Abstract
Bacterial communities are vital factors for regulating litter decomposition in forests. With the gradual increase in global climate change, the effects of global warming on the decomposition of forest litter have become a cause for concern. In this study, we used a displacement test to simulate the effects of warming (+2.5, +4.6, and +6.5 °C) on the composition and diversity of bacterial communities in Chinese fir litters based on the natural differences in temperature between the elevation of Wuyi Mountain. Warming significantly increased the abundance and diversity of bacterial communities and altered the functional abundance of bacterial communities. Warming significantly increased the relative abundance of Acidothermus and decreased that of Bacteroidetes and Mucilaginibacter. Structural variations in the bacterial communities were closely related to the chemical properties of the litter. The relative abundance of Planctomycetes exhibited a positive association with nitrogen content and a negative association with cellulose content, whereas phosphorus content was the main driving factor of Acidothermus abundance. Warming increased the complexity of the bacterial community structure, which promoted the decomposition of Chinese fir litter and accelerated nutrient cycling in the Chinese fir plantation ecosystem.
1. Introduction
Litter is an important carrier of nutrient cycling in forest ecosystems, and its decomposition determines the storage and turnover of organic matter [1]. Temperature is a vital climatic factor affecting litter decomposition, and a slight change directly affects its decomposition rate [2]. IPCC AR6 reported that the global temperature rise will exceed 1.5 °C by 2030 and reach 3.2 °C by 2100 [3]. Rising global temperatures will change the litter decomposition process, which has a direct effect on carbon cycling and the ecological functions of forest ecosystems [4]. Therefore, insights into the response mechanism of forest litter decomposition to warming are of great significance for predicting the future processes of nutrient cycling in forest ecosystems.
Over the past decade, the responses of litter decomposition to warming in forest ecosystems have been widely studied. However, most of these studies are based on human-controlled experiments, such as indoor microenvironment simulation (artificial climate chambers [5] and greenhouses [6]) and in situ heating methods (resistance heating cables [7] and open-top chambers [8]), which have defects such as large differences between test and natural conditions in the field, as well as being underrepresented. Elevation gradient research was recognized as a powerful tool for exploring the processes of warming impact on ecosystems [9]. The process of litter decomposition in a field environment can be more accurately reconstructed using the natural temperature difference between altitudinal gradients to perform a displacement test of litter decomposition. Salinas et al. [10] found that soil temperature explained 95% of the variation in the annual weight loss rate of forest litter using a displacement test. Klimek et al. [11] found that temperature range was the main factor affecting litter respiration rate and microbial biomass in the field experiment of litter translocation along the altitudinal gradient, and microbial biomass increased with the extension of decomposition time. Bohara et al. [12] found that temperature was the best predictor of litter mass loss in the study of in situ litter decomposition along the elevation gradient. Therefore, the displacement test between altitudes was considered an effective way to simulate the effects of warming on forest litter decomposition. Wuyi Mountain is known as the first peak in East China and has obvious vertical distribution characteristics of climate zones [13], making it a good experimental site for this study.
Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) is an important quick-growing timber species in China. According to data from the 9th National Forest Resources Inventory, the planting area of Chinese fir is 9.87 × 106 ha, which is the largest afforestation species in China’s plantation area and is crucial in forestry production in China [14]. In recent years, with improvements in intensive forestry production, the problem of forest productivity decline caused by multi-generation continuous planting of Chinese fir plantations has become increasingly serious, which greatly limits the sustainable management and development of Chinese fir plantations. Studies have found that the slow decomposition rate and reduced nutrient return from litter in Chinese fir plantations may be the reasons for the decline in continuous planting productivity [15,16]. Microbial communities are vital drivers of forest litter decomposition and are essential for mass loss, nutrient conversion, and humus formation, explaining 85% of the annual weight loss rate of litter [17]. Previous studies have reported that bacteria can change the chemical composition and structure of litter and have a strong ability to degrade keratin and aromatic compounds in coniferous litter [18]. Furthermore, various enzymes produced by microbes are involved in ammonification, nitrification, and nutrient cycling during litter decomposition, which are closely related to the decomposition rate [19].
The bacterial community structure is usually regulated by temperature, and warming can increase the bacterial quantity when external environmental factors are consistent [20]. However, several studies have reported that environmental fluctuations and the reduced availability of unstable substrates caused by warming may result in a decline or maintenance of bacterial quantity [21]. However, the effects of warming on the bacterial community structure in Chinese fir litter remain unclear. Warming affects the litter decomposition process by changing the composition of the bacterial community, which affects forest productivity and plantation ecological function [22]. Therefore, further research on the effect of warming on the bacterial community structure in litter is important for a comprehensive understanding of the nutrient cycling process in the Chinese fir plantation ecosystem. In this study, the response of the decomposition of Chinese fir litter to warming (+2.5, +4.6, and +6.5 °C) was studied by using a remote displacement test based on the natural differences in temperature between the elevation of Wuyi Mountain. The differences in bacterial community structure under different temperature conditions were observed after 360 days of decomposition. This study provides a scientific basis for future litter management for Chinese fir plantations.
2. Materials and Methods
2.1. Study Site
The study area is located at the Nature Reserve of Wuyi Mountain National Park (117°27′–117°51′ E, 27°33′–27°54′ N) in the northwest of Fujian Province, China. The area has a subtropical monsoon climate, with an annual mean temperature of 8.5–18.0 °C and mean annual precipitation of 1486–2150 mm. The average temperature is about 27 °C in July and about 8 °C in January. The annual accumulated temperature from the mountaintop to the foothills is 1500–5800 °C, with evident vertical distribution characteristics of the climate zone. The soil types from bottom to top are red soil, yellow–red soil, yellow soil, and meadow soil. The vegetation types are subtropical evergreen broad-leaved forest, coniferous mixed forest, coniferous forest, sub-alpine dwarf forest, and alpine meadow, respectively. The dominant vegetation types include Phyllostachys edulis, Cunninghamia lanceolata, Pinus massoniana, and Castanopisi carlesii. Because the temperature between altitudes is highly dependent on slope and insolation, all plots were established on southeastern slopes.
2.2. Experimental Design
To ensure consistency in the initial quality of Chinese fir litter, experimental materials were collected from a 15-year-old Chinese fir plantation at the Xinkou National Forest Farm. We took the Chinese fir litter back to the laboratory, separated the twigs and leaves, and air-dried them. In this study, twigs and leaves were placed into nylon decomposition bags with a size of 20 × 20 cm and a mesh size of 0.5 mm, and each bag weighed 10 ± 0.005 g air-dried (2.5 and 7.5 g of twigs and leaves, respectively) [23]. Five bags of the prepared samples were used to determine the initial nutrients. The initial total carbon content was 503.69 ± 13.62 mg/g, total phosphorus content was 0.29 ± 0.07 mg/g, total nitrogen content was 13.98 ± 0.22 mg/g, and total potassium content was 0.83 ± 0.17 mg/g.
In September 2021, we selected four altitudes (1948, 1410, 1003, and 620 m) as decomposition test sites in Wuyi Mountain. Warming was simulated using the natural differences in temperature between altitudinal gradients. The average annual temperature at 1948 m was regarded as the control condition (W0). The differences between the annual mean temperature at 620 m (+6.5 °C), 1003 m (+4.6 °C), 1410 m (+2.5 °C), and 1948 m were considered as warming treatments. Three plots (20 m × 20 m) were set at each altitude temperature gradient, and the original surface litter layer was removed. The canopy density, tree height, and tree diameter at breast height (DBH) were measured in each plot (Table 1). The litterbags were fully in contact with the soil and arranged longitudinally; each litterbag was adjacent but not overlapping. Subsequently, we collected soil samples at 0–20 cm depth from the center and four corners of each plot. The mixed soil samples (0.5 kg) were returned for physical and chemical properties determination (Table 2). Finally, temperature recorders (TMS-4, Saifu Biotechnology, Shanghai, China) were installed at each altitude.

Table 1.
Plot characteristics in different sites (location, temperature, and growth conditions).
Table 2.
Soil physical and chemical properties of sample plots at different test sites.
After 360 days of decomposition, 36 litter bags (3 bags/plot × 3 plots × 4 altitudes) were harvested in September 2022. Disposable rubber gloves were worn during sampling, and impurities were removed. A portion was used for DNA extraction, another portion was used to determine the chemical properties, and the remainder was used to determine enzyme activity. Litterbags for the determination of bacterial community structure and enzyme activity were individually packed into zip-lock bags, transported at a low temperature using dry ice, and then the determination was completed as soon as possible.
2.3. Sequencing of Litter Bacterial Community
A HiPure soil DNA Kit (Magen, Guangzhou, China) was used to extract DNA from the Chinese fir litter. DNA quality and concentration were assessed using 1% agarose gel electrophoresis and a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). Primers 341 F (5′-CCTACGGGNGGCWGCAG-3′) and 806 R (5′-GGACTACHVGGGTATCTAAT-3′) were used to amplify the V3–V4 regions of the bacterial 16S rRNA. The thermal cycle parameters were as follows: 95 °C for 2 min, 98 °C for 10 s, 62 °C for 30 s, 68 °C for 30 s (30 cycles), and a final extension at 68 °C for 10 min [24]. The amplified products were purified by AMPure XP Beads (Beckman Agency, Muskogee, OK, USA), and the ABI StepOnePlus Real-Time PCR System (Life Technologies, Carlsbad, CA, USA) was used to quantify them. The products were sequenced using an Illumina NovaSeq 6000 (SanDiego, CA, USA).
FASTP (version 0.18.0) was used to further filter raw reads, and QIIME (version 1.9.1) was used to splice the bidirectional sequences to obtain high-quality effective tags. The effective tags were clustered into operational taxonomic units (OTUs) at 97% identity using UPARSE software (version 9.2.64). The tag sequence with the highest abundance was selected as a representative within each cluster. Bacterial OTUs were taxonomically assigned with the Silva database (http://www.arb-silva.de; accessed on 10 November 2022).
2.4. Measurements of Chemical Properties and Enzyme Activities
The samples for chemical properties determination were dried and sieved in a 0.178 mm mesh. In this study, an Element Analyzer (VARIO MAX CN, Hanau, Germany) was used to measure total C and N, and an inductively coupled plasma optical emission spectrometer (PerkinElmer, Richmond, CA, USA) was used to measure total P and K. An ANKOM 200/220 fiber analyzer (Macedon, NY, USA) was used to measure lignin and cellulose concentrations.
Enzyme activities related to the bacterial quantity were also studied, and enzyme activity was measured using a kit (SpectraMax M4, Silicon Valley, CA, USA). Sucrase activity assays were based on the ability to catalyze the degradation of sucrose to produce reducing sugars, which can produce brown-red amino compounds [25]. Urease activity was determined using the indophenol blue method [26], and the litter homogenization was incubated with 0.5 mL of 10% urea solution and 1 mL of citric acid buffer at 37 °C for 24 h. For peroxidase activity, 0.6 mL of 0.3% hydrogen peroxide was added. All reaction mixtures were incubated at 37 °C for 1 h and centrifuged for 2 min at 5000 rpm. It catalyzes the oxidation of organic matter to quinone [27]. Their absorbance was measured at 540 nm, 630 nm, and 430 nm, respectively.
2.5. Date Analysis
Chao1, Simpson, and Shannon indices and species classification were calculated using QIIME software (version 1.9.1). One-way variance analysis with the least significant difference (p < 0.05) was used to test the significance using SPSS (version 19.0 Statistics; SPSS Inc., Chicago, MI, USA). LEfSe software (version 1.0; Gene Denovo, Guangzhou, China) was used to analyze the biomarkers between the groups. The Functional Annotation of Prokaryotic Taxa (FAPROTAX) database was used to annotate bacterial community functions, and the functional abundance was subjected to a “Hellinger” transformation. The R “vegan” package was used for principal coordinate analysis (PCoA) and redundancy analysis (RDA). Origin (version 2021) was used to draw a bar chart and heat map of the correlation between the bacterial alpha diversity index, chemical properties, and enzyme activities. The partial least-squares path model analysis (PLS-PM) was conducted via the “plspm” package to estimate the effects of warming on the microbial community composition of litter.
3. Results
3.1. Effect of Warming on the Composition of Bacterial Communities in Litters
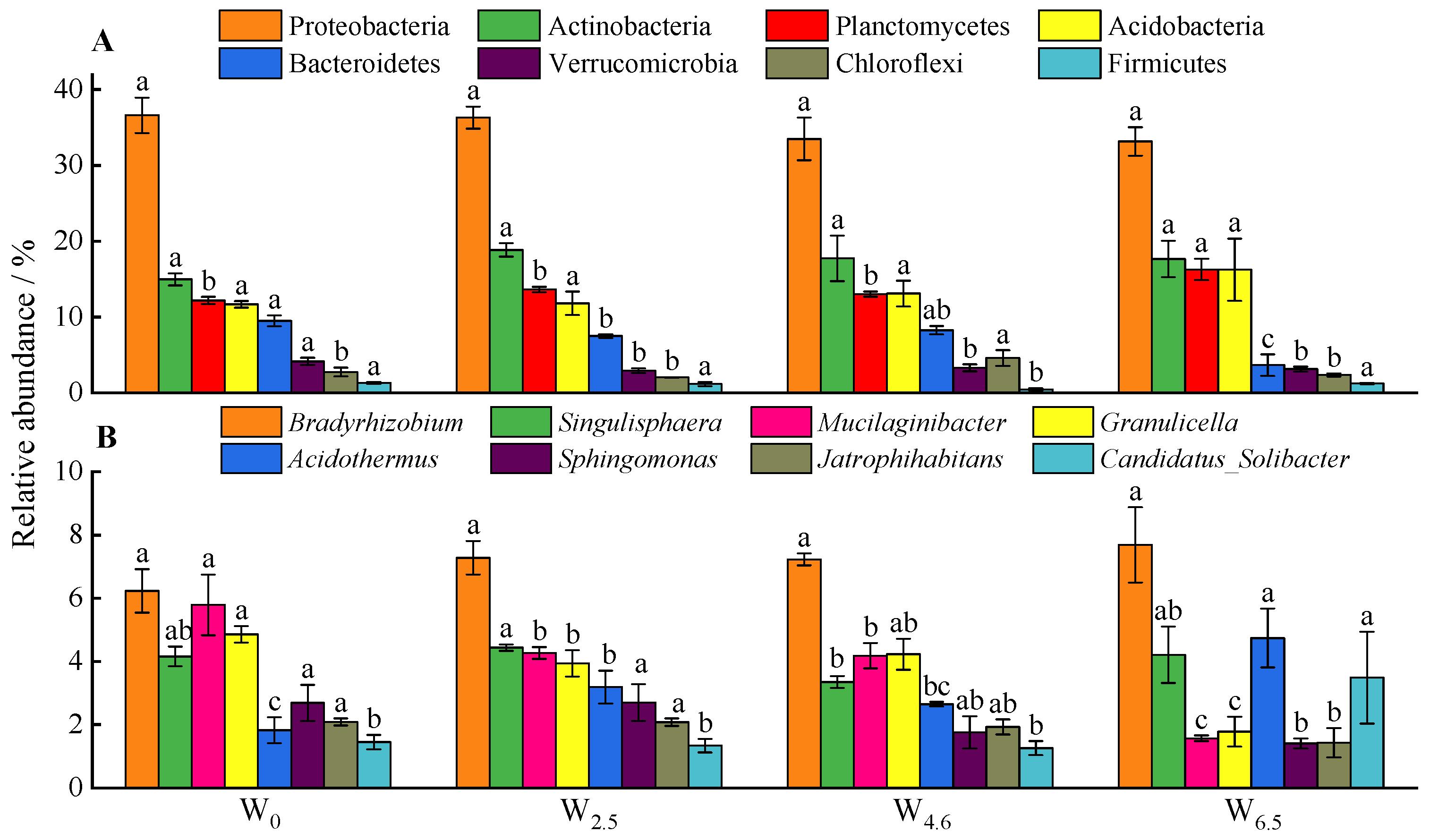
Generally, the composition of bacterial communities is closely related to temperature changes (Figure 1). At the phylum level, the relative abundance of Planctomycetes, Proteobacteria, Actinobacteria, and Acidobacteria was more than 10%. With the rising temperature, the relative abundance of Bacteroidetes decreased significantly. The differences in the relative abundance of Planctomycetes between W6.5 and other conditions were significant. Warming had no significant effects on the relative abundance of Actinobacteria, Proteobacteria, and Acidobacteria. At the genus level, the top three dominant genera of bacteria were Bradyrhizobium, Singulisphaera, and Mucilaginibacter. With an increase in temperature, the relative abundance of Mucilaginibacter and Granulicella significantly decreased, while that of Candidatus_Solibacter was highest at W6.5. The order of the relative abundance of Acidothermus under different temperature conditions was W6.5 > W2.5 > W4.6 > W0, and the differences between W6.5, W2.5, and W0 were significant.
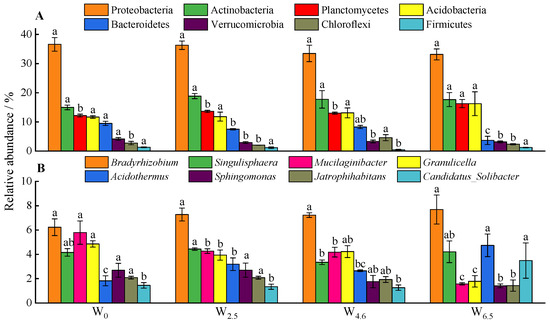
Figure 1.
Eight most abundant phyla (A) and genera (B) in litters. Different lowercase letters indicate significant differences between different temperature conditions of the same bacterial community abundance at the same taxonomic level (p < 0.05).
3.2. Analysis of Diversity and Biomarkers in Litters
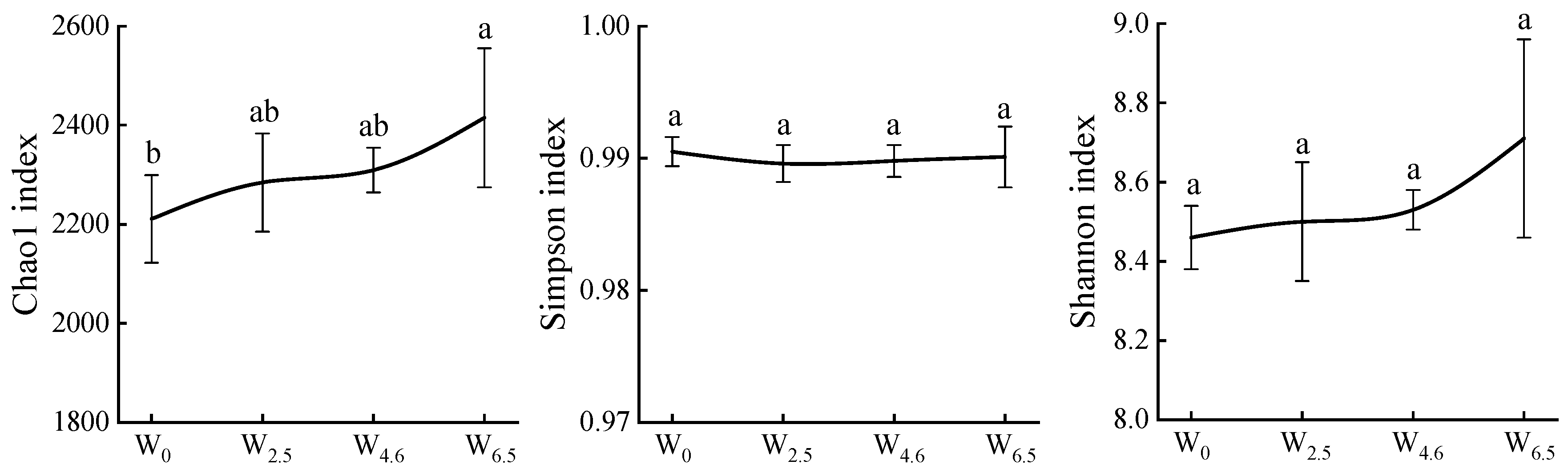
The Chao1 index is often used to measure changes in species richness, while the Simpson and Shannon indices are used to measure changes in community diversity. As shown in Figure 2, the Chao1 and Shannon indices showed an increasing trend with increasing temperature, while the Simpson index remained constant. Warming had no significant effects on the Simpson and Shannon indices.
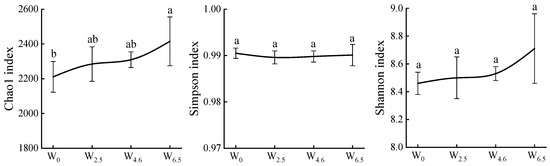
Figure 2.
Chao1, Simpson, and Shannon indices of bacterial communities in litters. Different lowercase letters indicate significant differences between different temperature conditions (p < 0.05).
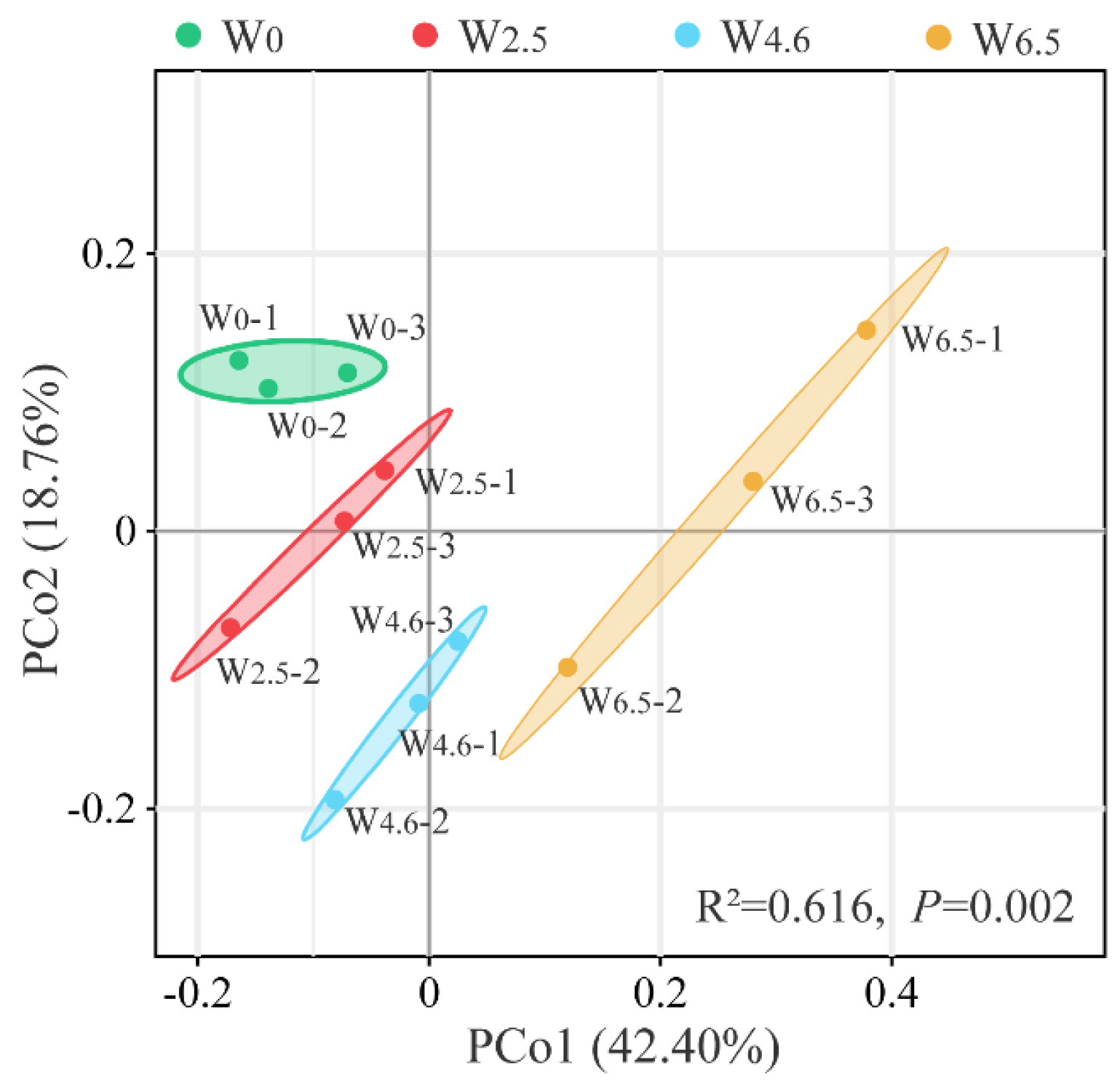
PCoA analysis was performed based on the OUT level and the Bary distance algorithm. As shown in Figure 3, the cumulative contribution of PCo1 (42.40%) and PCo2 (18.76%) was 61.16%. The bacterial communities in the same group clustered well, with no overlapping areas. It showed that temperature variation could explain the differences in the composition of bacterial communities in Chinese fir litters well. Combined with the Adonis (Permanova) intergroup difference test (R2 = 0.616, p = 0.002), the composition of the bacterial community in Chinese fir litter was significantly different between the control and warming conditions.
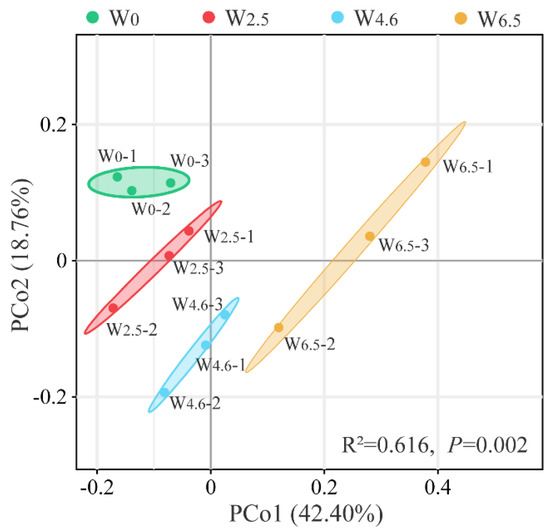
Figure 3.
PCoA analysis of bacterial communities in litters; 1, 2, and 3 indicate the repetition of 3 samples under the same temperature condition.
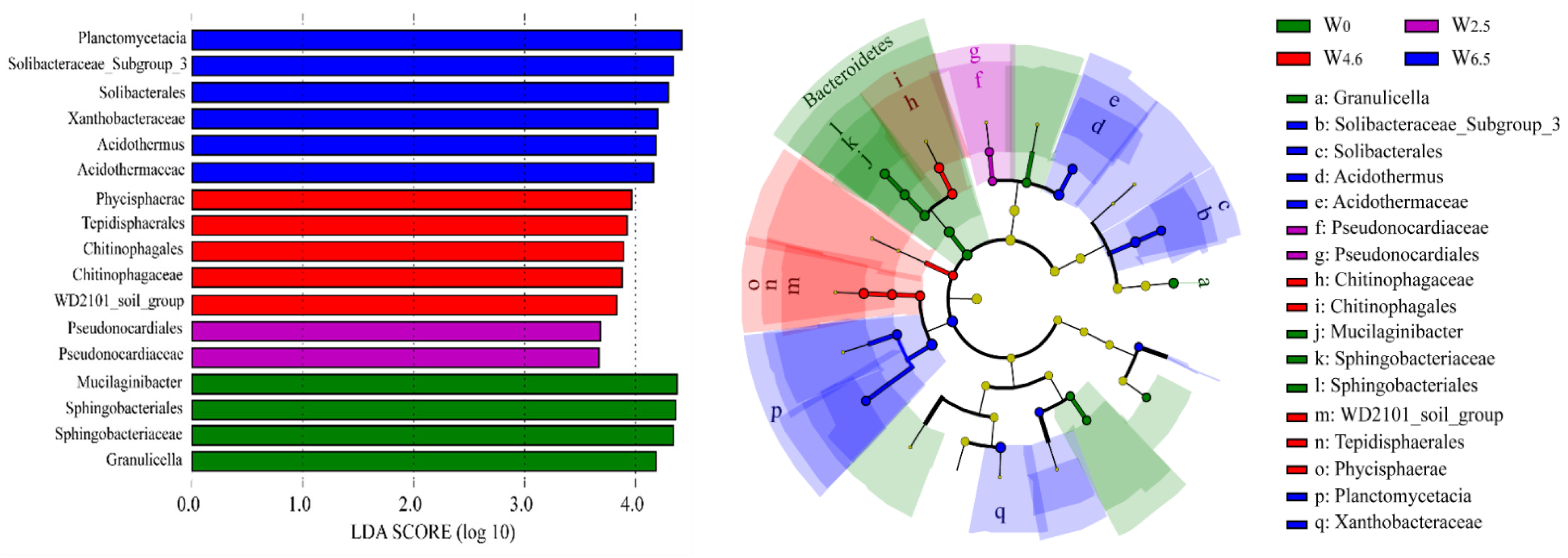
The LEfSe analysis showed that the number of biomarkers increased from the control condition to 6.5 °C warming when the LDA SCORE (LOG 10) was set to 3.5 (Figure 4). The biomarkers under control condition were Mucilaginibacter, Sphingobacteriales, Sphingobacteriaceae, and Granulicella. The biomarkers under 2.5 °C warming conditions were Pseudonocardiaceae and Pseudonocardiales. At the same time, the biomarkers under 6.5 °C warming conditions were Planctomycetacia, Solibacteraccac_Subgroup_3, Solibacterales, Xanthobacteraceae, Acidothermus, and Acidothermaceae.
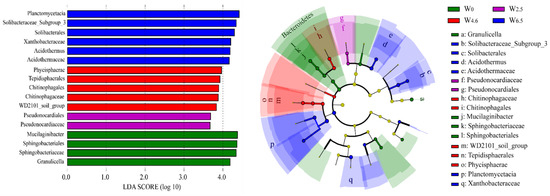
Figure 4.
LEfSe analysis of bacterial communities in litters. The yellow dots in the figure indicate no significant difference in the abundance of this bacterium at this level amongst different temperature conditions.
3.3. Effect of Warming on Bacterial Community Function in Litters
The FAPROTAX database was used to annotate bacterial community functions, and 50 functional groups were obtained. Heatmap analysis of the top 12 bacterial functions in abundance showed that the functional types were similar among the different treatments, but the functional abundances were different (Figure 5). The main functional type among the bacterial communities was fermentation (1.63%) under the control conditions. Under 6.5 °C warming conditions, the main types were cellulolysis (1.57%), oxygenic photoautotrophy (1.64%), and animal parasites or symbionts (1.6%).

Figure 5.
Abundance heatmap of bacterial community function prediction in litters.
3.4. Correlation Analysis of Chemical Properties and Enzyme Activity with Bacterial Community
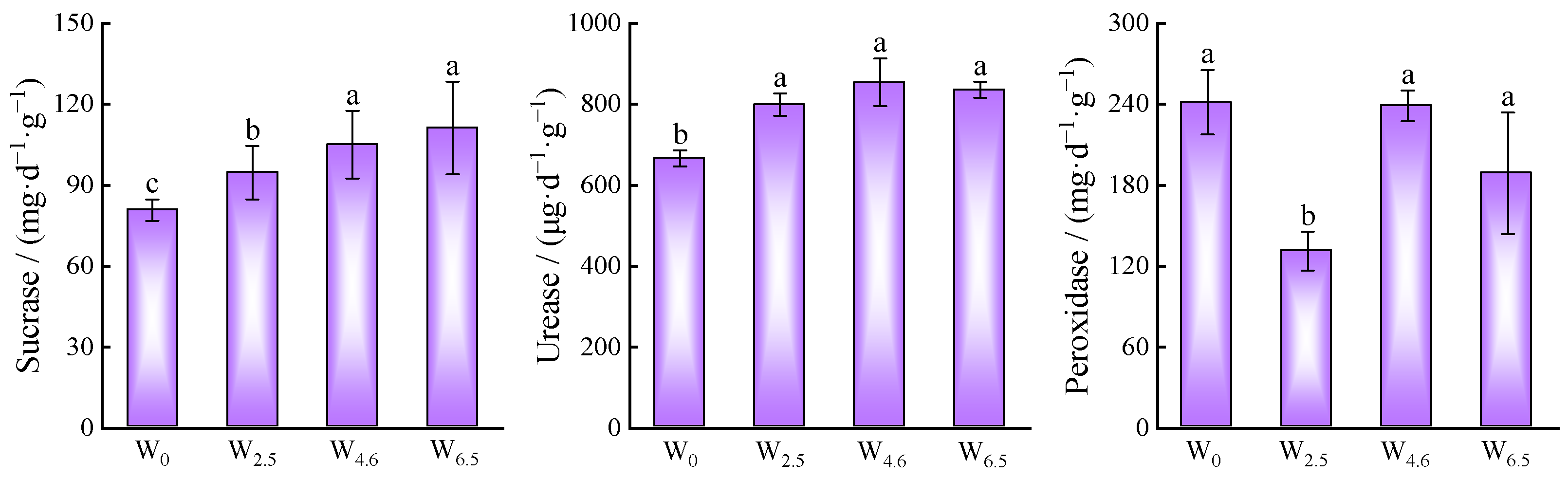
After 360 days of decomposition, N, P, and K showed an increasing trend with increasing temperature, while the dry weight remaining and cellulose content showed a decreasing trend (Table 3). The activities of sucrase and urease showed an increasing trend with increasing temperature, while the activity of peroxidase decreased first, then increased, and then decreased, with the highest activity under the control condition and the lowest activity under 2.5 °C warming (Figure 6).
Table 3.
Dry weight remaining and chemical properties of litters after decomposition for 360 days.
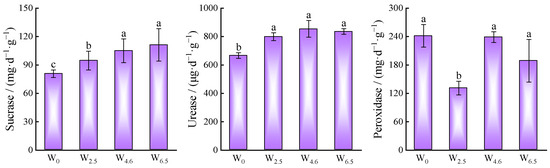
Figure 6.
Enzyme activities of litters under different temperature conditions. Different lowercase letters indicate significant differences between different treatments (p < 0.05).
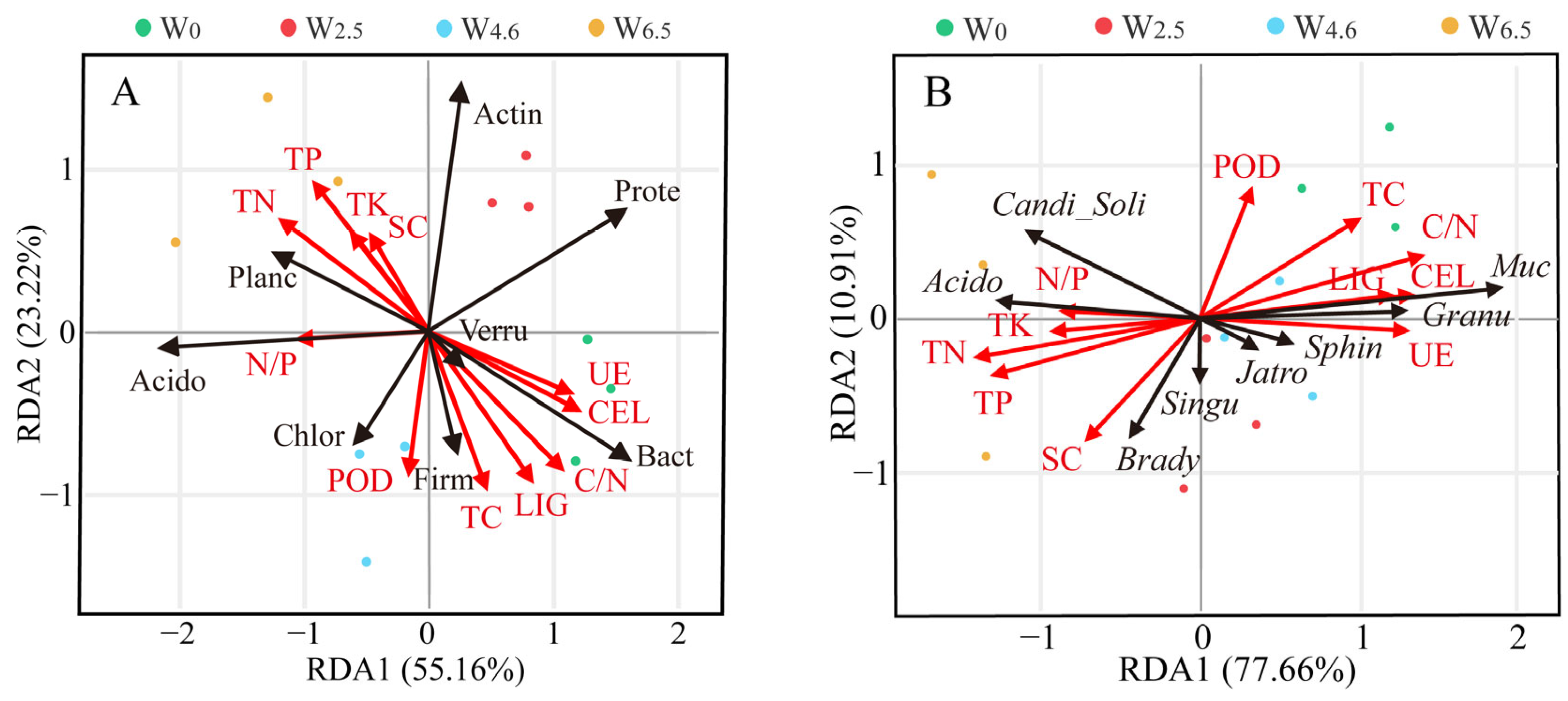
The redundancy analysis showed that the chemical properties and enzyme activities of the litter had significant effects on the relative abundance of the dominant bacterial communities (Figure 7). At the phylum level, axis-1 and axis-2 explained 78.38% of the variation in the relative abundance of bacteria under different temperature conditions. Some phyla had a strong association with the chemical properties of the litter, such as Planctomycetes, Bacteroidetes, and Verrucomicrobia. However, Proteobacteria showed no significant relationship with the chemical properties or enzyme activity factors. At the genus level, axis-1 and axis-2 explained 88.57% of the variation. Mucilaginibacter and Acidothermus had a strong association with lignin and cellulose content. Bradyrhizobium had a strong association with the activity of sucrase and peroxidase.
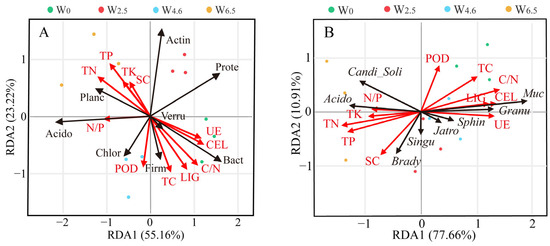
Figure 7.
Redundancy analysis relating phylum (A) and genus (B) to chemical properties and enzyme activity factors. The dots represent the samples in the same group, and the arrow lengths represent the relative significance, while the angle between the arrows represents the degree to which they are correlated. Prote: Proteobacteria, Actin: Actinobacteria, Planc: Planctomycetes, Acido: Acidobacteria, Bact: Bacteroidetes, Verru: Verrucomicrobia, Chlor: Chloroflexi, Firm: Firmicutes, Brady: Bradyrhizobium, Singu: Singulisphaera, Muc: Mucilaginibacter, Granu: Granulicella, Acido: Acidothermus, Sphin: Sphingomonas, Jatro: Jatrophihabitans, Candi_soli, Candidatus_solibacter, LIG: lignin, CEL: cellolose, POD: peroxidase, SC: sucrase, UE: urease.
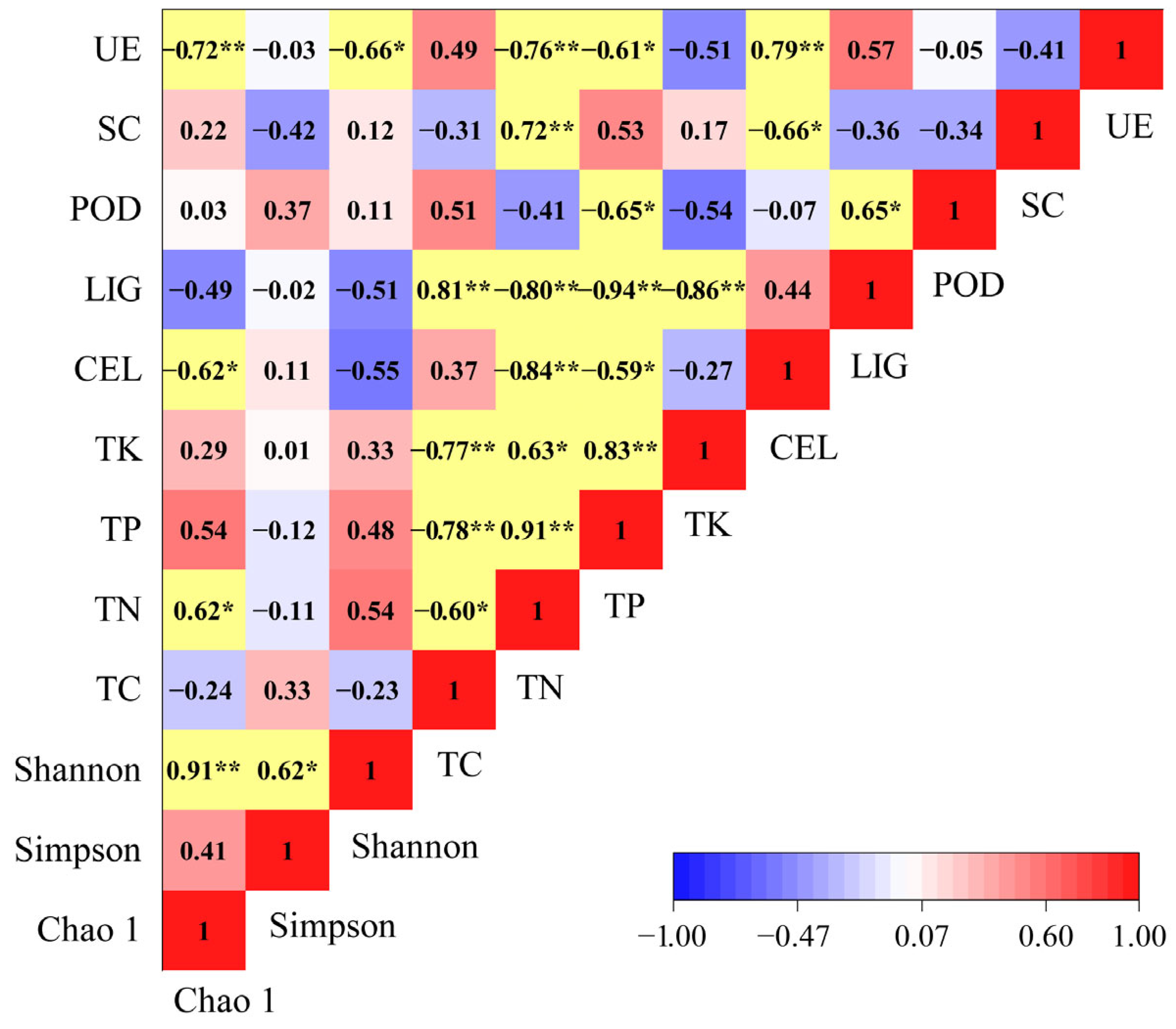
We found that the chemical properties and enzyme activities of the litter had significant effects on the Chao1, Simpson, and Shannon indices (Figure 8). The Chao1 index had a negative association with cellulose content and urease activity and a positive association with nitrogen content. The Shannon index was significantly negatively correlated with the urease activity.
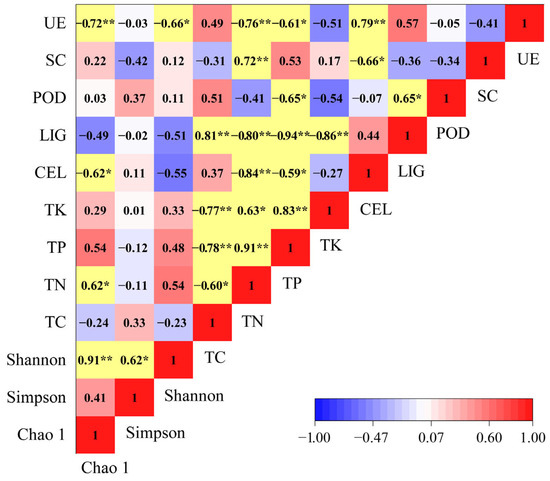
Figure 8.
Correlation between α-diversity indices, chemical properties, and enzyme activities factors. *: p < 0.05; **: p < 0.01. The yellow color indicates a significant correlation between each other.
3.5. Analysis of Importance of Warming on Bacterial Community, Enzyme Activity, and Chemical Properties of Litter
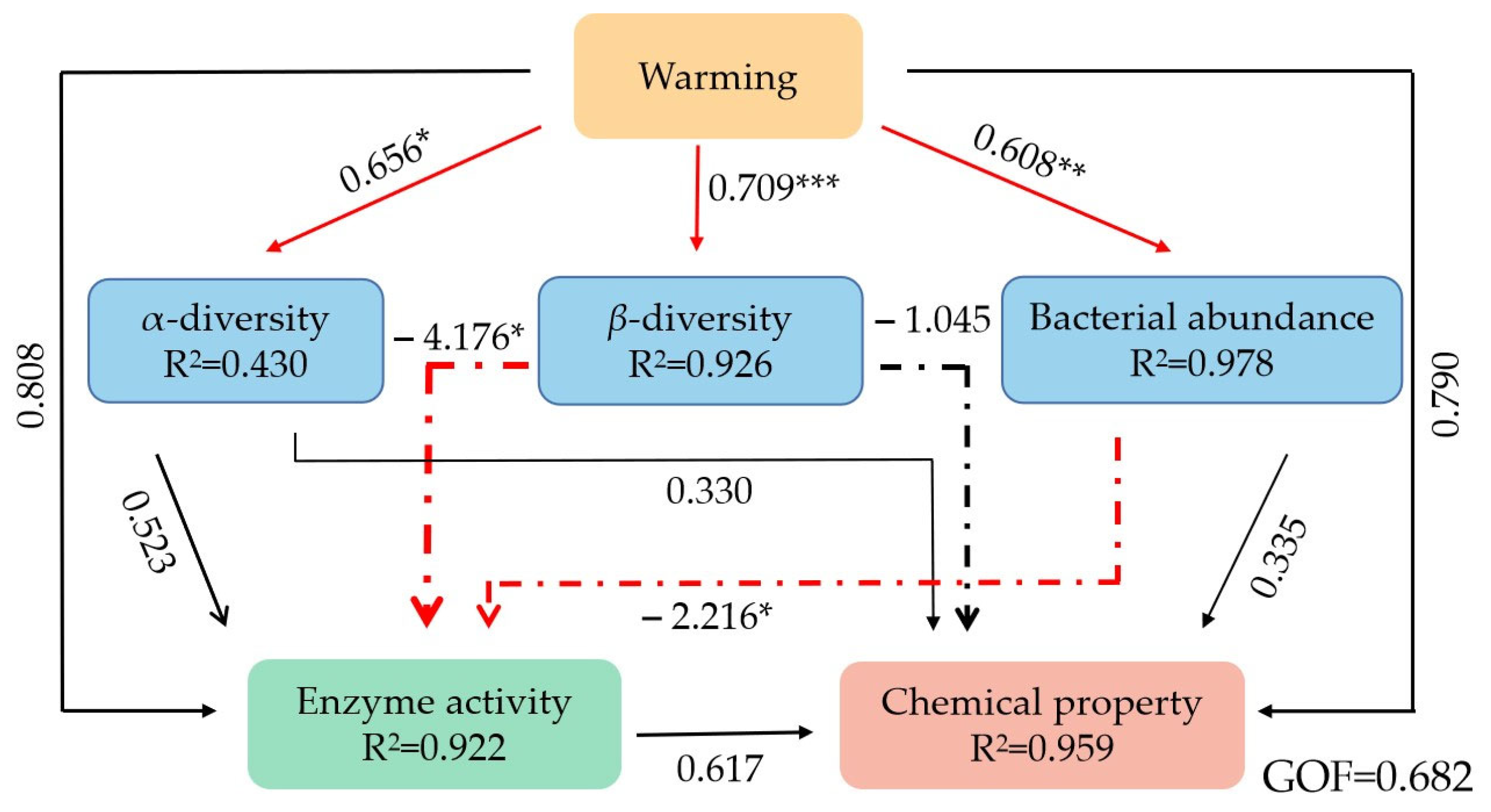
The PLS-PM was used to further explore the possible association of warming effects on the bacterial community structure, enzyme activity, and chemical properties of litter. Our model generated a goodness-of-fit value of 0.682, showing that warming had direct positive effects on α-diversity, β-diversity, and bacterial abundance. Similarly, β-diversity and bacterial abundance showed direct negative effects on enzyme activity (Figure 9).
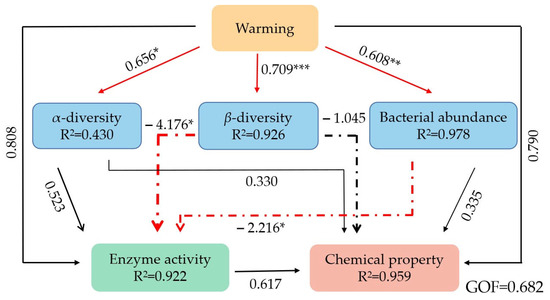
Figure 9.
Partial least-squares path models (PLS-PM) showing the effects of warming on bacterial community structure, enzyme activity, and chemical properties. *: p < 0.05; **: p < 0.01; ***: p < 0.001.
4. Discussion
4.1. Effects of Warming on Bacterial Community Structure in Forest Litters
Microbial communities can degrade nearly 90% of the organic components in litter. Studies have found that fungi secrete enzymes that catalyze the degradation of complex compounds during litter decomposition [28]. Compared to fungi, bacteria play a more important role in the degradation of keratin and aromatic compounds [29]. A dynamic equilibrium occurs during litter decomposition; warming causes changes in microbial community structure and affects the activity and ecological function of microorganisms, resulting in changes in the litter decomposition rate [2]. Song [30] simulated the effects of soil warming on bacterial communities in Chinese fir leaf litter by embedding resistance heating cables and found that Acidobacteria, Proteobacteria, and Actinobacteria were the most abundant phyla; soil warming significantly affects the relative abundance of Actinobacteria and Proteobacteria. After 360 days of decomposition, we found that Planctomycetes, Proteobacteria, Acidobacteria, and Actinobacteria were the dominant phyla in Chinese fir litters under different temperature conditions, and their relative abundance was more than 10%. A recent study has shown that Proteobacteria are crucial in the oxidation and energy metabolism of inorganic and organic compounds, whereas Actinobacteria are mainly involved in the degradation of complex carbohydrates, both of which are important saprophytic bacteria in the litter decomposition process [31]. What is noteworthiness is that warming had no significant effects on the abundance of Proteobacteria, Actinobacteria, and Acidobacteria, which may be due to their wide ecological ranges and strong adaptability to external temperature change [32]. Moreover, they might be abundant in the soil and transferred to the vegetal material in the litterbags. Soil quality is an important factor affecting the abundance of bacterial communities, and the availability of soil nutrients significantly affects the metabolic turnover of microorganisms [33].
Warming significantly decreased the relative abundance of Bacteroidetes, and that of Planctomycetes significantly increased under 6.5 °C warming conditions. The differences in the responses of bacterial communities to warming may have resulted from litter nutrients. Our results showed that the abundance of bacterial communities was strongly associated with the litter nutrient content. Bacteria as a heterotrophic organism, warming alters the content of organic compounds preferentially absorbed by the bacteria, causing changes in the abundance of bacterial communities. Previous studies have found that Planctomycetes are primarily responsible for the degradation of polycyclic aromatic compounds and promote the mineralization of organic carbon [34], whereas Bacteroidetes are mainly involved in nitrification and the degradation of complex biomacromolecules [35]. Interestingly, with the rising of temperature, the lignin content, cellulose content, and relative abundance of Bacteroidetes showed a decreasing trend, while nitrogen content showed an increasing trend. This indicates that Bacteroidetes may not be the main bacterial community involved in the degradation of structural carbohydrates in Chinese fir litter. The increase in nitrogen content may result from an increase in the species richness and diversity of bacterial communities. Nitrogen is essential for the growth and turnover of microbial communities as it participates in the construction of proteins [36]. At the genus level, warming decreased the relative abundance of Mucilaginibacter and Granulicella and increased that of Acidothermus. The abundance of Mucilaginibacter was closely related to lignin and cellulose contents, and the abundance of Acidothermus was related to N/P and K content (Figure 7B). In forest ecosystems, a variety of bacterial communities play important roles in nutrient transformation between litter and soil [37].
The bacterial community structure is highly sensitive to changes in temperature. Warming changes the composition and ecological function of the bacterial community and affects the decomposition process of litter. Xie et al. [2] simulated warming using a top-open chamber and found that warming increased the abundance and diversity of bacterial communities and promoted the degradation of carbon compounds. Zhao et al. [38] simulated the effect of warming on the microbial community during litter decomposition using 1000 lux light and found that warming significantly increased the α-diversity index of the bacterial community. In our study, we found that warming increased the species richness and diversity of bacterial communities in Chinese fir litter. The LEfSe analysis showed that the biomarkers were different under different temperature conditions (Figure 5), and the functional type of bacterial communities changed from fermentation and chemoheterotrophy to cellulolysis and oxygenic photoautotrophy (Figure 6). Zhou et al. [39] have indicated that warming was conducive to improving the utilization rate of organic compounds and the absorption efficiency of nutrients by bacterial communities. Additionally, warming is beneficial for enhancing the symbiotic relationships and information exchange between bacterial communities [40], strengthening the interactions between bacterial communities [41] and thus improving the litter decomposition rate. However, in the elevation gradient study, besides temperature among the climatic factors, humidity, and radiation may also be relative factors that make bacterial community structure vary with altitudes. Different precipitation and UV-B radiation intensities significantly affect the release rate of easily decomposed components in the litter and alter the composition of bacterial communities. Ren et al. [42] reported that increased precipitation significantly enhanced soil microbial biomass by 16.18%, which in turn accelerated the decomposition rate of forest litter. Different forest gaps along the altitude gradients significantly affect the microbial biomass in litter. Zhang et al. [43] found that a forest gap of 100–900 square meters was conducive to the increment of microbial biomass carbon and nitrogen during the decomposition of forest litter.
In recent years, there has been a significant decline in productivity caused by the continuous multi-generation planting of Chinese fir. Strengthening the exploration of the ecological function of the bacterial community in litter is beneficial for gaining insight into the nutrient return status in the forest ecosystem. Studies have found that bacterial and fungal communities are closely linked and form a complex symbiotic network. Warming changes the interaction patterns between them, enhancing the complexity and stability of the microbial network [44], and further affecting litter decomposition. Therefore, in the future, strengthening dynamic research on the interaction between bacterial and fungal communities will be more valuable for understanding the mechanisms of forest litter decomposition.
4.2. Effects of Warming on the Activity of Enzymes in Forest Litter Decomposition
The enzymes involved in forest litter decomposition act as catalysts in the process of litter degradation, and their activity determines the decomposition rate. Studies have reported that the activity of enzymes involved in litter decomposition is closely related to temperature and increases with increasing temperature within a certain range [45]. Xu et al. [46] found that the influence of temperature on enzyme activity can be either direct or indirect. On one hand, temperature directly changes enzyme activity by affecting enzyme kinetics. On the other hand, temperature indirectly changes enzyme activity by affecting the microbial community structure and nutrient mineralization rate. In our study, we found that the activities of sucrase and urease in the litter increased significantly under warming conditions. Under high temperatures, the microbial community has high activity and a fast turnover rate, which accelerates the nutrient conversion efficiency of litter and provides a sufficient nutrient supply for the synthesis of microbial extracellular enzymes [47]. Meng et al. [48] found that the response of enzyme activity to warming was closely related to the bacterial quantity. Warming results in an increase in the abundance and diversity of bacterial communities in the litter, which promotes the synthesis of more enzymes.
Peroxidase is an important enzyme that degrades lignin and keratin during litter decomposition. Our results showed that the response of peroxidase activity to warming did not show a consistent increase but showed a fluctuating trend, with the highest activity under control conditions, similar to the results of Chen et al. [49]. This may have been caused by the effect of warming on the growth of white rot fungi that secrete lignin enzymes by reducing the content of available carbon in the litter. Tan et al. [50] found that peroxidase activity might have a stronger adaptability to low temperatures, whereas warming inhibits its activity. In addition, the differences between the altitudes at which the litterbags were located may have contributed to the decrease in peroxidase activity. Studies have reported that enzyme activity was extremely sensitive to temperature and humidity in the external environment [51], and changes in temperature, humidity, and understory light environment at different altitudes had an impact on the activity of enzymes in litter decomposition.
5. Conclusions
Warming significantly affected the bacterial community structure in Chinese fir litter. Our findings showed that warming significantly increased the abundance and diversity of bacterial communities in Chinese fir litter and accelerated the dry weight loss of litters. The abundance and diversity of bacterial communities are closely related to the chemical properties and enzymatic activities of the litter. Climate warming will help enhance information exchange and symbiotic relationships between bacterial communities, promote the decomposition rate of litter, and accelerate nutrient cycling in Chinese fir plantation ecosystems. However, because of the wide distribution area and large span of climatic zone, there is diversity in the morphology and physiology of Chinese fir. In the future, the effects of warming on carbon sink function and nutrient cycling in Chinese fir plantation ecosystems with different geographical provenances still need to be further investigated.
Author Contributions
Conceptualization, X.W. and X.M.; methodology, X.W., J.L., Z.H. and Y.J.; formal analysis, X.W., J.L., Z.H. and J.Z.; investigation, X.W., Y.J. and J.Z.; writing—original draft preparation, X.W.; writing—review and editing, P.W. and X.M.; project administration, X.M.; funding acquisition, X.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2021YFD2201302), the Key Research and Development Topic of China (2023YFF130440502), and the Scientific Research Foundation of the Graduate School of Fujian Agriculture and Forestry University (324-1122YB071).
Data Availability Statement
The data presented in this study are available upon request from the corresponding author. The data are not publicly available due to privacy concerns but may be provided by the corresponding author on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, M.; Liu, G.; Xing, Y.; Yan, G.; Wang, Q. Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest. Forests 2024, 15, 372. [Google Scholar] [CrossRef]
- Xie, P.; Liu, G.; Sun, J.; Guo, C.; Zhu, K.; Zhang, X.; Wang, L. Effects of Simulated warming on litter decomposition and bacterial communities of wetland plants. Acta Ecol. Sin. 2023, 43, 10308–10319. [Google Scholar]
- Zhou, T.J. New physical science behind climate change: What does IPCC AR6 tell us? Innovation 2021, 2, 100173. [Google Scholar] [CrossRef] [PubMed]
- Kolomyts, E.G. Predictive modelling of boreal forest resources in regulation of the carbon cycle and mitigation of the global warming. Int. J. Glob. Warm. 2022, 27, 333–364. [Google Scholar] [CrossRef]
- Park, H.J.; Lim, S.S.; Yang, H.I.; Lee, K.S.; Kwak, J.H.; Park, S.I.; Kim, H.Y.; Lee, S.M.; Choi, W.J. Nitrogen effects on quantity, chemistry, and decomposability of Pinus densiflora and Quercus variabilis litters under elevated CO2 and warming. For. Ecol. Manag. 2020, 473, 118315. [Google Scholar] [CrossRef]
- Verónica, F.; Eric, C. Future increase in temperature more than decrease in litter quality can affect microbial litter decomposition in streams. Oecologia 2011, 167, 279–291. [Google Scholar]
- Liu, X.; Chen, S.; Li, X.; Yang, Z.; Xiong, D.; Xu, C.; Wanek, W.; Yang, Y. Soil warming delays leaf litter decomposition but exerts no effect on litter nutrient release in a subtropical natural forest over 450 days. Geoderma 2022, 427, 116139. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Xie, P.; Guo, C.; Zhu, K.; Tian, K. Climate warming enhances microbial network complexity by increasing bacterial diversity and fungal interaction strength in litter decomposition. Sci. Total Environ. 2023, 908, 168444. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, J.; Jia, H.; Lyu, L.; Langzhen, J.; Zhang, Q.B. Changes of growth-climate relationships of Smith fir forests along an altitudinal gradient. J. For. Res. 2024, 35, 76. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Roman, C.R.; Huaman, J.; Salinas, D.; Huaman, V.; Gibaja, A.; Mamani, M.; et al. The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New Phytol. 2011, 189, 967–977. [Google Scholar] [CrossRef]
- Klimek, B.; Niklińska, M. Changes in temperature sensitivity of forest litter during decomposition along an altitudinal gradient in temperate mountains—A reciprocal litter transplantation study. Catena 2024, 240, 107977. [Google Scholar] [CrossRef]
- Bohara, M.; Acharya, K.; Perveen, S.; Manevski, K.; Hu, C.; Yadav, R.K.P.; Shrestha, K.; Li, X. In situ litter decomposition and nutrient release from forest trees along an elevation gradient in Central Himalaya. Catena 2020, 194, 104698. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, X.; Zhong, A.; Guo, S.; Zhang, H. Variations in microbial residue and its contribution to SOC between organic and mineral soil layers along an altitude gradient in the Wuyi Mountains. Forests 2023, 14, 1678. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Q.; An, B.; Wu, X.; Sun, L.; Wu, P.; Liu, B.; Ma, X. Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings. Forests 2021, 12, 853. [Google Scholar] [CrossRef]
- Xia, L.; Yu, J.; Deng, L.; Li, X.; Zhou, C.; Xu, Y. Researches on Soil Decline of Chinese Fir Plantation. World For. Res. 2018, 31, 37–42. [Google Scholar]
- Sheng, W. On the maintenance of long-term productivity of plantation in China. For. Res. 2018, 31, 1–14. [Google Scholar]
- Wang, X.; Yang, X.; Yang, N.; Xin, X.; Qu, Y.; Zhao, N.; Gao, Y. Effects of litter diversity and composition on litter decomposition characteristics and soil microbial community. Acta Ecol. Sin. 2019, 39, 6264–6272. [Google Scholar]
- Bao, Y.; Dolfing, J.; Li, X.; Chen, R.; Cui, X.; Li, Z.; Lin, X.; Feng, Y. Bacterial interference competition and environmental filtering reduce the fungal taxonomic and functional contribution to plant residue decomposition in anoxic paddy soils. Soil Tillage Res. 2024, 236, 105938. [Google Scholar] [CrossRef]
- Chen, Y.; Lie, Z.; Liu, X.; Li, X.; Wu, T.; Chu, G.; Meng, Z.; Liu, J. The effect of warming on the activity of decomposition enzymes in litter of mixed coniferous and broad-leaved forests in South Asia. Chin. J. Appl. Environ. Biol. 2021, 27, 923–929. [Google Scholar]
- Van Meeteren, M.J.M.; Tietema, A.; van Loon, E.E.; Verstraten, J.M. Microbial dynamics and litter decomposition under a changed climate in a Dutch heathland. Appl. Soil Ecol. 2007, 38, 119–127. [Google Scholar] [CrossRef]
- Rinnan, R.; Stark, S.; Tolvanen, A. Responses of Vegetation and Soil Microbial Communities to Warming and Simulated Herbivory in a Subarctic Heath. J. Ecol. 2009, 97, 788–800. [Google Scholar] [CrossRef]
- Mao, R.; Zhang, X.; Song, C.; Wang, X.; Finnegan, P.M. Plant functional group controls litter decomposition rate and its temperature sensitivity: An incubation experiment on litters from a boreal peatland in northeast China. Sci. Total Environ. 2018, 626, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cao, Y.; Jiang, Y.; Chen, M.; Zhang, H.; Wu, P.; Ma, X. Dynamics of Non-Structural Carbohydrates Release in Chinese Fir Topsoil and Canopy Litter at Different Altitudes. Plants 2023, 12, 729. [Google Scholar] [CrossRef]
- Yan, Y.; Li, B.; Huang, Z.; Zhang, H.; Wu, X.; Farooq, T.H.; Wu, P.; Li, M.; Ma, X. Characteristics and Driving Factors of Rhizosphere Bacterial Communities of Chinese Fir Provenances. Forests 2021, 12, 1362. [Google Scholar] [CrossRef]
- Johannes, E.F.; André, N.; Konrad, K.; Lars, N.; Marcel, H. Rapid, real-time sucrase characterization: Showcasing the feasibility of a one-pot activity assay. J. Biotechnol. 2022, 354, 21–33. [Google Scholar]
- Jin, K.; Sleutel, S.; Buchan, D.; Never, S.D.; Cai, D.; Gabriels, D.; Jin, J. Changes of soil enzyme activities under different tillage practices in the Chinese Loess Plateau. Soil Tillage Res. 2009, 104, 115–120. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 173–174, 330–338. [Google Scholar] [CrossRef]
- Contos, P.; Murphy, N.P.; Kayll, Z.J.; Morgan, T.; Vido, J.J.; Decker, O.; Gibb, H. Rewilding soil and litter invertebrates and fungi increases decomposition rates and alters detritivore communities. Ecol. Evol. 2024, 14, e11128. [Google Scholar] [CrossRef] [PubMed]
- Witoon, P.; Tesfaye, W.; Guillaume, L.; Michael, S.; Marek, J.P.; Danuta, K.; Martin, H.; Dirk, K.; François, B. Life in leaf litter: Novel insights into community dynamics of bacteria and fungi during litter decomposition. Mol. Ecol. 2016, 25, 4059–4074. [Google Scholar]
- Song, H.W. Effects of Soil Warming on Litter Decomposition in Subtropical Forests and Its Microbial Mechanisms. Master’s Thesis, Fujian Normal University, Fuzhou, China, 2021. [Google Scholar]
- Kim, H.S.; Lee, S.H.; Young, J.H.; Finneran, K.T.; Kwon, M.J. Diversity and composition of soil Acidobacteria and Proteobacteria communities as a bacterial indicator of past land-use change from forest to farmland. Sci. Total Environ. 2021, 797, 148944. [Google Scholar] [CrossRef]
- Jiang, X.; Ma, D.; Zang, S.; Zhang, D.; Sun, H. Characteristics of soil bacterial and fungal community of typical forest in the Greater Khingan Mountains based on high-throughput sequencing. Microbiol. China 2021, 48, 1093–1105. [Google Scholar]
- Bradford, M.A.; Keiser, A.D.; Davies, C.A.; Mersmann, C.A.; Strickland, M.S. Empirical evidence that soil carbon formation from plant inputs is positively related to microbial growth. Biogeochemistry 2013, 113, 271–281. [Google Scholar] [CrossRef]
- Zheng, R.; Wang, C.; Liu, R.; Cai, R.; Sun, C. Physiological and metabolic insights into the first cultured anaerobic representative of deep-sea Planctomycetes bacteria. eLife 2024, 12, 89874. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, Z.M.; Yu, Z.D.; Zhu, L. Optimization of F/M ratio for stability of aerobic granular process via quantitative sludge discharge. Bioresour. Technol. 2018, 252, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhou, J.; Lin, K.; Zhang, Q.; Yuan, P.; Zeng, X.; Su, Y.; Xu, J.; Chen, Y.; Yang, Y. Effect of different altitudes on soil microbial biomass and community structure of Pinus taiwanensis forest in mid-subtropical zone. Acta Ecol. Sin. 2019, 39, 2215–2225. [Google Scholar]
- Yu, F.; Liang, J.; Shi, J.; Wang, S.; Lu, J. Effects of canopy damaged on the soil CO2 fixation bacterial community structure in Xiaokeng Forest Farm. Microbiol. China 2017, 44, 2297–2306. [Google Scholar]
- Zhao, B.; Xing, P.; Wu, Q. Interactions between bacteria and fungi in macrophyte leaf litter decomposition. Environ. Microbiol. 2020, 23, 1130–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chen, C.; Wang, Y.; Xu, Z.; Han, H.; Li, L.; Wan, S. Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland. Sci. Total Environ. 2013, 444, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sun, J.; Xie, P.; Guo, C.; Li, M.; Tian, K. Mechanism of bacterial communities regulating litter decomposition under climate warming in temperate wetlands. Environ. Sci. Pollut. Res. Int. 2023, 30, 60663–60677. [Google Scholar] [CrossRef] [PubMed]
- Meena, C.; Hens, C.; Acharyya, S.; Haber, S.; Boccaletti, S.; Barzel, B. Emergent stability in complex network dynamics. Nat. Phys. 2023, 19, 1033–1042. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Shi, Z.; Chen, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation. Soil Biol. Biochem. 2017, 115, 1–10. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, L.; Zhang, J.; Yuan, W.; Liu, H.; Li, X.; Zhang, Y. Dynamics of microbial biomass carbon and nitrogen during foliar litter decomposition under artificial forest gap in Pinus massoniana plantation. Chin. J. Appl. Ecol. 2016, 27, 672–680. [Google Scholar]
- Chen, M.; Zhu, X.; Zhao, C.; Yu, P.; Abulaizi, M.; Jia, H. Rapid microbial community evolution in initial Carex litter decomposition stages in Bayinbuluk alpine wetland during the freeze–thaw period. Ecol. Indic. 2021, 121, 107180. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Treseder, K.K. Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition. (Special Issue: Enzymes in biogeochemical cycles: Integrating experimental data, theory, and models). Biogeochemistry 2014, 117, 23–37. [Google Scholar] [CrossRef]
- Xu, Z.; Tang, Z.; Wang, C.; Xiong, P.; Cao, G.; Liu, Q. Effects of simulated warming on soil enzyme activities in two subalpine coniferous forests in west Sichuan. Chin. J. Appl. Ecol. 2010, 21, 2727–2733. [Google Scholar]
- Sardans, J.; Peñuelas, J.; Estiarte, M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Appl. Soil Ecol. 2007, 39, 223–235. [Google Scholar] [CrossRef]
- Meng, C.; Tian, D.; Zeng, H.; Li, Z.; Chen, H.; Niu, S. Global meta-analysis on the responses of soil extracellular enzyme activities to warming. Sci. Total Environ. 2020, 705, 135992. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, G.; Yang, Y.; Yang, Y. The response of soil surface enzyme activities to short time warming and litter decomposition in mountain forest. Acta Ecol. Sin. 2015, 35, 7071–7079. [Google Scholar]
- Tan, B.; Wu, F.; Yang, W.; Yu, S.; Liu, L.; Wang, A.; Yang, Y. Activities Soil oxidoreductase and their response to seasonal freeze-thaw in the subalpine/alpine forests of western Sichuan. Acta Ecol. Sin. 2012, 32, 6670–6678. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Xing, B.S.; Dukes, J.S. Labile compounds in plant litter reduce the sensitivity of decomposition to warming and altered precipitation. New Phytol. 2013, 200, 122–133. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).