

Alien vs. Native—Influence of Fallow Deer (Dama dama) Introduction on the Native Roe Deer (Capreolus capreolus) Population


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. Estimation of Fallow Deer Population Development after Its Introduction to the Głuchów Forest
2.2.2. Estimation of Roe Deer Density Prior to and after Fallow Deer Introduction
2.2.3. Estimation of Roe Deer Productivity Prior to and after Fallow Deer Introduction
2.2.4. Spatial Relationship between Roe Deer and Fallow Deer
2.2.5. Hunting Bag and Financial Data
2.3. Statistical Analysis
3. Results
3.1. Fallow Deer Population Development in Głuchów Forest
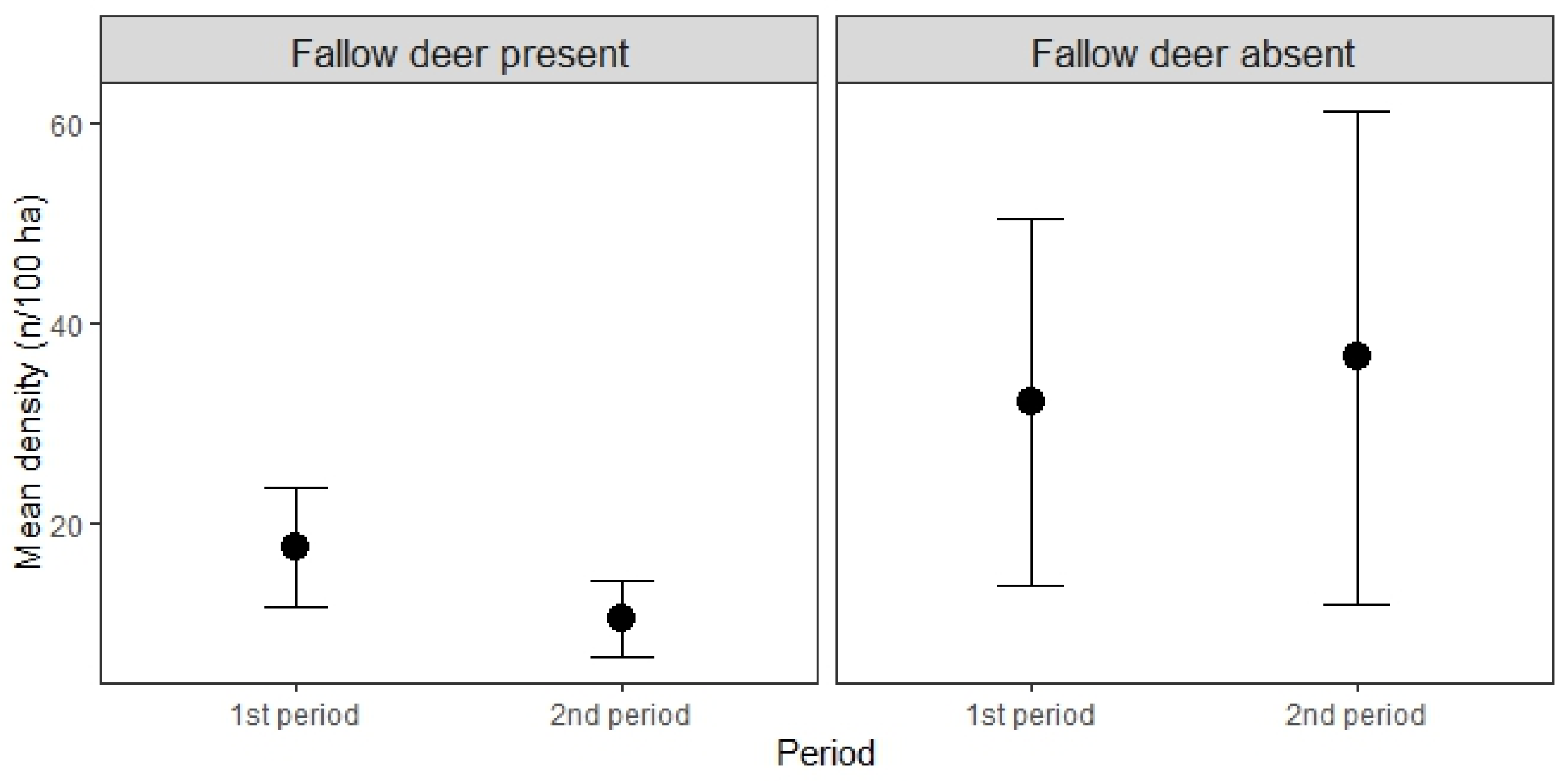
3.2. Changes in Roe Deer Density after Fallow Deer Introduction
3.3. Roe Deer Population Productivity before and after Fallow Deer Introduction
3.4. Spatial Relationship between Roe Deer and Fallow Deer
3.5. Economical Effect of Fallow Deer Introduction
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cause of Death | n of Cases |
|---|---|
| Harvested by hunters | 39 |
| Killed by stray dogs | 6 |
| Died due to starvation * | 3 |
| Trapped by a baller twine ** | 2 |
| Killed by car | 2 |
| Drowned | 1 |
| Poached | 1 |
| Unidentified reasons | 16 |
Appendix B. Behavioral Interactions between Fallow and Roe Deer
- Direct attack of a fallow deer male on roe deer; roe deer is chased away.
- Roe deer interrupts its previous activity and moves away when fallow deer approaches.
- Roe deer waiting in the vicinity of a feeding place, young forest plantation, or margins of forest meadow until the fallow deer is gone.
Appendix C. Correlation between Roe Deer Density and Hunting Bag in Two Research Areas

References
- Craik, C. Long-term effects of North American Mink Mustela vison on seabirds in western Scotland. Bird Study 1997, 44, 303–309. [Google Scholar] [CrossRef]
- Simberloff, D. Eradication of island invasives: Practical actions and results achieved. Trends Ecol. Evol. 2001, 16, 273–274. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Wauters, L.A.; Lurz, P.W.W.; Santicchia, F.; Romeo, C.; Ferrari, N.; Martinoli, A.; Gurnell, J. Interactions between native and invasive species: A systematic review of the red squirrel-gray squirrel paradigm. Front. Ecol. Evol. 2023, 11, 1083008. [Google Scholar] [CrossRef]
- Spear, D.; Chown, S. Non-indigenous ungulates as a threat to biodiversity. J. Zool. 2009, 279, 1–17. [Google Scholar] [CrossRef]
- Volery, L.; Jatavallabhula, D.; Scillitani, L.; Bertolino, S.; Bacher, S. Ranking alien species based on their risks of causing environmental impacts: A global assessment of alien ungulates. Glob. Chang. Biol. 2021, 27, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Genovesi, P.; Bacher, S.; Kobelt, M.; Pascal, M.; Scalera, R. Alien Mammals of Europe. In Handbook of Alien Species in Europe, Invading Nature; Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2009; Volume 3, pp. 119–128. [Google Scholar]
- Chapman, D.; Chapman, N. Fallow Deer. Their History, Distribution and Biology; Terence Dalton Ltd.: Lavenham, Suffolk, UK, 1975. [Google Scholar]
- Dzięciołowski, R. Daniel; Warsaw University of Life Science: Warsaw, Poland, 1994. [Google Scholar]
- Masseti, M.; Cavallaro, A.; Pecchioli, E.; Vernesi, C. Artificial occurrence of the fallow deer, Dama dama dama (L, 1758), on the island of Rhodes (Greece): Insight from mtDNA Analysis. Hum. Evol. 2005, 21, 167–175. [Google Scholar] [CrossRef]
- Heidemann, G. Damwild, Cervus dama Linné, 1758, in Kleinasien. Bestand un Schutz. Säugetierkundliche Mitteilungen 1976, 24, 124–132. [Google Scholar]
- Marinis, A.D.; Masseti, M. Mammalian fauna of the Temessos National Park, Turkey. ZooKeys 2009, 31, 221–228. [Google Scholar] [CrossRef]
- Baker, K.H.; Miller, H.; Doherty, S.; Gray, H.W.I.; Daujat, J.; Çakırlar, C.; Spassov, N.; Trantalidou, K.; Madgwick, R.; Lamb, A.L.; et al. The 10,000-year biocultural history of fallow deer and its implications for conservation policy. Proc. Natl. Acad. Sci. USA 2024, 121, e2310051121. [Google Scholar] [CrossRef]
- Chapman, N.G.; Chapman, D.I. The distribution of fallow deer: A worldwide review. Mamm. Rev. 1980, 10, 61–138. [Google Scholar] [CrossRef]
- Esattore, B.; Saggiomo, L.; Sensi, M.; Vanessa, F.; Cherin, M. Tell me what you eat and I’ll tell you…where you live: An updated review of the worldwide distribution and foraging ecology of the fallow deer (Dama dama). Mamm. Biol. 2022, 102, 321–338. [Google Scholar] [CrossRef]
- Chankaya, C.; Dokora, A.; Muchenje, V.; Hoffman, L. The fallow deer (Dama spp); endangered of not? Zool. Gart 2016, 85, 160–172. [Google Scholar] [CrossRef]
- Apollonio, M. Dama dama (Linnaeus, 1758). In The Atlas of European Mammals; Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P.J.H., Spitzenberger, F., Stubbe, M., Thissen, J.B.M., Vohralík, V., Zima, J., Eds.; T & AD Poyser Natural History: London, UK, 1999; pp. 1–484. [Google Scholar]
- Bijl, H.; Csányi, S. Fallow Deer (Dama dama). Population and Harvest Changes in Europe since the Early 1980s. Sustainability 2022, 14, 12198. [Google Scholar] [CrossRef]
- Daszkiewicz, T.; Hnatyk, N.; Dąbrowski, D.; Janiszewski, P.; Gugolek, A.; Kubiak, D.; Śmiecińska, K.; Winarski, R.; Koba-Kowalczyk, M. A comparison of the quality of the Longisissimus lumborum muscle from wild and farm raised fallow deer (Dama dama L). Small Rumin. Res. 2015, 129, 77–83. [Google Scholar] [CrossRef]
- Solarz, W. Dama dama (Linnaeus, 1758). In Gatunki Obce w Faunie Polski; Głowaciński, Z., Okarma, H., Pawłowski, J., Solarz, W., Eds.; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2012. [Google Scholar]
- Kamieniarz, R.; Rzońca, Z. Metody oraz skala introdukcji, reintrodukcji i wsiedlania zwierzyny w Polsce. In Łowiectwo w Zrównoważonej Gospodarce Łowieckiej, Zimowa Szkoła Leśna; Gil, W., Ed.; Instytut Badawczy Leśnictwa: Sękocin Stary, Poland, 2015; pp. 91–103. [Google Scholar]
- Statistical Yearbook of Forestry; Central Statistical Office: Warsaw, Poland, 2022.
- Kamieniarz, R.; Jakubowski, M.; Dyderski, M.K.; Górecki, G.; Nasiadka, P.; Okarma, H.; Pudełko, M.; Skubis, J.; Tomek, A.; Wajdzik, M.; et al. Application of the tyraliera counting method to the large-scale inventory of red deer Cervus elaphus in the northern part of Western Pomerania, Poland. Ann. For. Res. 2023, 66, 33–34. [Google Scholar] [CrossRef]
- Jamrozy, G. Carnivores, even-toed ungulates, lagomorphs and large rodents in Polish national parks. Ann. Zool. Fenn. 2008, 45, 299–307. [Google Scholar] [CrossRef]
- Jamrozy, G. (Ed.) Ssaki Polskich Parków Narodowych: Drapieżne, Kopytne, Zajęczaki i duże Gryzonie; University of Agriculture: Kraków, Poland, 2014. [Google Scholar]
- Olszewski, A. Using camera-trapping as a non-invasive technique the study of wildlife in Kampinos National Park—The first results. Stud. Mater. CEPL 2016, 49, 42–49. [Google Scholar]
- Solarz, W. Fallow Deer. In Atlas of Polish Mammals. Available online: https://wwwiopkrakowpl/Ssaki/gatunek/124 (accessed on 13 May 2024).
- Pucek, Z.; Raczyński, J. (Eds.) Atlas Rozmieszczenia Ssaków w Polsce; PWN: Warsaw, Poland, 1983. [Google Scholar]
- Bombik, E.; Kondracki, S. Znaczenie hodowli zwierząt wolno żyjących (cz II). Przegl. Hod. 2007, 75, 18–21. [Google Scholar]
- Borys, B.; Bogdaszewska, Z.; Bogdaszewski, M. Dynamiczny wzrost fermowej hodowli jeleni i danieli w Polsce. Wiad. Zootech. 2012, 1, 33–44. [Google Scholar]
- European Union. Regulation (EU) no 1143/2014 of the European parliament and of the Council of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species. Off. J. Eur. Union 2014, L317, 35–55. [Google Scholar]
- Langbein, J.; Thirgood, S.J. Variation in Mating Systems of Fallow Deer (Dama dama) in Relation to Ecology. Ethology 1989, 83, 195–214. [Google Scholar] [CrossRef]
- Obidziński, A.; Kiełtyk, P.; Borkowski, J.; Bolibok, L.; Remuszko, K. Autumn-winter diet overlap of fallow, red, and roe deer in forest ecosystems, Southern Poland. Cent. Eur. J. Biol. 2013, 8, 8–17. [Google Scholar] [CrossRef]
- Ferretti, F.; Fattorini, N. Competitor densities, habitat, and weather: Effects on interspecific interactions between wild deer species. Integr. Zool. 2020, 16, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, F.; Bertoldi, G.; Sforzi, A.; Fattori, L. Roe and fallow deer: Are they compatible neighbours? Eur. J. Wildl. Res. 2011, 57, 775–783. [Google Scholar] [CrossRef]
- Focardi, S.; Aragno, P.; Montanaro, P.; Riga, F. Inter-specific competition from fallow deer Dama dama reduces habitat quality for the Italian roe deer Capreolus capreolus italicus. Ecography 2006, 29, 407–417. [Google Scholar] [CrossRef]
- Bartoš, L.; Vaňková, D.; Šiler, J.; Losos, S. Fallow deer tactic to compete over food with red deer. Aggress. Behav. 1996, 22, 375–385. [Google Scholar] [CrossRef]
- Dróżdż, J. Studies on helminths and helminthiases in Cervidae. III. Historical formation of helminthofauna in Cervidae. Acta Parasitol. Pol. 1967, 14, 287–300. [Google Scholar]
- Moriarty, A. The liberation, distribution, abundance and management of wild deer in Australia. Wildl. Res. 2004, 31, 291–299. [Google Scholar] [CrossRef]
- Dolman, P.M.; Wäber, K. Ecosystem and competition impacts of introduced deer. Wildl. Res. 2008, 35, 202–214. [Google Scholar] [CrossRef]
- Novillo, A.; Ojeda, R.A. The exotic mammals of Argentina. Biol. Invasions 2008, 10, 1333–1344. [Google Scholar] [CrossRef]
- Relva, M.A.; Nuñez, M.A.; Simberloff, D. Introduced deer reduce native plant cover and facilitate invasion of non-native tree species: Evidence for invasional meltdown. Biol. Invasions 2010, 12, 303–311. [Google Scholar] [CrossRef]
- Martin, T.G.; Arcese, P.; Scheeder, N. Browsing down our natural heritage: Deer impacts on vegetation structure and songbird populations across an island archipelago. Biol. Conserv. 2011, 144, 459–469. [Google Scholar] [CrossRef]
- Tanentzap, A.J.; Kirby, K.J.; Goldberg, E. Slow responses of ecosystems to reductions of deer (Cervidae) populations and strategies for achieving recovery. For. Ecol. Manag. 2012, 264, 159–166. [Google Scholar] [CrossRef]
- Wright, D.M.; Andrew, J.; Tanentzap, A.J.; Flores, O.; Husheer, S.W.; Duncan, R.P.; Wiser, S.K.; Coomes, D.A. Impacts of culling and exclusion of browsers on vegetation recovery across New Zealand forests. Biol. Conserv. 2012, 153, 64–71. [Google Scholar] [CrossRef]
- Borkowski, J.; Palmer, S.; Borowski, Z. Drive counts as a method of estimating ungulate density in forests: Mission impossible? Acta Theriol. 2011, 56, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Marcon, A.; Battocchio, D.; Apollonio, M.; Grignolio, S. Assessing precision and requirements of three methods to estimate roe deer density. PLoS ONE 2019, 14, e0222349. [Google Scholar] [CrossRef] [PubMed]
- Gryz, J.; Krauze-Gryz, D. Why Did Brown Hare Lepus europaeus Disappear from Some Areas in Central Poland? Diversity 2022, 14, 465. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D. Occurrence of moose Alces alces L in the Rogów Forest (central Poland). For. Res. Pap. 2022, 82, 179–186. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Gryz, J. Den dwelling carnivores in central Poland: Long term trends in abundance and productivity. Diversity 2022, 15, 32. [Google Scholar] [CrossRef]
- Mayle, B.A.; Peace, A.J.; Gill, M.A. How Many Deer? A Field Guide to Estimating Deer Population Size; Forestry Commission: Dry Branch, GA, USA, 1999; Volume 18. [Google Scholar]
- Borkowski, J.; Banul, R.; Jurkiewicz-Azab, J.; Hołdyński, C.; Święczkowska, J.; Nasiadko, M.; Dariusz Załuski, D. There is only one winner: The negative impact of red deer density on roe deer numbers and distribution in the Słowiński National Park and its vicinity. Ecol. Evol. 2021, 11, 6889–6899. [Google Scholar] [CrossRef] [PubMed]
- Aulak, W.; Babińska-Werka, J. Estimation of roe deer density based on the abundance and rate of disappearance of their faeces from forest. Acta Theriol. 1990, 35, 111–120. [Google Scholar] [CrossRef]
- Spitzer, R.; Churski, M.; Felton, A.; Heurich, M.; Kuijper, D.P.J.; Landman, M.; Rodriguez, E.; Singh, N.J.; Taberlet, P.; van Beeck Calkoen, S.; et al. Doubting dung: eDNA reveals high rates of misidentification in diverse European ungulate communities. Eur. J. Wildl. Res. 2019, 65, 28. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D.; Klich, D. Alien or Native? How to Distinguish Feces of Fallow and Roe Deer Using Central Poland as a Case Study. Animals 2022, 12, 290. [Google Scholar] [CrossRef] [PubMed]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-Normal Plots and Overdispersed Models in R: The hnp Package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://wwwR-projectorg (accessed on 10 September 2023).
- Forest Data Bank. Hunting Reports. Available online: https://wwwbdllasygovpl/portal/ (accessed on 8 September 2023).
- Hofmann, R.R. Digestive physiology of the deer—Their morphophysiological specialisation and adaptation. R. Soc. N. Z. Bull. 1985, 22, 393–407. [Google Scholar]
- Latham, J. Interspecific interactions of ungulates in European forests: An overview. For. Ecol. Manag. 1999, 120, 13–21. [Google Scholar] [CrossRef]
- Latham, J.; Staines, B.W.; Gorman, M.L. Correlations of red (Cervus elaphus) and roe (Capreolus capreolus) deer densities in Scottish forests with environmental variables. J. Zool. 2009, 242, 681–704. [Google Scholar] [CrossRef]
- Ferretti, F.; Sforzi, A.; Lovari, S. Behavioural interference between ungulate species: Roe are not on velvet with fallow deer. Behav. Ecol. Soc. 2011, 65, 875–887. [Google Scholar] [CrossRef]
- Ferretti, F.; Lovari, S. Introducing Aliens: Problems Associated with Invasive Exotics. In Behaviour and Management of European Ungulates, 1st ed.; Putman, R., Apollonio, M., Eds.; Whittles Publishing: Dunbeath, UK, 2014. [Google Scholar]
- Thirgood, S.J. The effects of sex, season and habitat availability on patterns of habitat use by fallow deer (Dama dama). J. Zool. 1995, 235, 645–659. [Google Scholar] [CrossRef]
- Apollonio, M.; Focardi, S.; Toso, S.; Nacci, L. Habitat selection and group formation pattern of fallow deer Dama dama in a submediterranean environment. Ecography 1998, 21, 225–234. [Google Scholar] [CrossRef]
- Thirgood, S.J. Alternative Mating Strategies and Reproductive Success in Fallow Deer. Behaviour 1991, 116, 1–10. [Google Scholar] [CrossRef]
- Ferretti, F.; Sforzi, A.; Lovari, S. Intolerance amongst deer species at feeding: Roe deer are uneasy banqueters. Behav. Proc. 2008, 78, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Rautiainen, H.; Bergvall, U.A.; Felton, A.M.; Tigabu, M.; Kjellander, P. Nutritional niche separation between native roe deer and the nonnative fallow deer—A test of interspecific competition. Mamm. Res. 2021, 66, 443–455. [Google Scholar] [CrossRef]
- Balik, B.; Moskalik, T.; Sadowski, J.; Zastocki, D. Selected aspects of forest protection from game. Stud. Mater. CEPL 2016, 46, 181–191. [Google Scholar]
- Wawrzyniak, P. Dynamika liczebności, jej wpływ na środowisko bytowania, a konieczność zarządzania populacja łosia Alces alces w Polsce. In Zarządzanie Populacjami Zwierząt; Polski Związek Łowiecki, Łowiec Polski: Warsaw, Poland, 2016; pp. 17–27. [Google Scholar]
- Wawrzyniak, P. Funkcjonowanie Populacji łosia (Alces alces), a Skala Konfliktów z Działalnością Człowieka w Warunkach Polski Północno-Wschodniej. 2017. Available online: https://wwwuwmedupl/wbz/sites/default/files/uploads/WBZ/Dokumenty/Katedry/KHZFiL/Konferencja24-11-2017/2_wawrzyniak_p_uwm_2017_lospdf (accessed on 8 September 2023).
- Bobek, B.; Błaszczyk, J.; Frąckowiak, W.; Furtek, J.; Morow, K.; Wojciuch-Poskonka, M.; Wyrobek, K. Łoś Alces Alces-Biologia, Ochrona i Gospodarka Łowiecka; Międzynarodowy Instytut Ekologii: Kraków, Poland, 2021. [Google Scholar]
- Nasiadka, P.; Wajdzik, M.; Skubis, J. A comprehensive over 100 years history of mouflon (Ovis musimon) in Poland: From the promising beginning in 1902 to questionable future in 2014. A case study of wildlife management history. Appl. Ecol. Environ. Res. 2021, 19, 993–1017. [Google Scholar] [CrossRef]
- Vincent, J.; EBideau, E.; Hewison, A.; Angibault, J. The influence of increasing density on body weight, kid production, home range and winter grouping in roe deer (Capreolus capreolus). J. Zool. 1995, 236, 371–382. [Google Scholar] [CrossRef]
- Andersen, R.; Linnell, J.D.C. Irruptive potential in roe deer: Density-dependent effects on body mass and fertility. J. Wildl. Manag. 2000, 64, 698–706. [Google Scholar] [CrossRef]
- Clutton Brock, T.H.A.; Illius, W.; Wilson, K.; Grenfell, B.T.; Maccoll, A.D.C.; Albon, S.D. Stability and Instability in Ungulate Populations: An Empirical Analysis. Am. Nat. 1997, 149, 195–219. [Google Scholar] [CrossRef]
- Genovesi, P. Eradications of invasive alien species in Europe: A review. Biol. Invasions 2005, 7, 127–133. [Google Scholar] [CrossRef]
- Gryz, J.; Chojnacka-Ożga, L.; Krauze-Gryz, D. Long-term stability of tawny owl (Strix aluco) population despite varying environmental conditions—A case study from central Poland. Pol. J. Ecol. 2019, 67, 75–83. [Google Scholar] [CrossRef]


| Variables | H | Df | p Value |
|---|---|---|---|
| Area | 14.1373 | 1 | <0.001 |
| Period | 0.0578 | 1 | >0.05 |
| Area ∗ Period | 10.6360 | 1 | <0.01 |
| Variables | Estimate | Std. Error | z Value | p Value |
|---|---|---|---|---|
| Intercept (roe deer, Głuchów) | 0.30438 | 0.05616 | 5.420 | <0.0001 |
| Fallow deer | −0.10178 | 0.04362 | −2.333 | <0.05 |
| Location_Spała | −0.84147 | 0.10231 | −8.2255 | <0.0001 |
| Fallow deer ∗ Location_Spała | −1.09219 | 0.41819 | −2.612 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryz, J.; Krauze-Gryz, D.; Jasińska, K.D. Alien vs. Native—Influence of Fallow Deer (Dama dama) Introduction on the Native Roe Deer (Capreolus capreolus) Population. Forests 2024, 15, 1014. https://doi.org/10.3390/f15061014
Gryz J, Krauze-Gryz D, Jasińska KD. Alien vs. Native—Influence of Fallow Deer (Dama dama) Introduction on the Native Roe Deer (Capreolus capreolus) Population. Forests. 2024; 15(6):1014. https://doi.org/10.3390/f15061014
Chicago/Turabian StyleGryz, Jakub, Dagny Krauze-Gryz, and Karolina D. Jasińska. 2024. "Alien vs. Native—Influence of Fallow Deer (Dama dama) Introduction on the Native Roe Deer (Capreolus capreolus) Population" Forests 15, no. 6: 1014. https://doi.org/10.3390/f15061014
APA StyleGryz, J., Krauze-Gryz, D., & Jasińska, K. D. (2024). Alien vs. Native—Influence of Fallow Deer (Dama dama) Introduction on the Native Roe Deer (Capreolus capreolus) Population. Forests, 15(6), 1014. https://doi.org/10.3390/f15061014