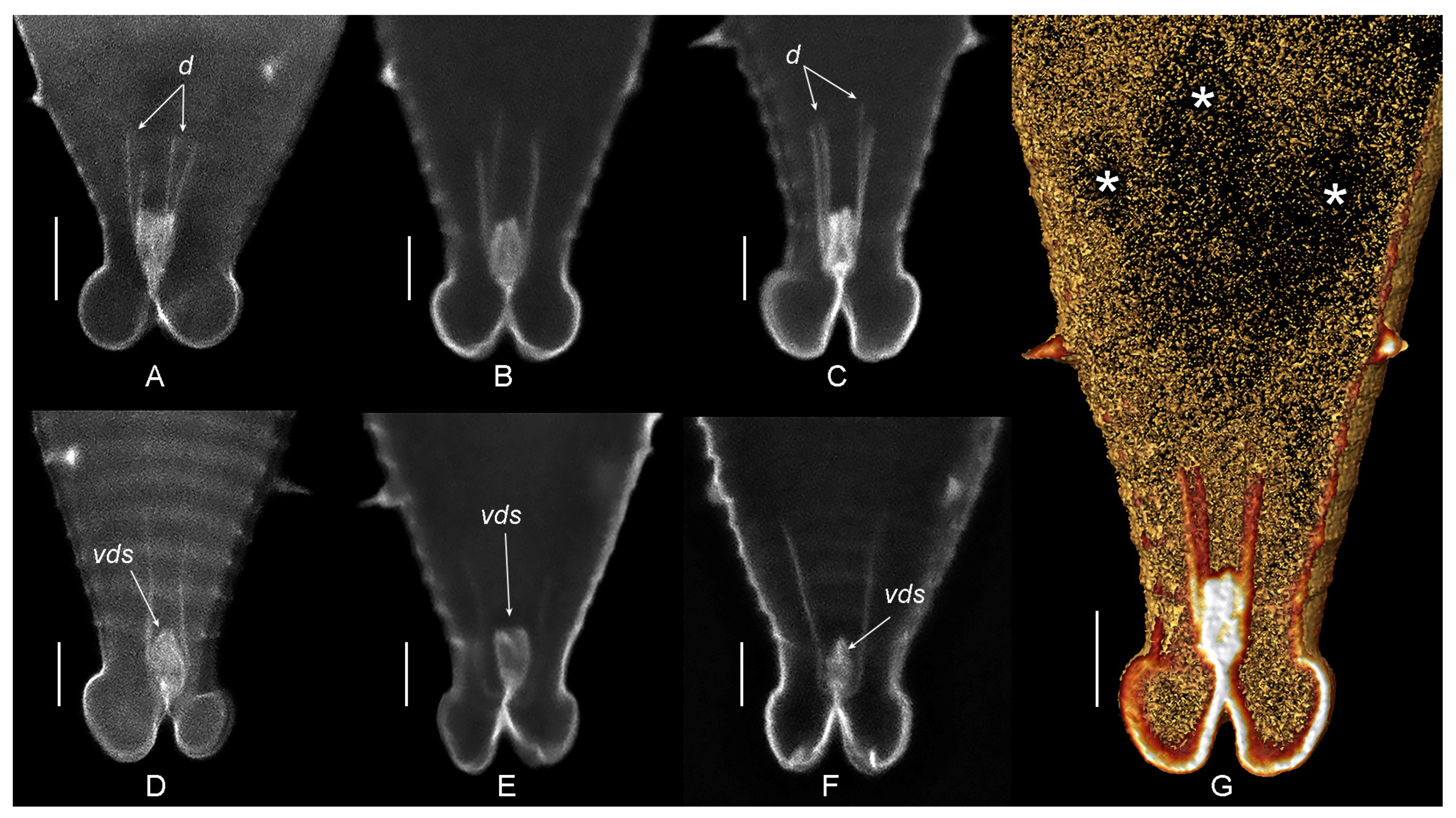
3. Results
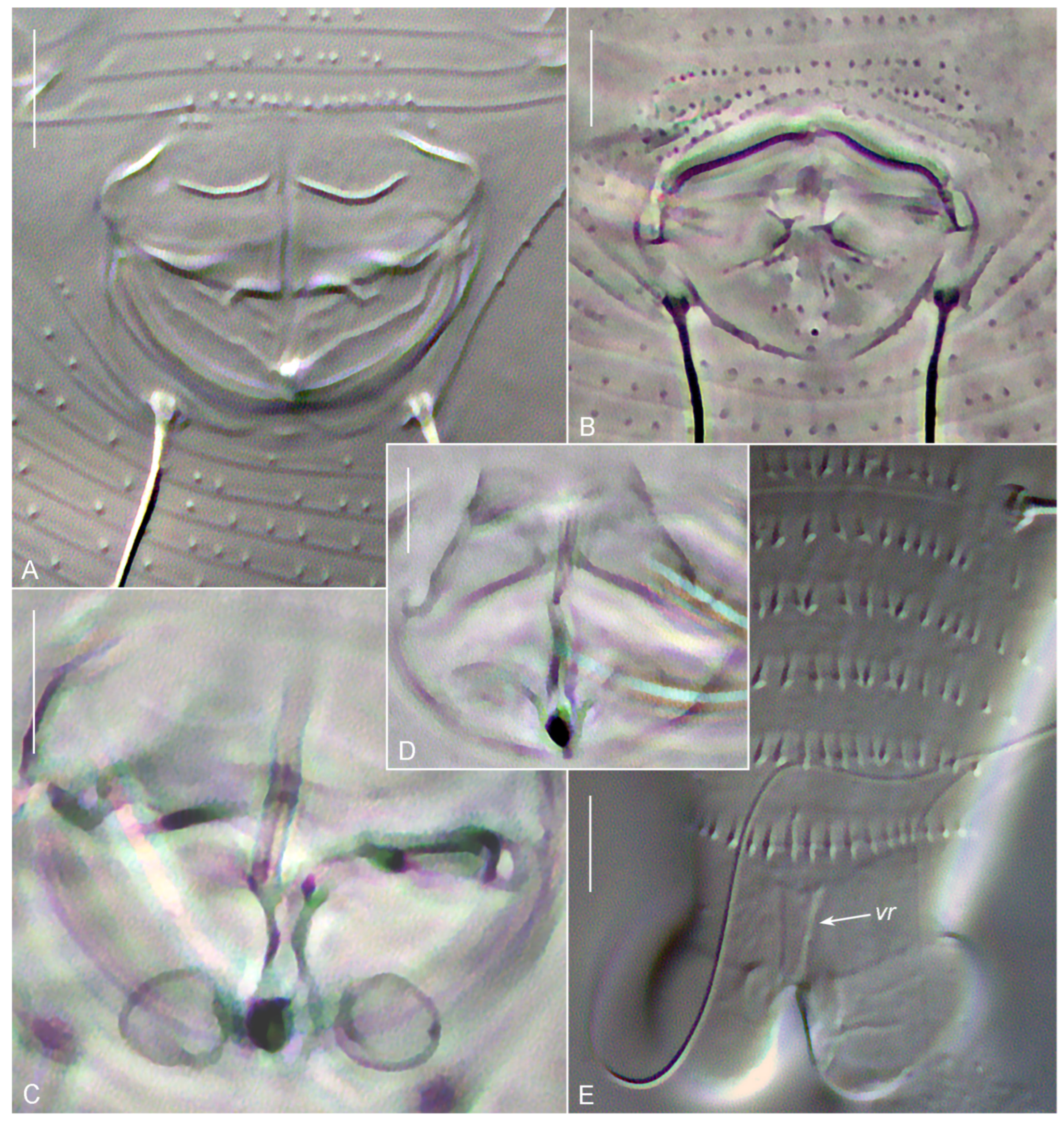
3.1. Systematics
3.2. Blast Search Results
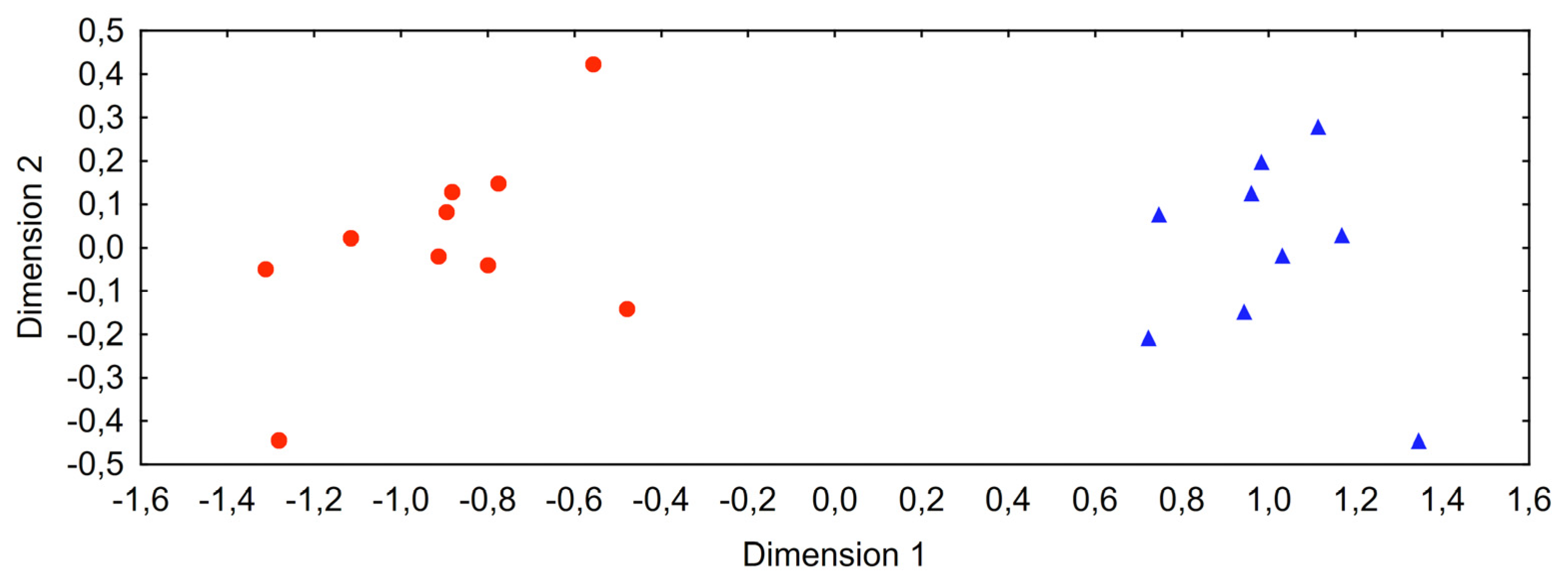
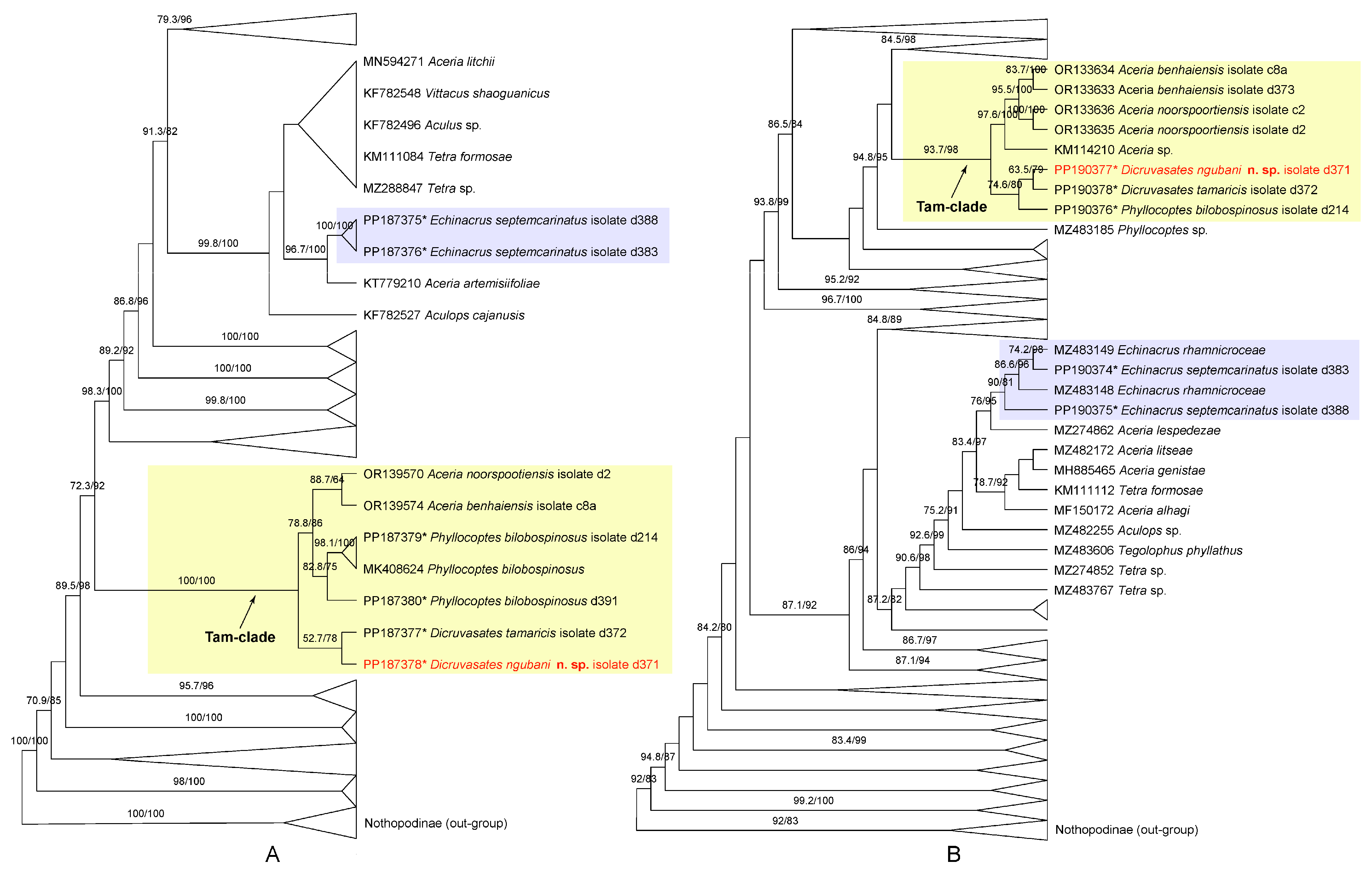
3.3. Molecular Phylogenetic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fenton, B.; Birch, A.N.E.; Malloch, G.; Lanham, P.G.; Brennan, R.M. Gall mite molecular phylogeny and its relationship to the evolution of plant host specificity. Exp. Appl. Acarol. 2000, 24, 831–861. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Klimov, P.; Letukhova, V.Y.; Ripka, G.; Zukoff, S. Two new phyllocoptine species and new records of eriophyoid mites (Eriophyidae, Phyllocoptinae) from Crimea, with evaluation of the phylogenetic position of the new taxa using GenBank data. Syst. Appl. Acarol. 2019, 24, 882–904. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Cvrković, T.; Efimov, P.G.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Schubert, M.A.; Sukhareva, S.I.; Zukoff, S.N.; Amrine, J. Molecular phylogenetic analyses reveal a deep dichotomy in the conifer-inhabiting genus Trisetacus (Eriophyoidea: Nalepellidae), with the two lineages differing in their female genital morphology and host associations. Exp. Appl. Acarol. 2020, 81, 287–316. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Craemer, C.; Cvrković, T.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Sukhareva, S.I.; Zukoff, S.N.; Bolton, S.; Amrine, J. Molecular phylogeny of the phytoparasitic mite family Phytoptidae (Acariformes: Eriophyoidea) identified the female genitalic anatomy as a major macroevolutionary factor and revealed multiple origins of gall induction. Exp. Appl. Acarol. 2021, 83, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S.; Xue, X.F.; Hong, X.Y. Homoplastic evolution and host association of Eriophyoidea (Acari, Prostigmata) conflict with the morphological-based taxonomic system. Mol. Phylogenet. Evol. 2014, 78, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.F.; Yao, L.F.; Yin, Y.; Liu, Q.; Li, N.; Hoffmann, A.A.; Sun, J.-T.; Hong, X.Y. Macroevolutionary analyses point to a key role of hosts in diversification of the highly speciose eriophyoid mite superfamily. Mol. Phylogenet. Evol. 2023, 179, 107676. [Google Scholar] [CrossRef] [PubMed]
- de Lillo, E.; Pozzebon, A.; Valenzano, D.; Duso, C. An intimate relationship between eriophyoid mites and their host plants—A review. Front. Plant Sci. 2018, 9, 418308. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.; de Lillo, E.; Amrine, J.W. Effectiveness of eriophyid mites for biological control of weedy plants and challenges for future research. Exp. Appl. Acarol. 2010, 51, 115–149. [Google Scholar] [CrossRef]
- Marini, F.; Weyl, P.; Vidović, B.; Petanović, R.; Littlefield, J.; Simoni, S.; de Lillo, E.; Cristofaro, M.; Smith, L. Eriophyid Mites in Classical Biological Control of Weeds: Progress and Challenges. Insects 2021, 12, 513. [Google Scholar] [CrossRef] [PubMed]
- de Lillo, E.; Sobhian, R. Taxonomy, distribution, and host specificity of a gall-making mite, Aceria tamaricis (Trotter) (Acari Eriophyoidea), associated with Tamarix gallica L. (Parietales: Tamaricaceae) in southern France. Entomologica 1994, 28, 5–16. [Google Scholar]
- Marlin, D.; Newete, S.W.; Mayonde, S.G.; Etienne, R.S.; Byrne, M.J. Invasive Tamarix (Tamaricaceae) in South Africa: Current research and the potential for biological control. Biol. Invasions 2017, 19, 2971–2992. [Google Scholar] [CrossRef]
- Mayonde, S.; Cron, G.V.; Glennon, K.L.; Byrne, M.J. Genetic diversity assessment of Tamarix in South Africa–Biocontrol and conservation implications. S. Afr. J. Bot. 2019, 121, 54–62. [Google Scholar] [CrossRef]
- WFO. World Flora Online. 2024. Available online: http://www.worldfloraonline.org (accessed on 24 February 2024).
- Bredenkamp, C.L. Tamaricaceae. In Plants of Southern Africa; Germishuizen, G., Meyer, N.L., Eds.; National Botanical Institute: Pretoria, South Africa, 2003; p. 927. [Google Scholar]
- Harms, R.S.; Hiebert, R.D. Vegetation response following invasive Tamarisk (Tamarix spp.) removal and implications for riparian restoration. Restor. Ecol. 2006, 14, 461–472. [Google Scholar] [CrossRef]
- Byrne, M.J.; Mayonde, S.; Venter, N.; Chidawanyika, F.; Zachariades, C.; Martin, G. Three New Biological Control Programmes for South Africa: Brazilian Pepper, Tamarix and Tradescantia. Afr. Entomol. 2021, 29, 965–982. Available online: https://hdl.handle.net/10520/ejc-ento_v29_n3_a19 (accessed on 24 February 2024). [CrossRef]
- Joshi, S.; Menon, P.; Ramamurthy, V.V. A new Eriophyid mite (Acari: Prostigmata) from India. Bioscan 2013, 8, 339–342. [Google Scholar]
- Situngu, S.; Elhalawany, A.S.; Ngubane-Ndhlovu, N.S.; Chetverikov, P.E. New species and records of gall mites of the genus Aceria (Eriophyoidea, Eriophyidae) associated with Tamarix in Egypt and South Africa. Acarologia 2023, 63, 1271–1303. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Bolton, S.J.; Gubin, A.I.; Letukhova, V.Y.; Vishnyakov, A.E.; Zukoff, S. The anal secretory apparatus of Eriophyoidea and description of Phyllocoptes bilobospinosus n. sp. (Acariformes: Eriophyidae) from Tamarix (Tamaricaceae) from Ukraine, Crimea and USA. Syst. Appl. Acarol. 2019, 24, 139–157. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Vishnyakov, A.E.; Zhuk, A.S. A New Webbing Aberoptus Species from South Africa Provides Insight in Silk Production in Gall Mites (Eriophyoidea). Diversity 2023, 15, 151. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 24 February 2024).
- Abou-Awad, B.A.; El-Borolossy, M.A. Two eriophyid mites on tamarisk trees in Egypt (Acari: Eriophyoidea: Eriophyidae). Acarologia 1995, 36, 145–148. [Google Scholar]
- Amrine, J.W.; Stasny, T.A.H.; Flechtmann, C.H.W. Revised Keys to World Genera of Eriophyoidea (Acari: Prostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2003; pp. 1–244. [Google Scholar]
- Chandrapatya, A.; Konvipasruang, P.; Amrine, J.W. Catalog of Thai Eriophyoidea (Acari: Prostigmata) with Illustrations and Keys to Genera of Thai Mites; Ku Digital Printing Center, Kasetsart University: Bangkok, Thailand, 2017; 526p. [Google Scholar]
- Elhalawany, A.S. First record of the genus Echinacrus Keifer, 1966 (Acari: Eriophyidae) on Acacia from Egypt, with description of a new species. Acarines J. Egypt. Soc. Acarol. 2013, 7, 7–12. [Google Scholar] [CrossRef]
- Amrine, J.W.; Manson, D.C.M. Preparation, mounting and descriptive study of eriophyoid mites. In World Crop Pests: Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 383–396. [Google Scholar]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–31. [Google Scholar] [CrossRef]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Stalling, D.; Westerhoff, M.; Hege, H.C. Amira: A highly interactive system for visual data analysis. Vis. Handb. 2005, 38, 749–767. [Google Scholar]
- Chetverikov, P.E.; Bertone, M.A. First rhyncaphytoptine mite (Eriophyoidea, Diptilomiopidae) parasitizing American hazelnut (Corylus americana): Molecular identification, confocal microscopy, and phylogenetic position. Exp. Appl. Acarol. 2022, 88, 75–95. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignments based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. Model Finder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Cvrković, T.; Vidović, B.; Petanović, R.U. Description of a new relict eriophyoid mite, Loboquintus subsquamatus n. gen. & n. sp. (Eriophyoidea, Phytoptidae, Pentasetacini) based on confocal microscopy, SEM, COI barcoding and novel CLSM anatomy of internal genitalia. Exp. Appl. Acarol. 2013, 61, 1–30. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Data Science Textbook. Available online: https://docs.tibco.com/data-science/textbook (accessed on 24 February 2024).
- Chetverikov, P.E. Distal oviduct and genital chamber of eriophyoids (Acariformes, Eriophyoidea): Refined terminology and remarks on CLSM technique for studying musculature of mites. Exp. Appl. Acarol. 2014, 64, 407–428. [Google Scholar] [CrossRef]
- Özdikmen, H. Nomenclatural changes for a family group name and twelve genus group names in Acari. Munus Entomlogy Zool. 2008, 3, 217–230. [Google Scholar]
- Vidović, B.; Stanisavljević, L.; Petanović, R. Phenotypic variability in five Aceria spp. (Acari: Prostigmata: Eriophyoidea) inhabiting Cirsium species (Asteraceae) in Serbia. Exp. Appl. Acarol. 2010, 52, 169–181. [Google Scholar] [CrossRef]
- Vidović, B.; Cvrković, T.; Marić, I.; Chetverikov, P.E.; Cristofaro, M.; Rector, B.G.; Petanović, R. A new Metaculus species (Acari: Eriophyoidea) on Diplotaxis tenuifolia (Brassicaceae) from Serbia: A combined description using morphology and DNA barcode data. Ann. Entomol. Soc. 2015, 108, 922–931. [Google Scholar] [CrossRef]
- Vidović, B.; Anđelković, N.; Jojić, V.; Cvrković, T.; Petanović, R.; Marini, F.; Cristofaro, M.; Rector, B.G. A New Aculodes Species (Prostigmata: Eriophyidae) Described from an Invasive Weed by Morphological, Morphometric and DNA Barcode Analyses. Insects 2022, 13, 877. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, A.; Kuczyński, L.; Rector, B.; Amrine, J.W., Jr. Wheat curl mite and dry bulb mite: Untangling a taxonomic conundrum through a multidisciplinary approach. Biol. J. Linn. Soc. 2014, 111, 421–436. [Google Scholar] [CrossRef]
- Navia, D.; Ferreira, C.B.; Reis, A.C.; Gondim, M.G. Traditional and geometric morphometrics supporting the differentiation of two new Retracrus (Phytoptidae) species associated with heliconias. Exp. Appl. Acarol. 2015, 67, 87–121. [Google Scholar] [CrossRef] [PubMed]
- Buttachon, S.; Arikit, S.; Nuchchanart, W.; Puangmalee, T.; Duanchay, T.; Jampameung, N.; Sanguansub, S. Geometric Morphometric Analysis and Molecular Identification of Coconut Mite, Aceria guerreronis Keifer (Acari: Eriophyidae) Collected from Thailand. Insects 2022, 13, 1022. [Google Scholar] [CrossRef]
- Reis, A.C.; Gondim, M.G.C., Jr.; Flechtmann, C.H.W.; Navia, D. New eriophyoid mites (Acari: Prostigmata: Eriophyoidea) from cultivated plants from northeastern Brazil, including the second taxon in the Prothricinae. J. Nat. Hist. 2014, 48, 1135–1152. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Desnitskiy, A.G.; Navia, D. Confocal microscopy refines generic concept of a problematic taxon: Rediagnosis of the genus Neoprothrix and remarks on female anatomy of eriophyoids (Acari: Eriophyoidea). Zootaxa 2015, 3919, 179–191. [Google Scholar] [CrossRef]
- Boczek, J. Studies on eriopyid mites of Poland. II. Acarologia 1961, 3, 562–570. [Google Scholar]
- Sukhareva, S.I.; (Saint-Petersburg State University, Saint-Petersburg, Russia). Personal communication, 2024.
- Chetverikov, P.E.; Craemer, C. Gnathosomal interlocking apparatus and remarks on functional morphology of frontal lobes of eriophyoid mites (Acariformes, Eriophyoidea). Exp. Appl. Acarol. 2015, 66, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.W.; Kono, T.; Amrine, J.W., Jr.; Delfinado-Baker, M.; Stasny, T.A. Eriophyoid Mites of the United States; Indira Publishing House: West Bloomfield, MI, USA, 1996; p. 394. [Google Scholar]
- Manson, D.C.M.; Gerson, U. Web spinning, wax secretion and liquid secretion by eriophyoid mites. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 251–258. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Craemer, C.; Bolton, S. Exoskeletal transformations in Eriophyoidea: New pseudotagmic taxon Pseudotagmus africanus ng & n. sp. from South Africa and remarks on pseudotagmosis in eriophyoid mites. Syst. Appl. Acarol. 2017, 22, 2093–2118. [Google Scholar] [CrossRef]
- Nuzzaci, G.; Alberti, G. Internal anatomy and physiology. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 101–150. [Google Scholar] [CrossRef]
- Propistsova, E.A.; Makarova, A.A.; Chetverikov, P.E.; Polilov, A.A. Anatomy of the miniature four-legged mite Achaetocoptes quercifolii (Arachnida: Acariformes: Eriophyoidea). Arthropod Struct. Dev. 2023, 72, 101235. [Google Scholar] [CrossRef] [PubMed]
- Pepato, A.R.; da Rocha, C.E.; Dunlop, J.A. Phylogenetic position of the acariform mites: Sensitivity to homology assessment under total evidence. BMC Evol. Biol. 2010, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Bolton, S.J.; Chetverikov, P.E.; Ochoa, R.; Klimov, P.B. Where Eriophyoidea (Acariformes) Belong in the Tree of Life. Insects 2023, 14, 527. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Craemer, C.; Neser, S.; Peralta, L.; Amrine, J.W. Transcontinental dispersal, common ancestry or convergent evolution? New phyllocoptines (Eriophyidae) from American and South African relict conifers. Syst. Appl. Acarol. 2017, 22, 724–748. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Zhuk, A.S. Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny and Mitogenomics. Insects 2023, 14, 507. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sun, J.-T.; Yin, Y.; Hong, X.-Y.; Xue, X.-F. Global patterns, and drivers of herbivorous eriophyoid mite species diversity. J. Biogeogr. 2023, 50, 330–340. [Google Scholar] [CrossRef]
- Navia, D.; Ochoa, R.; Welbourn, C.; Ferragut, F. Adventive eriophyoid mites: A global review of their impact, pathways, prevention and challenges. Exp. Appl. Acarol. 2013, 51, 225–255. [Google Scholar] [CrossRef] [PubMed]
- Navajas, M.; Ochoa, R. Integrating ecology and genetics to address Acari invasions. Exp. Appl. Acarol. 2013, 59, 1–10. [Google Scholar] [CrossRef][Green Version]
- Knorr, L.C.; Phatak, H.C.; Keifer, H.H. Web-spinning eriophyid mites. J. Wash. Acad. Sci. 1976, 66, 228–234. [Google Scholar]
- Michalska, K.; Skoracka, A.; Navia, D.; Amrine, J.W. Behavioural studies on eriophyoid mites: An overview. Exp. Appl. Acarol. 2010, 51, 31–59. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mite Species | Collecting Data | DNA Isolate | Cox1 | 28S D1D2 | ITS1-5.8S-ITS2 |
|---|---|---|---|---|---|
| Dicruvasates ngubani n. sp. | SOUTH AFRICA: Noorspoort farm, Steytlerville, Eastern Cape (33°17′44″ S, 24°21′34″ E), vagrant on bud scales and twigs of Tamarix usneoides E.Mey. ex Bunge (Tamaricaceae), 19 October 2022, coll. N.P. Ngubane-Ndhlovu and S. Situngu. | d371 | PP190377 | PP187378 | PP187385 |
| SOUTH AFRICA: near Dwayka River, Western Cape (33°05′09″ S, 21°34′40″ E), vagrant on bud scales and twigs of Tamarix usneoides E.Mey. ex Bunge (Tamaricaceae), 19 October 2022, coll. N.P Ngubane-Ndhlovu and S. Situngu. | d9 | – | – | PP187384 | |
| c11 | – | – | PP187382 | ||
| c12 | – | – | PP187383 | ||
| Dicruvasates tamaricis Abou-Awad and El-borolossy, 1995 | EGYPT: Qalyubia governorate, 30°17′20.02″ N, 31°14′51.85″ E, on twigs of Tamarix nilotica (Ehrenb.) Bunge (Tamaricaceae), 9 March 2021, coll. A. Elhalawany. | d372 | PP190378 | PP187377 | PP187381 |
| Echinacrus septemcarinatus (Liro, 1941) | RUSSIA: Krasnodar Krai, Adler City District, near Krasnaya Polyana, right bank of river Mzymta, 43°39′21.9″ N, 40°09′23.0″ E, a vagrant on the lower leaf surface of Frangula alnus Mill. (Rhamnaceae), 10 July 2021, coll. P.E.Chetverikov. | d388 | PP190375 | PP187375 | PP187388 |
| RUSSIA: Leningrad Oblast, Gatchina District, vil. Vyritza, 59°23′43.3″ N, 30°18′00.1″ E, a vagrant on the lower leaf surface of Frangula alnus Mill. (Rhamnaceae), 25 September 2017, coll. P.E.Chetverikov. | d383 | PP190374 | PP187376 | PP187387 | |
| Phyllocoptes bilobospinosus Chetverikov et al., 2019 | USA: Kansas, Garden City, right bank of Arkansas river, 37°57′24.9″ N 100°53′57.3″ W, on young shoots of Tamarix ramosissima Ledeb. (Tamaricaceae), 12 July 2018, coll. S. Zukoff. | d391 | – | PP187380 | PP187386 |
| RUSSIA: Crimea, Alushta Municipality, vil. Pryvetnoye, Kanaka Nature Reserve, roadside near the coast of Black Sea, 44°47′13′ N′, 34°39′01′ E′, on young shoots of Tamarix tetrandra Pall. ex Bieb. (Tamaricaceae), 3 June 2018, coll. P.E.Chetverikov. | d214 | PP190376 | PP187379 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chetverikov, P.E.; Ngubane-Ndhlovu, N.P.; Situngu, S.; Elhalawany, A.; Amrine, J. Confocal Microscopy and Molecular Analyses Reveal Anal Secretory Apparatus in Immatures and Recover Transcontinental Clade of Gall Mites (Eriophyoidea) from Tamarisks. Forests 2024, 15, 715. https://doi.org/10.3390/f15040715
Chetverikov PE, Ngubane-Ndhlovu NP, Situngu S, Elhalawany A, Amrine J. Confocal Microscopy and Molecular Analyses Reveal Anal Secretory Apparatus in Immatures and Recover Transcontinental Clade of Gall Mites (Eriophyoidea) from Tamarisks. Forests. 2024; 15(4):715. https://doi.org/10.3390/f15040715
Chicago/Turabian StyleChetverikov, Philipp E., Nompumelelo P. Ngubane-Ndhlovu, Sivu Situngu, Ashraf Elhalawany, and James Amrine. 2024. "Confocal Microscopy and Molecular Analyses Reveal Anal Secretory Apparatus in Immatures and Recover Transcontinental Clade of Gall Mites (Eriophyoidea) from Tamarisks" Forests 15, no. 4: 715. https://doi.org/10.3390/f15040715
APA StyleChetverikov, P. E., Ngubane-Ndhlovu, N. P., Situngu, S., Elhalawany, A., & Amrine, J. (2024). Confocal Microscopy and Molecular Analyses Reveal Anal Secretory Apparatus in Immatures and Recover Transcontinental Clade of Gall Mites (Eriophyoidea) from Tamarisks. Forests, 15(4), 715. https://doi.org/10.3390/f15040715