
Volatile Organic Compounds (VOCs) in Mediterranean Oak Forests of Hungarian Oak (Quercus frainetto Ten) Affected by Dieback Phenomena

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Sampling and Dendrochronological Data
2.3. Sample Preparation
2.4. Statistical Analyses
2.5. Solid Phase Micro-Extraction–GC/MS Analyses
3. Results and Discussion
3.1. Dendrochronological Analysis
3.2. SPME-GC/MS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Gentilesca, T.; Camarero, J.J.; Colangelo, M.; Nolã, A.; Ripullone, F.; Nole, A. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. IForest 2017, 10, 796–806. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Ofori-Amanfo, K.K.; Klem, K.; Veselá, B.; Holub, P.; Agyei, T.; Juráň, S.; Grace, J.; Marek, M.V.; Urban, O. The effect of elevated CO2 on photosynthesis is modulated by nitrogen supply and reduced water availability in Picea abies. Tree Physiol. 2023, 43, 925–937. [Google Scholar] [CrossRef]
- Rita, A.; Camarero, J.J.; Nolè, A.; Borghetti, M.; Brunetti, M.; Pergola, N.; Serio, C.; Vicente-Serrano, S.M.; Tramutoli, V.; Ripullone, F. The impact of drought spells on forests depends on site conditions: The case of 2017 summer heat wave in southern Europe. Glob. Chang. Biol. 2020, 26, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Quentin, A.G.; Beadle, C.L.; O’grady, A.P.; Pinkard, E.A. Effects of partial defoliation on closed canopy Eucalyptus globulus Labilladière: Growth, biomass allocation and carbohydrates. For. Ecol. Manag. 2011, 261, 695–702. [Google Scholar] [CrossRef]
- Iqbal, N.; Masood, A.; Khan, N.A. Analyzing the significance of defoliation in growth, photosynthetic compensation and source-sink relations. Photosynthetica 2012, 50, 161–170. [Google Scholar] [CrossRef]
- Hagedorn, F.; Joseph, J.; Peter, M.; Luster, J.; Pritsch, K.; Geppert, U.; Kerner, R.; Molinier, V.; Egli, S.; Schaub, M.; et al. Recovery of trees from drought depends on belowground sink control. Nat. Plants 2016, 2, 16111. [Google Scholar] [CrossRef]
- Schmid, S.; Palacio, S.; Hoch, G. Growth reduction after defoliation is independent of CO2 supply in deciduous and evergreen young oaks. New Phytol. 2017, 214, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Furze, M.E.; Wainwright, D.K.; Huggett, B.A.; Knipfer, T.; McElrone, A.J.; Brodersen, C.R. Ecologically driven selection of nonstructural carbohydrate storage in oak trees. New Phytol. 2021, 232, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Zeppel, M.J.B.; Anderegg, W.R.L.; Bloemen, J.; De Kauwe, M.G.; Hudson, P.; Ruehr, N.K.; Powell, T.L.; von Arx, G.; Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: Processes and trade-offs. Ecol. Res. 2018, 33, 839–855. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gazol, A.; Gentilesca, T.; Ripullone, F. Size matters a lot: Drought-affected Italian oaks are smaller and show lower growth prior to tree death. Front. Plant Sci. 2017, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Ripullone, F.; Camarero, J.J.; Colangelo, M.; Voltas, J. Variation in the access to deep soil water pools explains tree-to-tree differences in drought-triggered dieback of Mediterranean oaks. Tree Physiol. 2020, 40, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, M.; Camarero, J.J.; Battipaglia, G.; Borghetti, M.; De Micco, V.; Gentilesca, T.; Ripullone, F. A multi-proxy assessment of dieback causes in a Mediterranean oak species. Tree Physiol. 2017, 37, 617–631. [Google Scholar] [CrossRef]
- Cailleret, M.; Dakos, V.; Jansen, S.; Robert, E.M.R.; Aakala, T.; Amoroso, M.M.; Antos, J.A.; Bigler, C.; Bugmann, H.; Caccianaga, M.; et al. Early-warning signals of individual tree mortality based on annual radial growth. Front. Plant Sci. 2019, 9, 1964. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Gärtner, H.; Nievergelt, D. The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 2010, 28, 85–92. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.R.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J.; et al. Low growth resilience to drought is related to future mortality risk in trees. Nat. Commun. 2020, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Wajs, A.; Pranovich, A.; Reunanen, M.; Willför, S.; Holmbom, B. Headspace-SPME analysis of the sapwood and heartwood of Picea abies, Pinus sylvestris and Larix decidua. J. Essent. Oil Res. 2007, 19, 125–133. [Google Scholar] [CrossRef]
- Mecca, M.; Todaro, L.; Lo Giudice, V.; Lovaglio, T.; D’Auria, M. GC-MS and SPME techniques highlighted contrasting chemical behaviour in the water extractives of modified Castanea sativa mill. and Fagus sylvatica L. wood. Forests 2021, 12, 986. [Google Scholar] [CrossRef]
- Levanič, T.; Čater, M.; McDowell, N.G. Associations between growth, wood anatomy, carbon isotope discrimination and mortality in a Quercus robur forest. Tree Physiol. 2011, 31, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Gentilesca, T.; Camele, I.N.; Colangelo, M.; Lauteri, M.; Lapolla, A.; Ripullone, F. Oak forest decline in southern Italy: The study case of Gorgoglione forest. In ATTI del Secondo Congresso Internazionale di Selvicoltura; Accademia Italiana di Scienze Forestali: Florence, Italy, 2015; Volume 2, pp. 123–129. [Google Scholar]
- Marques, I.G.; Campelo, F.; Rivaes, R.; Albuquerque, A.; Ferreira, M.T.; Rodríguez-González, P.M. Tree rings reveal long-term changes in growth resilience in Southern European riparian forests. Dendrochronologia 2018, 52, 167–176. [Google Scholar] [CrossRef]
- Duval, C.J.; Gourrat, K.; Perre, P.; Prida, A.; Gougeon, R.D. A HS–SPME–GC–MS analysis of IR heated wood: Impact of the water content on the depth profile of oak wood aromas extractability. Food Res. Int. 2013, 54, 277–284. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Virjamo, V.; Ghimire, R.P.; Blande, J.D.; Julkunen-Tiitto, R.; Kivimäenpää, M. Climate change effects on secondary compounds of forest trees in the northern hemisphere. Front. Plant Sci. 2018, 9, 395200. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Introduction: Biochemistry, physiology and ecological functions of secondary metabolites. Annu. Plant Rev. 2010, 40, 1–19. [Google Scholar]
- Lindroth, R.L. Atmospheric change, plant secondary metabolites and ecological interactions. In The Ecology of Plant Secondary Metabolites: From Genes to Global Processes; Cambridge University Press: Cambridge, UK, 2012; pp. 120–153. [Google Scholar]
- Lämke, J.S.; Unsicker, S.B. Phytochemical variation in treetops: Causes and consequences for tree-insect herbivore interactions. Oecologia 2018, 187, 377–388. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. Jasmonates: News on occurrence, biosynthesis, metabolism and action of an ancient group of signaling compounds. Int. J. Mol. Sci. 2018, 19, 2539. [Google Scholar] [CrossRef]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- De Carvalho, C.C.; Caramujo, M.J. The various roles of fatty acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in the integration of cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free Radic. Biol. Med. 2018, 122, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Kretovich, V.L. Molecular mechanisms of nitrogen assimilation by plants. Mol. Mech. Nitrogen Assim. Plants 1980, 29. [Google Scholar]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant–pathogen interactions. Front. Plant Sci. 2017, 8, 255703. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Bartram, S.; Jux, A.; Gleixner, G.; Boland, W. Dynamic pathway allocation in early terpenoid biosynthesis of stress-induced lima bean leaves. Phytochemistry 2006, 67, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.N.M.; Balakrishnan, R.S.; Shamsudeen, S.; Bahwani, S.A.; Adam, F. A concise review of the natural existance, synthesis, properties, and applications of syringaldehyde. BioResources 2012, 7, 4377–4399. [Google Scholar] [CrossRef]
- Sangeetha, C.; Krishnamoorthy, A.S.; Amirtham, D. Antifungal bioactive compounds from Chinese caterpillar fungus (Ophiocordyceps sinensis (Berk.) GH Sung et al.) against plant pathogens. Madras Agric. J. 2015, 102, 353–357. [Google Scholar]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef]
- Janaki, S.; Zandi-Sohani, N.; Ramezani, L.; Szumny, A. Chemical composition and insecticidal efficacy of Cyperus rotundus essential oil against three stored product pests. Int. Biodeterior. Biodegrad. 2018, 133, 93–98. [Google Scholar] [CrossRef]
- Langsi, J.D.; Nukenine, E.N.; Oumarou, K.M.; Moktar, H.; Fokunang, C.N.; Mbata, G.N. Evaluation of the insecticidal activities of α-Pinene and 3-Carene on Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Insects 2020, 11, 540. [Google Scholar] [CrossRef] [PubMed]
- Sfara, V.; Zerba, E.N.; Alzogaray, R.A. Fumigant insecticidal activity and repellent effect of five essential oils and seven monoterpenes on first-instar nymphs of Rhodnius prolixus. J. Med. Entomol. 2014, 46, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, R.; Triadiati, T.; Falah, S. Induction of agarwood in Aquilaria malaccensis using nitrogen fertilizer and Fusarium solani. J. Penelit. Kehutan. Wallacea 2018, 7, 165–171. [Google Scholar] [CrossRef]
- Martinez, R.G.; De La Serrana, H.L.G.; Mir, M.V.; Granados, J.Q.; Martinez, M.L. Influence of wood heat treatment, temperature and maceration time on vanillin, syringaldehyde, and gallic acid contents in oak wood and wine spirit mixtures. Am. J. Enol. Vitic. 1996, 47, 441–446. [Google Scholar] [CrossRef]
- Abbas, F.; O’Neill Rothenberg, D.; Zhou, Y.; Ke, Y.; Wang, H.C. Volatile organic compounds as mediators of plant communication and adaptation to climate change. Physiol. Plant. 2022, 174, e13840. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; D’Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; Mauch-Mani, B.; Turlings, T.C. Priming by airborne signals boosts direct and indirect resistance in maize. Plant J. 2007, 49, 16–26. [Google Scholar] [CrossRef]
- Camarero, J.J.; Sangüesa-Barreda, G.; Vergarechea, M. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak speciesk. Ann. For. Sci. 2016, 73, 341–351. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
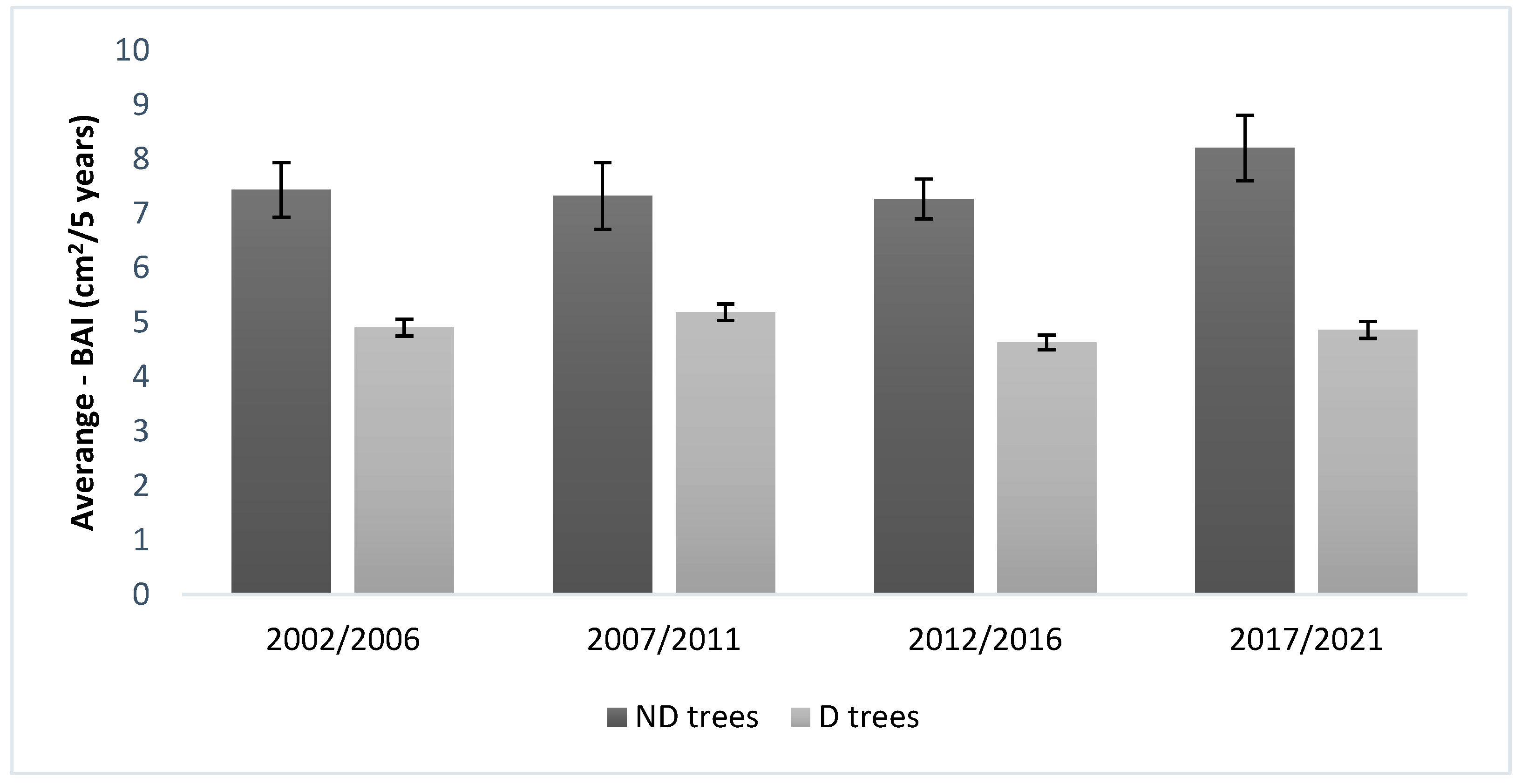
| Years | Average BAI (cm2/5 Years) ND Trees | SD | SE | Average BAI (cm2/5 Years) D Trees | SD | SE |
|---|---|---|---|---|---|---|
| 2002/2006 | 7.44 | 1.12 | 0.50 | 4.90 | 0.34 | 0.15 |
| 2007/2011 | 7.33 | 1.36 | 0.61 | 5.19 | 0.34 | 0.15 |
| 2012/2016 | 7.27 | 0.82 | 0.36 | 4.63 | 0.30 | 0.14 |
| 2017/2021 | 8.21 | 1.35 | 0.60 | 4.87 | 0.35 | 0.16 |
| Class of Compounds | Compound | RI | 1D | 2D | 3D | |||||||||
| I | II | III | IV | I | II | III | IV | I | II | III | IV | |||
| Acids | Acetic acid | 655 | 2.75 | 1.35 | 1.37 | 1.27 | 1.37 | 1.83 | 4.15 | 3.42 | 2.83 | 1.35 | 1.38 | 1.28 |
| Aromatic Aldehydes | Vanillin | 1407 | 6.88 | 0.92 | 10.95 | 2.95 | 18.90 | 6.53 | 2.68 | 8.08 | ||||
| Syringaldehyde | 1662 | 4.15 | 3.15 | 2.05 | 1.90 | 3.95 | 4.00 | 2.60 | 2.43 | 1.90 | 1.82 | 1.72 | 1.63 | |
| Coniferaldehyde | 1741 | |||||||||||||
| Sinapaldehyde | 1909 | 1.27 | 4.17 | 1.42 | ||||||||||
| Aliphatic Aldehydes | 3-methylbutanal | 659 | ||||||||||||
| Heptanal | 1176 | |||||||||||||
| Decanal | 1182 | |||||||||||||
| Octanal | 1282 | |||||||||||||
| Nonanal | 1387 | |||||||||||||
| Furfural | 1462 | |||||||||||||
| Alkanes | Eicosane | 2000 | ||||||||||||
| Nitrogen Compounds | Diethyltoluamide | 1571 | 6.30 | 4.75 | 4.48 | 20.70 | 20.10 | 6.52 | 11.58 | 11.38 | ||||
| N-p-bromophenylselenoacetamide | 1724 | 1.05 | ||||||||||||
| Dimethyl palmitamine | 1894 | |||||||||||||
| Aromatics | Xylene | 1137 | ||||||||||||
| 1,1,3-trimethyl-3-phenyl-2H-indene | 1714 | |||||||||||||
| Methyl n-hydroxybenzimidate | 1814 | 1.05 | 0.83 | 2.45 | 1.27 | 1.35 | 2.88 | 4.70 | ||||||
| 2,6-dimethoxyphenol | 2269 | 2.05 | 3.25 | |||||||||||
| 2,6-di-tert-butylphenol | 2327 | |||||||||||||
| Fatty acids | Nonanoic acid | 1265 | 0.37 | |||||||||||
| Pentanoic acid | 1685 | 0.60 | ||||||||||||
| Tetradecanoic acid | 1768 | |||||||||||||
| Hexadecanoic acid | 1968 | 2.62 | 2.75 | 2.15 | 2.45 | |||||||||
| Hexanedioic acid | 2137 | 2.22 | 2.03 | 1.98 | ||||||||||
| Octadecanoic acid | 2182 | 1.55 | 1.32 | |||||||||||
| Ketones | 4-oxo-β-dihydroionone | 1659 | 12.37 | 3.75 | ||||||||||
| 3-oxo-7,8-dihydro-α-ionone | 1731 | 1.52 | 1.70 | 5.75 | 6.35 | 12.22 | 2.10 | 2.35 | ||||||
| Silanols | Dimethylsilanediol | 649 | 8.22 | 9.33 | 7.43 | 7.05 | 3.43 | 3.72 | 7.03 | 5.48 | 5.50 | 7.02 | 7.63 | 7.28 |
| Terpenes | α-pinene | 936 | ||||||||||||
| Pinocarvone | 1138 | |||||||||||||
| Cyperene | 1398 | |||||||||||||
| α-bergamotene | 1407 | |||||||||||||
| β-panasinsene | 1527 | |||||||||||||
| γ-eudesmol | 1632 | |||||||||||||
| Class of Compounds | Compound | 1ND | 2ND | 3ND | ||||||||||
| I | II | III | IV | I | II | III | IV | I | II | III | IV | |||
| Acids | Acetic acid | 655 | ||||||||||||
| Aromatic Aldehydes | Vanillin | 1407 | 0.43 | 11.73 | 18.20 | 16.05 | 13.20 | 2.32 | 2.13 | |||||
| Syringaldehyde | 1662 | 14.45 | 13.20 | 12.20 | 6.40 | |||||||||
| Coniferaldehyde | 1741 | 17.13 | 20.38 | 12.40 | 8.35 | |||||||||
| Sinapaldehyde | 1909 | |||||||||||||
| Aliphatic Aldehydes | 3-methylbutanal | 659 | 0.93 | |||||||||||
| Heptanal | 1176 | 1.02 | 2.05 | 2.65 | 2.45 | 4.40 | 0.97 | 4.25 | 2.43 | 1.88 | ||||
| Decanal | 1182 | 0.58 | 16.45 | 3.27 | 4.77 | 4.27 | 2.13 | 1.77 | 7.92 | 5.57 | ||||
| Octanal | 1282 | 0.62 | 8.10 | 3.17 | 3.32 | |||||||||
| Nonanal | 1387 | 1.25 | 7.90 | 9.63 | 4.82 | 7.08 | ||||||||
| Furfural | 1462 | 0.52 | 2.07 | 0.70 | ||||||||||
| Alkanes | Eicosane | 2000 | 2.67 | 7.20 | 4.28 | |||||||||
| Nitrogen Compounds | Diethyltoluamide | 1571 | 5.87 | 8.47 | ||||||||||
| N-p-bromophenylselenoacetamide | 1724 | |||||||||||||
| Dimethyl palmitamine | 1894 | 5.93 | ||||||||||||
| Aromatics | Xylene | 1137 | 0.53 | 4.72 | 1.93 | |||||||||
| 1,1,3-trimethyl-3-phenyl-2H-indene | 1714 | 0.52 | 0.43 | 0.48 | ||||||||||
| Methyl n-hydroxybenzimidate | 1814 | 1.03 | 1.60 | 0.45 | 2.72 | 1.30 | 0.72 | |||||||
| 2,6-dimethoxyphenol | 2269 | 5.93 | 4.70 | 1.35 | ||||||||||
| 2,6-di-tert-butylphenol | 2327 | 2.27 | 1.73 | |||||||||||
| Fatty acids | Nonanoic acid | 1265 | 1.88 | 4.90 | ||||||||||
| Pentanoic acid | 1685 | 0.30 | 4.05 | 2.87 | 2.87 | 1.93 | 3.55 | 2.40 | ||||||
| Tetradecanoic acid | 1768 | 1.37 | 1.10 | 0.30 | 1.23 | |||||||||
| Hexadecanoic acid | 1968 | 20.75 | 0.98 | 1.92 | 1.55 | 2.08 | 2.12 | 1.63 | ||||||
| Hexanedioic acid | 2137 | 1.87 | 2.10 | 1.77 | 5.07 | 3.88 | ||||||||
| Octadecanoic acid | 2182 | 17.57 | 1.40 | 1.60 | 9.63 | |||||||||
| Ketones | 4-oxo-β-dihydroionone | 1659 | ||||||||||||
| 3-oxo-7,8-dihydro-α-ionone | 1731 | |||||||||||||
| Silanols | Dimethylsilanediol | 649 | 0.17 | 2.30 | 0.28 | 2.70 | 3.43 | 2.52 | ||||||
| Terpenes | α-pinene | 936 | 3.62 | 2.87 | ||||||||||
| Pinocarvone | 1138 | 1.02 | 1.40 | |||||||||||
| Cyperene | 1398 | 1.18 | 1.37 | 1.13 | ||||||||||
| α-bergamotene | 1407 | 0.60 | 1.08 | 0.98 | ||||||||||
| β-panasinsene | 1527 | 1.80 | 0.43 | 1.62 | ||||||||||
| γ-eudesmol | 1632 | 1.27 | 1.70 | |||||||||||
| Wood Type | n | Acids | Aromatic Aldehydes | Aliphatic Aldehydes | Alkanes | Nitrogen Compounds | Aromatics | Fatty Acids | Ketones | Silanols | Terpenes | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| average | D | 12 | 2.0 | 4.5 | n.d. | n.d. | 9.7 | 2.2 | 1.8 | 5.3 | 6.6 | n.d. |
| st. dev | 1.0 | 4.1 | 6.5 | 1.2 | 0.7 | 4.1 | 1.7 | |||||
| average | ND | 12 | n.d. | 10.6 | 3.8 | 4.7 | 6.8 | 1.9 | 3.6 | n.d. | 1.9 | 1.5 |
| st. dev | 5.9 | 3.4 | 1.9 | 1.2 | 1.6 | 4.6 | 1.2 | 0.8 | ||||
| t Test (prob) | 0.008 | n.d. | n.d. | 0.502 | 0.649 | 0.220 | n.d. | 0.000 | n.d. | |||
| significance | ** | n.s. | n.s. | n.s. | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mecca, M.; Todaro, L.; D’Auria, M.; Italiano, S.S.P.; Sofo, A.; Ripullone, F. Volatile Organic Compounds (VOCs) in Mediterranean Oak Forests of Hungarian Oak (Quercus frainetto Ten) Affected by Dieback Phenomena. Forests 2024, 15, 1072. https://doi.org/10.3390/f15061072
Mecca M, Todaro L, D’Auria M, Italiano SSP, Sofo A, Ripullone F. Volatile Organic Compounds (VOCs) in Mediterranean Oak Forests of Hungarian Oak (Quercus frainetto Ten) Affected by Dieback Phenomena. Forests. 2024; 15(6):1072. https://doi.org/10.3390/f15061072
Chicago/Turabian StyleMecca, Marisabel, Luigi Todaro, Maurizio D’Auria, Santain Settimio Pino Italiano, Adriano Sofo, and Francesco Ripullone. 2024. "Volatile Organic Compounds (VOCs) in Mediterranean Oak Forests of Hungarian Oak (Quercus frainetto Ten) Affected by Dieback Phenomena" Forests 15, no. 6: 1072. https://doi.org/10.3390/f15061072
APA StyleMecca, M., Todaro, L., D’Auria, M., Italiano, S. S. P., Sofo, A., & Ripullone, F. (2024). Volatile Organic Compounds (VOCs) in Mediterranean Oak Forests of Hungarian Oak (Quercus frainetto Ten) Affected by Dieback Phenomena. Forests, 15(6), 1072. https://doi.org/10.3390/f15061072