
Effects of UVA on Flavonol Accumulation in Ginkgo biloba
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
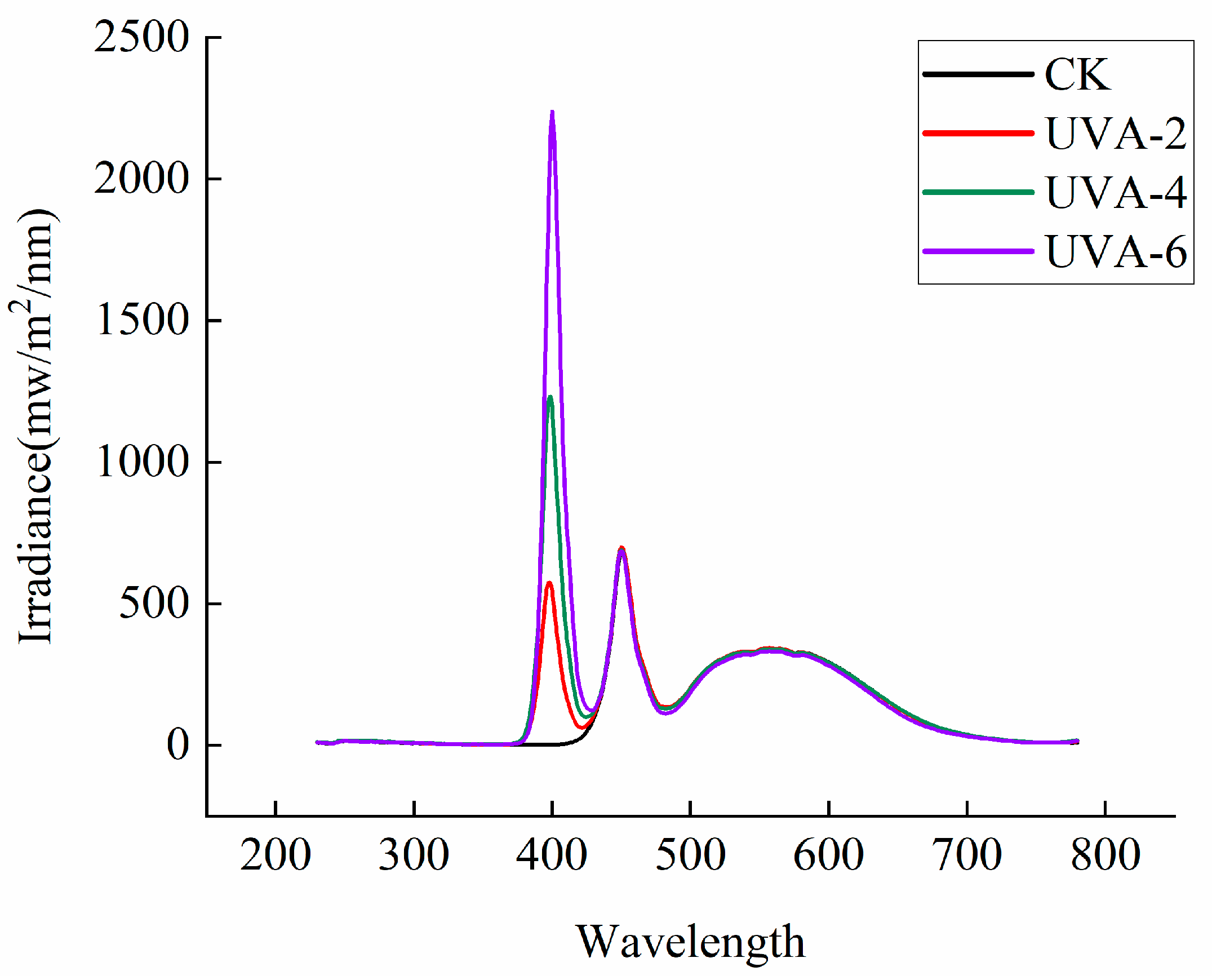
2.2. Experimental Design
2.3. Test Method
2.3.1. Determination of the Metabolic Enzyme Activity of Phenylpropane
2.3.2. Determination of the Flavonol Content
2.3.3. RNA Extraction, Transcriptome Sequencing, and Library Construction
2.3.4. Real-Time Quantitative PCR (qRT-PCR)
3. Result Analysis
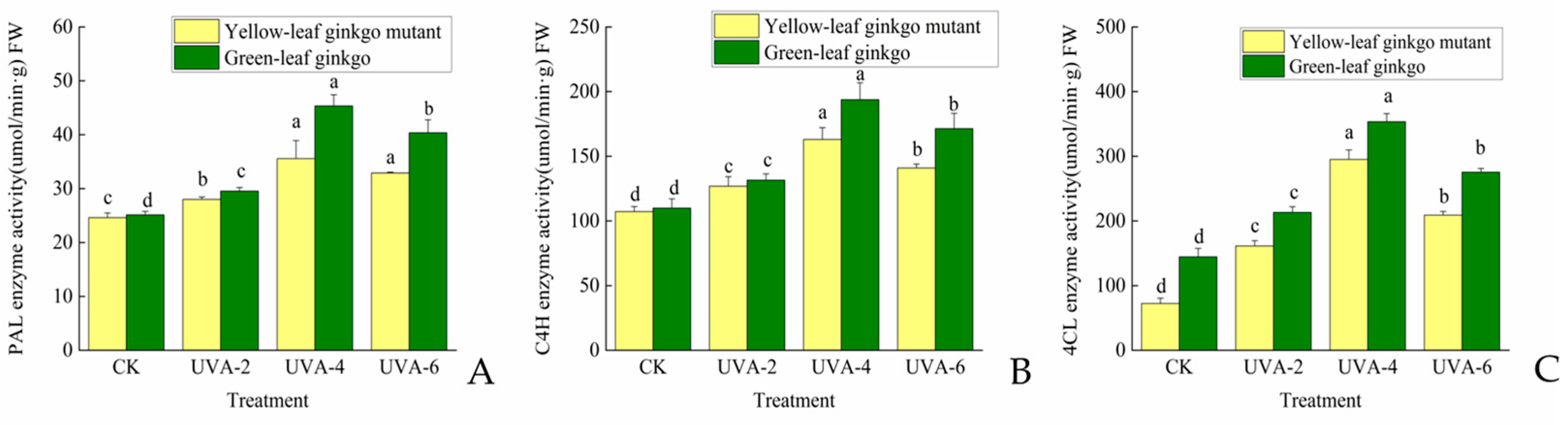
3.1. Effects of Different UVA Intensities on Phenylpropane Metabolizing Enzyme Activities in Ginkgo Hybrid Offspring
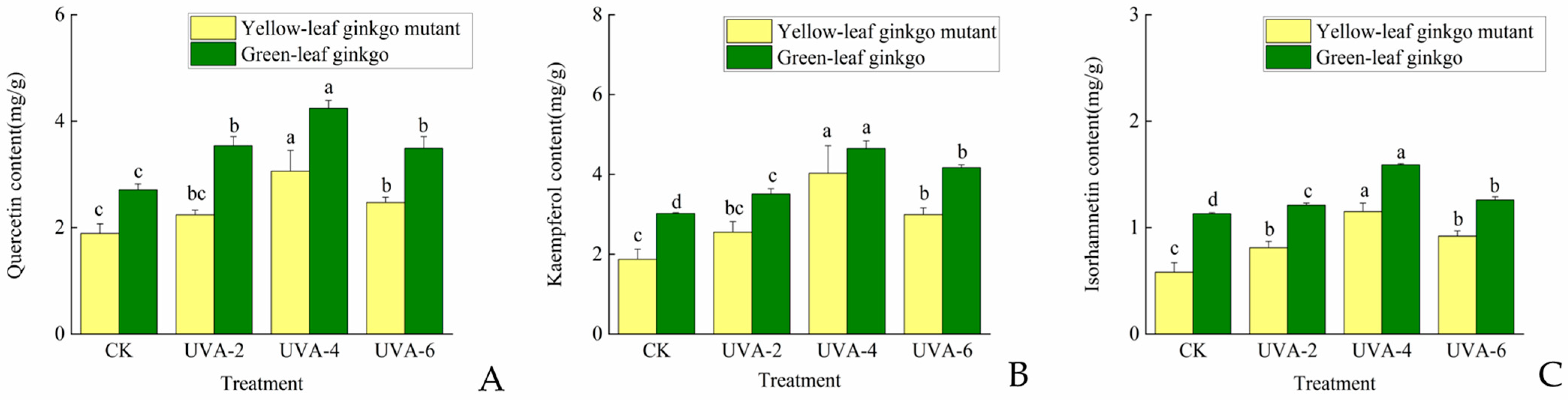
3.2. Effects of Different Intensities of UVA on Flavonol Content of G. biloba Hybrid Offspring
3.3. Transcriptome Sequencing Statistics and Quality Evaluation
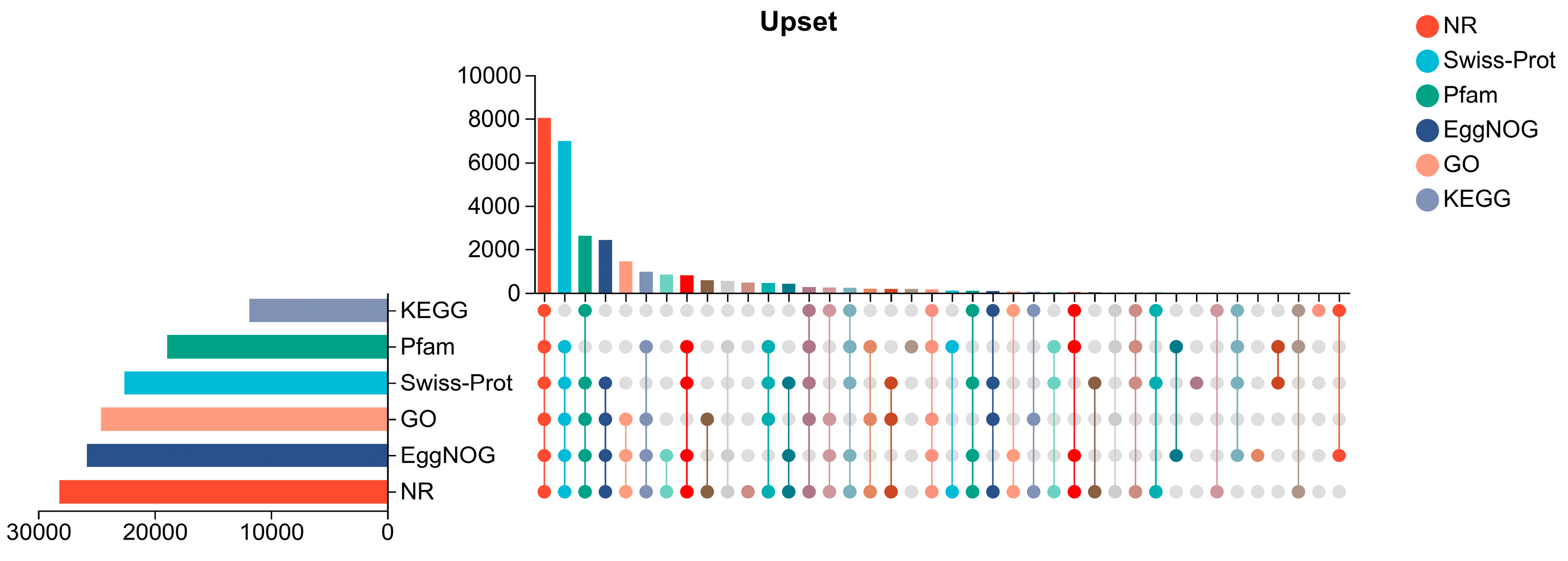
3.4. Functional Annotation of the Six Major Databases
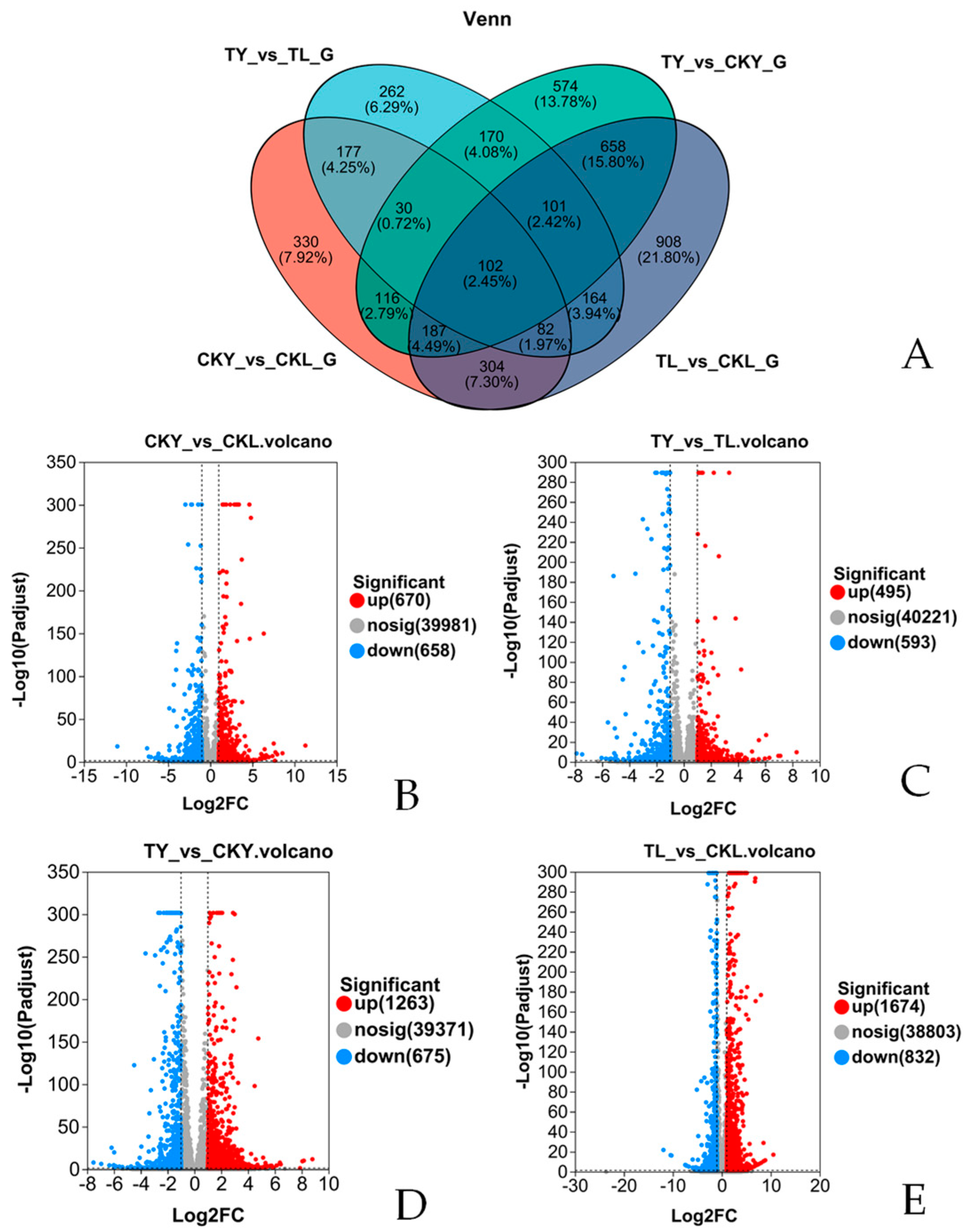
3.5. Differential Expression Analysis of Ginkgo Genes under Different Intensities of UVA Treatment
3.6. GO Annotation Analysis of DEGs
3.7. KEGG Enrichment Analysis of Differentially Expressed Genes
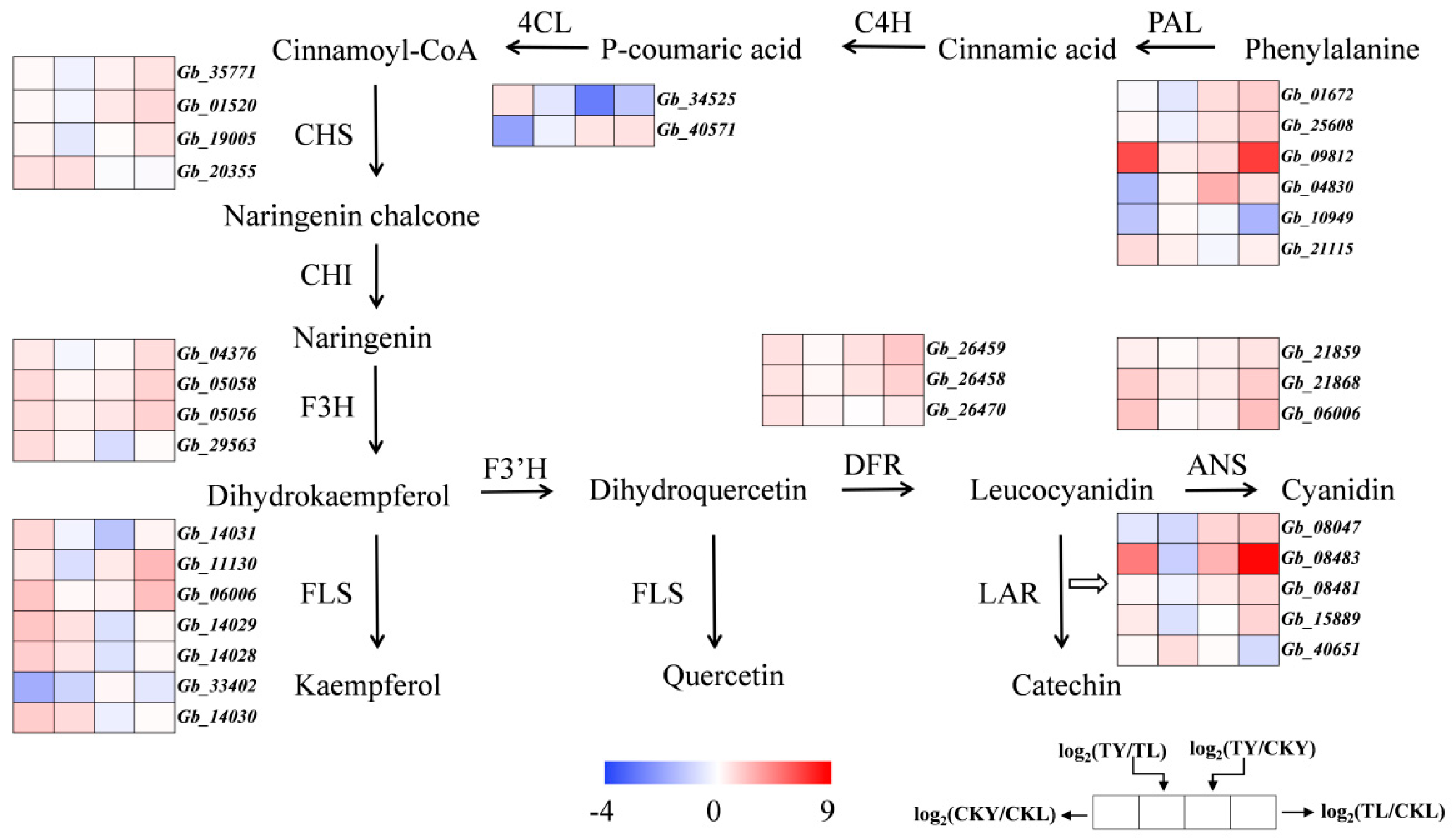
3.8. Differential Expression of Genes Related to Flavonoid Biosynthesis
3.9. Validation of qRT-PCR for Key Differential Enzyme Genes of the Ginkgo Flavonoid Synthesis Pathway
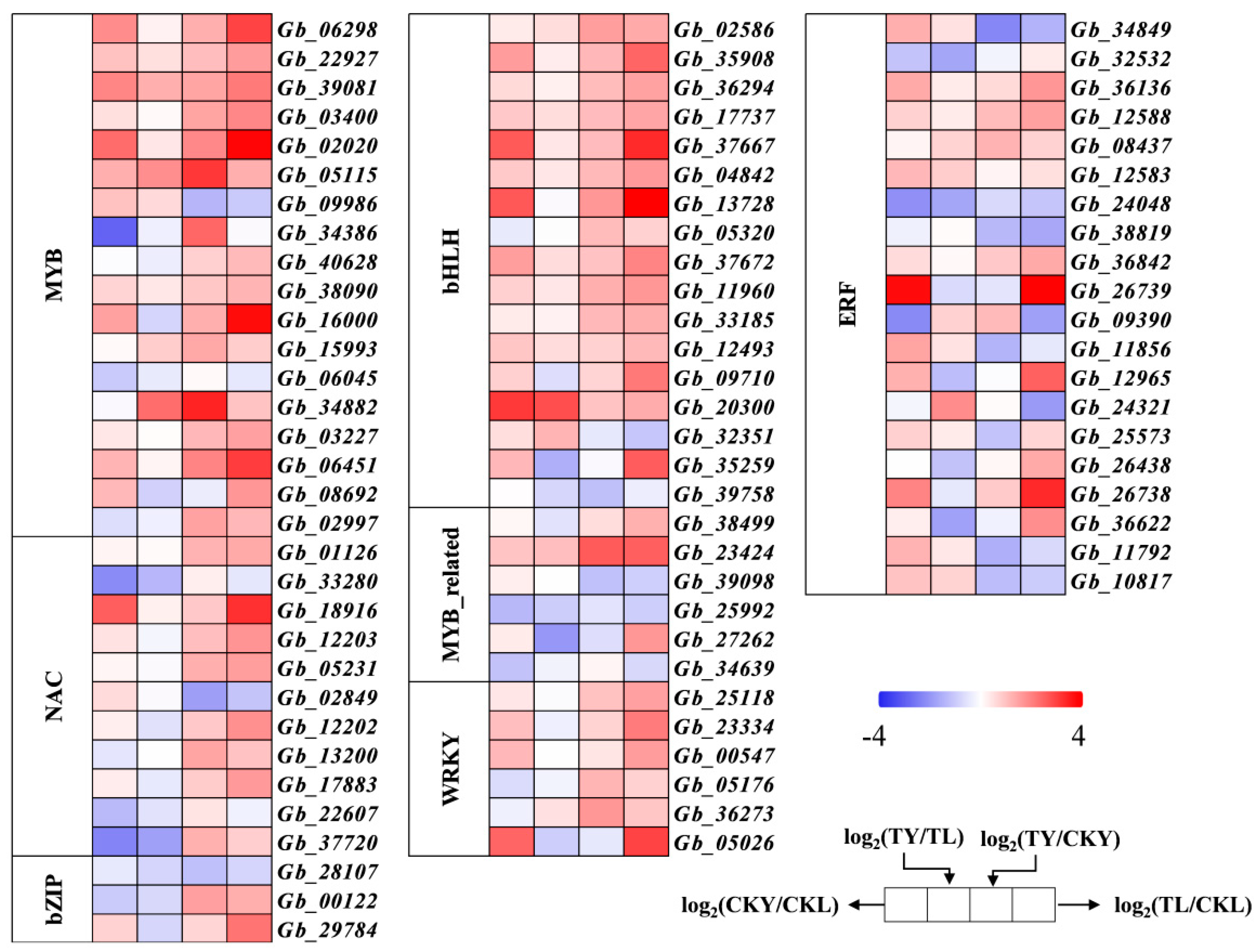
3.10. Differential Expression of Transcription Factors
3.11. Validation of qRT-PCR for Key Transcription Factors of Ginkgo Flavonoid Synthesis
4. Discussion
4.1. Effects of Different Intensities of UVA on the Enzyme Activity of the Phenylpropane Pathway in Hybrid Ginkgo Offspring
4.2. Effects of Different Intensities of UVA on Flavonols in Ginkgo Hybrid Offspring
4.3. Effects of Different Intensities of UVA on Enzyme Genes Related to Flavonoid Metabolism of Hybrid G. biloba
4.4. Analysis of the Transcriptional Regulation Mechanism of Ginkgo biloba Flavonoid Synthesis by Different Intensities of UVA
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smarda, P.; Vesely, P.; Smerda, J.; Bures, P.; Knápek, O.; Chytrá, M. Polyploidy in a “living fossil” Ginkgo biloba. New Phytol. 2016, 212, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Pan, S. Advance in study of ginkgolic acid contained in Ginkgo biloba preparations. China J. Chin. Mater. Medica 2012, 37, 274–277. [Google Scholar]
- Liu, X.; Yu, W.; Wang, G.; Cao, F.; Cai, J.; Wang, H. Comparative Proteomic and Physiological Analysis Reveals the Variation Mechanisms of Leaf Coloration and Carbon Fixation in a Xantha Mutant of Ginkgo biloba L. Int. J. Mol. Sci. 2016, 17, 1794. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-X.; Yang, S.-B.; Lu, Z.; He, Z.-C.; Ye, Y.-L.; Zhao, B.-B.; Wang, L.; Jin, B. Cytological, physiological, and transcriptomic analyses of golden leaf coloration in Ginkgo biloba L. Hortic. Res. 2018, 5, 32. [Google Scholar] [CrossRef]
- Sun, Y.; Bai, P.-P.; Gu, K.-J.; Yang, S.-Z.; Lin, H.-Y.; Shi, C.-G.; Zhao, Y.-P. Dynamic transcriptome and network-based analysis of yellow leaf mutant Ginkgo biloba. BMC Plant Biol. 2022, 22, 465. [Google Scholar] [CrossRef] [PubMed]
- Kressmann, S.; Biber, A.; Wonnemann, M.; Schug, B.; Blume, H.H.; Müller, W.E. Influence of pharmaceutical quality on the bioavailability of active components from Ginkgo biloba preparations. J. Pharm. Pharmacol. 2002, 54, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.T.; Lv, S.D.; Zhao, L.; Gao, T.; Yu, C.; Hu, J.N.; Ma, F. Advances in the study of the function and mechanism of the action of flavonoids in plants under environmental stresses. Planta 2023, 257, 108. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.X.; Hu, H.Q.; Wang, L.H.; Zhou, Q.; Huang, X.H. Enzymological mechanism for the regulation of lanthanum chloride on flavonoid synthesis of soybean seedlings under enhanced ultraviolet-B radiation. Environ. Sci. Pollut. Res. 2014, 21, 8792–8800. [Google Scholar] [CrossRef]
- Kanazawa, K.; Hashimoto, T.; Yoshida, S.; Sungwon, P.; Fukuda, S. Short Photoirradiation Induces Flavonoid Synthesis and Increases Its Production in Postharvest Vegetables. J. Agric. Food Chem. 2012, 60, 4359–4368. [Google Scholar] [CrossRef]
- Noda, N.; Kanno, Y.; Kato, N.; Kazuma, K.; Suzuki, M. Regulation of gene expression involved in flavonol and anthocyanin biosynthesis during petal development in lisianthus (Eustoma grandiflorum). Physiol. Plant. 2004, 122, 305–313. [Google Scholar] [CrossRef]
- Zhao, B.B.; Wang, L.; Pang, S.Y.; Jia, Z.C.; Wang, L.; Li, W.X.; Jin, B. UV-B promotes flavonoid synthesis in Ginkgo biloba leaves. Ind. Crop. Prod. 2020, 151, 112483. [Google Scholar] [CrossRef]
- Ni, J.; Hao, J.; Jiang, Z.F.; Zhan, X.R.; Dong, L.X.; Yang, X.L.; Sun, Z.H.; Xu, W.Y.; Wang, Z.K.; Xu, M.J. NaCl Induces Flavonoid Biosynthesis through a Putative Novel Pathway in Post-harvest Ginkgo Leaves. Front. Plant Sci. 2017, 8, 920. [Google Scholar] [CrossRef]
- Xu, W.J.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Premathilake, A.T.; Ni, J.B.; Bai, S.L.; Tao, R.Y.; Ahmad, M.; Teng, Y.W. R2R3-MYB transcription factor PpMYB17 positively regulates flavonoid biosynthesis in pear fruit. Planta 2020, 252, 59. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.L.; Li, W.D.; Zheng, Q.Y.; Wang, W.Q.; Xiao, B.; Xing, D. Effect of low light intensity on growth and accumulation of secondary metabolites in roots of Glycyrrhiza uralensis Fisch. Biochem. Syst. Ecol. 2010, 38, 160–168. [Google Scholar] [CrossRef]
- Sanchez-Campos, Y.; Cárcamo-Fincheira, P.; González-Villagra, J.; Jorquera-Fontena, E.; Acevedo, P.; Soto-Cerda, B.; Nunes-Nesi, A.; Inostroza-Blancheteau, C.; Tighe-Neira, R. Physiological and molecular effects of TiO2 nanoparticle application on UV-A radiation stress responses in Solanum lycopersicum L. Protoplasma 2023, 260, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Surjadinata, B.B.; Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. UVA, UVB and UVC Light Enhances the Biosynthesis of Phenolic Antioxidants in Fresh-Cut Carrot through a Synergistic Effect with Wounding. Molecules 2017, 22, 668. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, T.; Venäläinen, T.; Tegelberg, R.; Lindfors, A.; Julkunen-Tiitto, R.; Sutinen, S.; O’Hara, R.B.; Aphalo, P.J. Assessment of UV Biological Spectral Weighting Functions for Phenolic Metabolites and Growth Responses in Silver Birch Seedlings. Photochem. Photobiol. 2009, 85, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.W.; Tobler, M.A.; Keefover-Ring, K.; Flint, S.D.; Barkley, A.E.; Ryel, R.J.; Lindroth, R.L. Rapid modulation of ultraviolet shielding in plants is influenced by solar ultraviolet radiation and linked to alterations in flavonoids. Plant Cell Environ. 2016, 39, 222–230. [Google Scholar] [CrossRef]
- He, R.; Gao, M.F.; Li, Y.M.; Zhang, Y.T.; Song, S.W.; Su, W.; Liu, H.C. Supplemental UV-A Affects Growth and Antioxidants of Chinese Kale Baby-Leaves in Artificial Light Plant Factory. Horticulturae 2021, 7, 294. [Google Scholar] [CrossRef]
- Lim, Y.J.; Lyu, J.I.; Kwon, S.J.; Eom, S.H. Effects of UV-A radiation on organ-specific accumulation and gene expression of isoflavones and flavonols in soybean sprout. Food Chem. 2021, 339, 128080. [Google Scholar] [CrossRef] [PubMed]
- Son, Y. Effect of nitrogen fertilization on foliar nutrient dynamics of ginkgo seedlings. J. Plant Nutr. 2002, 25, 93–102. [Google Scholar] [CrossRef]
- Yu, W.W.; Liu, H.M.; Luo, J.Q.; Zhang, S.Q.; Xiang, P.; Wang, W.; Cai, J.F.; Lu, Z.G.; Zhou, Z.D.; Hu, J.J.; et al. Partial root-zone simulated drought induces greater flavonoid accumulation than full root-zone simulated water deficiency in the leaves of Ginkgo biloba. Environ. Exp. Bot. 2022, 201, 104998. [Google Scholar] [CrossRef]
- Wu, Z.F.; Wang, S.X.; Fu, Y.S.H.; Gong, Y.F. Spatial variation in the interaction between temperature and sunlight on ginkgo germination. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, G.B.; Cao, F.L.; Zhu, C.C.; Wang, G.Y.; El-Kassaby, Y.A. Light intensity affects the growth and flavonol biosynthesis of Ginkgo (Ginkgo biloba L.). New For. 2014, 45, 765–776. [Google Scholar] [CrossRef]
- Wang, G.B.; Cao, F.L.; Chang, L.; Guo, X.Q.; Wang, J. Temperature has more effects than soil moisture on biosynthesis of flavonoids in Ginkgo (Ginkgo biloba L.) leaves. New For. 2014, 45, 797–812. [Google Scholar] [CrossRef]
- Wang, G.P.; Zhang, L.; Wang, G.B.; Cao, F.L. Growth and flavonol accumulation of Ginkgo biloba leaves affected by red and blue light. Ind. Crop. Prod. 2022, 187, 115488. [Google Scholar] [CrossRef]
- Cheng, L.; Han, M.; Yang, L.M.; Yang, L.; Sun, Z.; Zhang, T. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crop. Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Zhao, Q.Y.; Ma, Y.; Huang, X.Q.; Song, L.J.; Li, N.; Qiao, M.W.; Li, T.E.; Hai, D.; Cheng, Y.X. GABA Application Enhances Drought Stress Tolerance in Wheat Seedlings (Triticum aestivum L.). Plants 2023, 12, 2495. [Google Scholar] [CrossRef]
- Lee, M.J.; Son, J.E.; Oh, M.M. Growth and phenolic content of sowthistle grown in a closed-type plant production system with a UV-A or UV-B lamp. Hortic. Environ. Biotechnol. 2013, 54, 492–500. [Google Scholar] [CrossRef]
- Miao, N.; Yun, C.; Shi, Y.T.; Gao, Y.; Wu, S.; Zhang, Z.H.; Han, S.L.; Wang, H.M.; Wang, W.J. Enhancement of flavonoid synthesis and antioxidant activity in Scutellaria baicalensis aerial parts by UV-A radiation. Ind. Crop. Prod. 2022, 187, 115532. [Google Scholar] [CrossRef]
- Kang, S.H.; Kim, J.E.; Zhen, S.Y.; Kim, J. Mild-Intensity UV-A Radiation Applied Over a Long Duration Can Improve the Growth and Phenolic Contents of Sweet Basil. Front. Plant Sci. 2022, 13, 858433. [Google Scholar] [CrossRef] [PubMed]
- Julkunen-Tiitto, R.; Häggman, H.; Aphalo, P.J.; Lavola, A.; Tegelberg, R.; Veteli, T. Growth and defense in deciduous trees and shrubs under UV-B. Environ. Pollut. 2005, 137, 404–414. [Google Scholar] [CrossRef]
- Fu, B.; Ji, X.M.; Zhao, M.Q.; He, F.; Wang, X.L.; Wang, Y.D.; Liu, P.F.; Niu, L. The influence of light quality on the accumulation of flavonoids in tobacco (Nicotiana tabacum L.) leaves. J. Photochem. Photobiol. B-Biol. 2016, 162, 544–549. [Google Scholar] [CrossRef]
- Chen, Y.C.; Li, T.; Yang, Q.C.; Zhang, Y.T.; Zou, J.; Bian, Z.H.; Wen, X.Z. UVA Radiation Is Beneficial for Yield and Quality of Indoor Cultivated Lettuce. Front. Plant Sci. 2019, 10, 1563. [Google Scholar] [CrossRef]
- Li, Y.M.; Zheng, Y.J.; Zheng, D.Q.; Zhang, Y.T.; Song, S.W.; Su, W.; Liu, H.C. Effects of Supplementary Blue and UV-A LED Lights on Morphology and Phytochemicals of Brassicaceae Baby-Leaves. Molecules 2020, 25, 5678. [Google Scholar] [CrossRef]
- Miao, N.; Yun, C.; Han, S.L.; Shi, Y.T.; Gao, Y.; Wu, S.; Zhao, Z.W.; Wang, H.M.; Wang, W.J. Postharvest UV-A radiation affects flavonoid content, composition, and bioactivity of Scutellaria baicalensis root. Postharvest Biol. Technol. 2022, 189, 111933. [Google Scholar] [CrossRef]
- Hu, T.; Gao, Z.Q.; Hou, J.M.; Tian, S.K.; Zhang, Z.X.; Yang, L.; Liu, Y. Identification of biosynthetic pathways involved in flavonoid production in licorice by RNA-seq based transcriptome analysis. Plant Growth Regul. 2020, 92, 15–28. [Google Scholar] [CrossRef]
- Lee, J.H.; Oh, M.M.; Son, K.H. Short-Term Ultraviolet (UV)-A Light-Emitting Diode (LED) Radiation Improves Biomass and Bioactive Compounds of Kale. Front. Plant Sci. 2019, 10, 1042. [Google Scholar] [CrossRef]
- Qian, M.J.; Kalbina, I.; Rosenqvist, E.; Jansen, M.A.K.; Strid, Å. Supplementary UV-A and UV-B radiation differentially regulate morphology in Ocimum basilicum. Photochem. Photobiol. Sci. 2023, 22, 2219–2230. [Google Scholar] [CrossRef]
- Li, W.; Tan, L.; Zou, Y.; Tan, X.; Huang, J.; Chen, W.; Tang, Q. The Effects of Ultraviolet A/B Treatments on Anthocyanin Accumulation and Gene Expression in Dark-Purple Tea Cultivar ‘Ziyan’ (Camellia sinensis). Molecules 2020, 25, 354. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Li, Y.Y.; She, G.B.; Zhang, X.C.; Jordan, B.; Chen, Q.; Zhao, J.; Wan, X.C. Metabolite profiling and transcriptomic analyses reveal an essential role of UVR8-mediated signal transduction pathway in regulating flavonoid biosynthesis in tea plants (Camellia sinensis) in response to shading. Bmc Plant Biol. 2018, 18, 233. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, N.; Luo, Z.; Zhao, X.; Yang, M.; Liu, Z.G.; Liu, M.J. Transcriptome-wide expression analysis of MYB gene family leads to functional characterization of flavonoid biosynthesis in fruit coloration of Ziziphus Mill. Front. Plant Sci. 2023, 14, 1171288. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Misra, P.; Trivedi, P.K. Constitutive expression of Arabidopsis MYB transcription factor, AtMYB11, in tobacco modulates flavonoid biosynthesis in favor of flavonol accumulation. Plant Cell Rep. 2015, 34, 1515–1528. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, C.; Wang, M.; Fu, F.-f.; El-Kassaby, Y.A.; Wang, T.; Wang, G. Metabolome and transcriptome analyses reveal flavonoids biosynthesis differences in Ginkgo biloba associated with environmental conditions. Ind. Crop. Prod. 2020, 158, 112963. [Google Scholar] [CrossRef]
- Xu, F.; Cheng, H.; Cai, R.; Li, L.L.; Chang, J.; Zhu, J.; Zhang, F.X.; Chen, L.J.; Wang, Y.; Cheng, S.H.; et al. Molecular Cloning and Function Analysis of an Anthocyanidin Synthase Gene from Ginkgo biloba, and Its Expression in Abiotic Stress Responses. Mol. Cells 2008, 26, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Xu, F.; Cheng, S.Y.; Liao, Y.L. Characterization and functional analysis of a MYB gene (GbMYBFL) related to flavonoid accumulation in Ginkgo biloba. Genes Genom. 2018, 40, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Li, L.L.; Zhang, W.W.; Cheng, H.; Sun, N.N.; Cheng, S.Y.; Wang, Y. Isolation, characterization, and function analysis of a flavonol synthase gene from Ginkgo biloba. Mol. Biol. Rep. 2012, 39, 2285–2296. [Google Scholar] [CrossRef]
- Lin, M.; Zhou, Z.Q.; Mei, Z.A. Integrative Analysis of Metabolome and Transcriptome Identifies Potential Genes Involved in the Flavonoid Biosynthesis in Entada phaseoloides Stem. Front. Plant Sci. 2022, 13, 792674. [Google Scholar] [CrossRef]
- Qian, M.J.; Wu, H.X.; Yang, C.K.; Zhu, W.C.; Shi, B.; Zheng, B.; Wang, S.B.; Zhou, K.B.; Gao, A.P. RNA-Seq reveals the key pathways and genes involved in the light-regulated flavonoids biosynthesis in mango (Mangifera indica L.) peel. Front. Plant Sci. 2023, 13, 1119384. [Google Scholar] [CrossRef]
- Xiang, L.L.; Liu, X.F.; Li, X.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. A Novel bHLH Transcription Factor Involved in Regulating Anthocyanin Biosynthesis in Chrysanthemums (Chrysanthemum morifolium Ramat.). PLoS ONE 2015, 10, e0143892. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, U.; Sagasser, M.; Mehrtens, F.; Stracke, R.; Weisshaar, B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol. Biol. 2005, 57, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC Transcription Factor, ANAC078, Regulates Flavonoid Biosynthesis under High-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer (5′-3′) | Reverse Primer (3′-5′) |
|---|---|---|
| GAPDH | ATCCACGGGAGTATTCAC | CTCATTCACGCCAACAAC |
| PAL (Gb_09812) | TCCTGACCTCGGCGTAGATTATGG | GGTGACTGGGTTTGCGAGATACTG |
| 4CL (Gb_40571) | AACAGAAGCGGATGAGAGCGAATG | TGTGAGTTAGCATGACGCCCTTTG |
| CHS (Gb_20355) | GCATGTGCCACCACTGGAGAAG | CGCTTCGCAAGACAACAGTTTCG |
| F3H (Gb_05058) | GGCGGCGTGCGAGGAATG | CTGGCGGGAGGGCAAAGAAATC |
| FLS (Gb_14030) | TGCCATCTCTCCCTCGCTCTTC | CATGCCAGTTTAGTGCCGTAGCC |
| DFR (Gb_26458) | GGCTGGTTATGCGTTTGCTTCAAC | TTCATCGTCCAAGTCGGCTTTCC |
| LAR (Gb_08481) | ATTGGTAATCGCAGCAGCAGAGTC | TGAGCGTACAAGAGCGTAAGTTGG |
| ANS (Gb_21859) | GTGCCTGGTCTCCAACTCTTCAAG | GCCCACTCTTGTATTTGCCATTGC |
| MYB (Gb_39081) | ATGGAGAATGGAAACACGGACTTG | ACCACGCCACTGCCTTGAG |
| MYB (Gb_06451) | AGCACAAGAAGCACGCACAAG | GATGGTAAGGCAGTTGGAGTGAAG |
| b ZIP (Gb_28107) | GCCAGCTTGTGCAGACTTTGAC | TTCAGCATTCGAGACCTCCCATC |
| bHLH (Gb_35908) | TCAGCAACAGATACAGTCACATTCC | AGCAGATTTGATGATCCACACTCAG |
| ERF (Gb_12588) | ATCGGCGGCGTCTGTAGC | TTGGGTCGTGCTTGATTCTTGAG |
| Sample | Raw Reads | Clean Reads | Total Mapped | Multiple Mapped | Uniquely Mapped | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| CKY_1 | 45,408,162 | 44,808,992 | 42,120,429 (94.0%) | 2,378,996 (5.31%) | 39,741,433 (88.69%) | 95.14 | 46.19 |
| CKY_2 | 54,468,294 | 53,771,538 | 50,289,981 (93.53%) | 2,810,629 (5.23%) | 47,479,352 (88.3%) | 94.94 | 46.25 |
| CKY_3 | 45,184,742 | 44,758,474 | 41,882,169 (93.57%) | 2,322,728 (5.19%) | 39,559,441 (88.38%) | 95.13 | 46.09 |
| CKL_1 | 49,435,520 | 48,592,064 | 45,742,011 (94.13%) | 2,952,929 (6.08%) | 42,789,082 (88.06%) | 95.09 | 46.31 |
| CKL_2 | 44,788,866 | 44,100,438 | 41,473,752 (94.04%) | 2,571,246 (5.83%) | 38,902,506 (88.21%) | 94.97 | 46.28 |
| CKL_3 | 46,647,470 | 46,141,812 | 43,506,464 (94.29%) | 2,767,065 (6.0%) | 40,739,399 (88.29%) | 95.72 | 46.12 |
| TY_1 | 41,700,364 | 41,246,156 | 38,580,392 (93.54%) | 2,113,162 (5.12%) | 36,467,230 (88.41%) | 95.83 | 45.81 |
| TY_2 | 52,013,850 | 51,369,770 | 48,034,026 (93.51%) | 2,681,649 (5.22%) | 45,352,377 (88.29%) | 95.76 | 45.88 |
| TY_3 | 54,527,510 | 53,960,036 | 50,488,323 (93.57%) | 2,847,049 (5.28%) | 47,641,274 (88.29%) | 95.76 | 45.9 |
| TL_1 | 46,585,016 | 46,059,222 | 42,780,240 (92.88%) | 2,588,474 (5.62%) | 40,191,766 (87.26%) | 95.67 | 45.87 |
| TL_2 | 60,566,890 | 59,865,396 | 55,681,034 (93.01%) | 3,426,978 (5.72%) | 52,254,056 (87.29%) | 95.96 | 45.85 |
| TL_3 | 61,495,202 | 60,944,886 | 56,738,281 (93.1%) | 3,509,698 (5.76%) | 53,228,583 (87.34%) | 95.96 | 45.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Wang, Z.; Wang, G.; Cao, F.; Yang, X.; Zhao, H.; Zhai, J. Effects of UVA on Flavonol Accumulation in Ginkgo biloba. Forests 2024, 15, 909. https://doi.org/10.3390/f15060909
Zhao Q, Wang Z, Wang G, Cao F, Yang X, Zhao H, Zhai J. Effects of UVA on Flavonol Accumulation in Ginkgo biloba. Forests. 2024; 15(6):909. https://doi.org/10.3390/f15060909
Chicago/Turabian StyleZhao, Qun, Zheng Wang, Gaiping Wang, Fuliang Cao, Xiaoming Yang, Huiqin Zhao, and Jinting Zhai. 2024. "Effects of UVA on Flavonol Accumulation in Ginkgo biloba" Forests 15, no. 6: 909. https://doi.org/10.3390/f15060909