
Introducing Native Tree Species Alter the Soil Organic Carbon, Nitrogen, Phosphorus, and Fine Roots in Moso Bamboo Plantations
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Soil and Fine Root Sampling
2.3. Extraction and Determination of Soil C, N and P
2.4. Determination of Soil Enzyme Activity and Microbial Abundance Measurements
2.5. Determination of Fine Root Characteristics
2.6. Statistical Analysis
3. Results
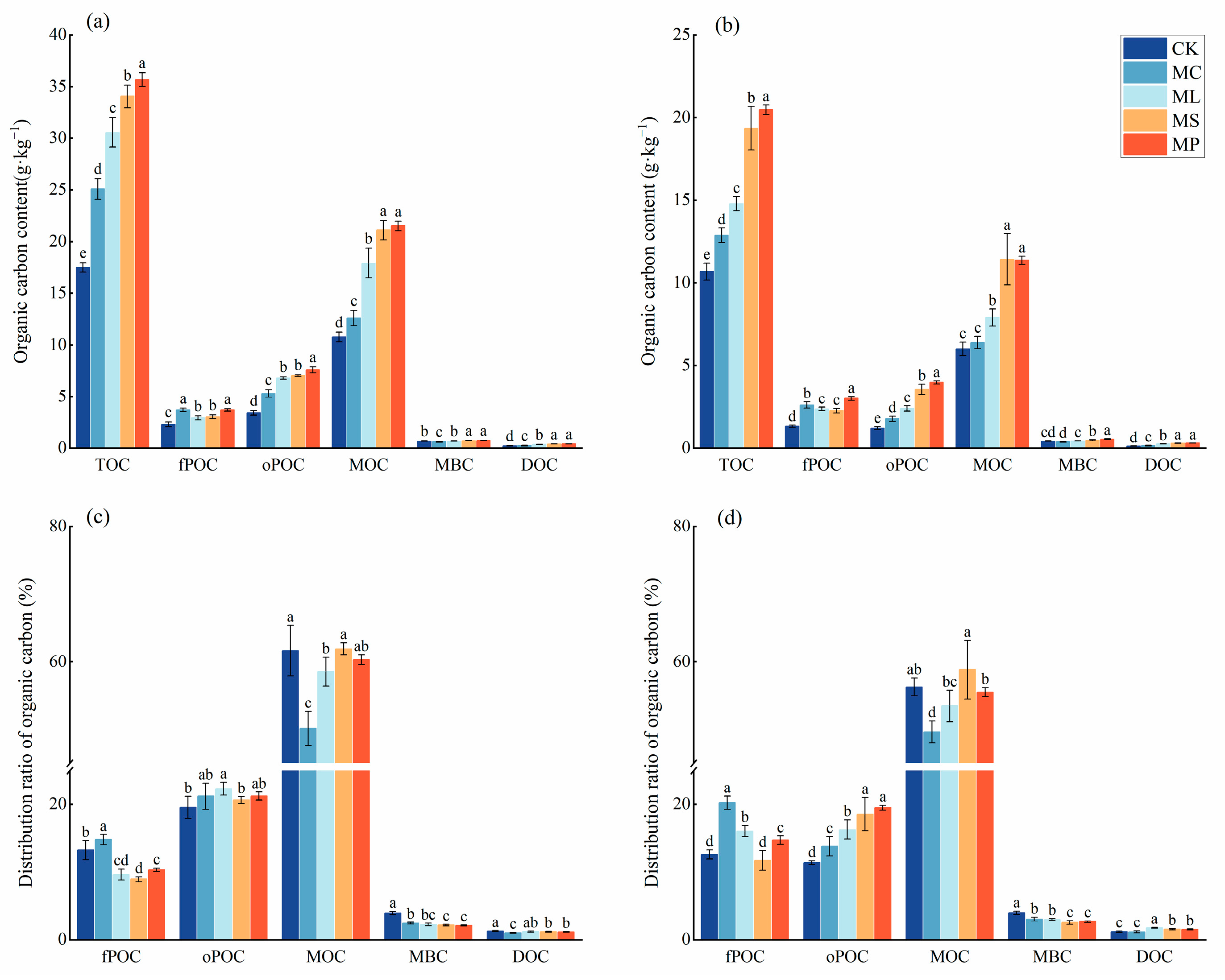
3.1. Soil C Fractions and Content
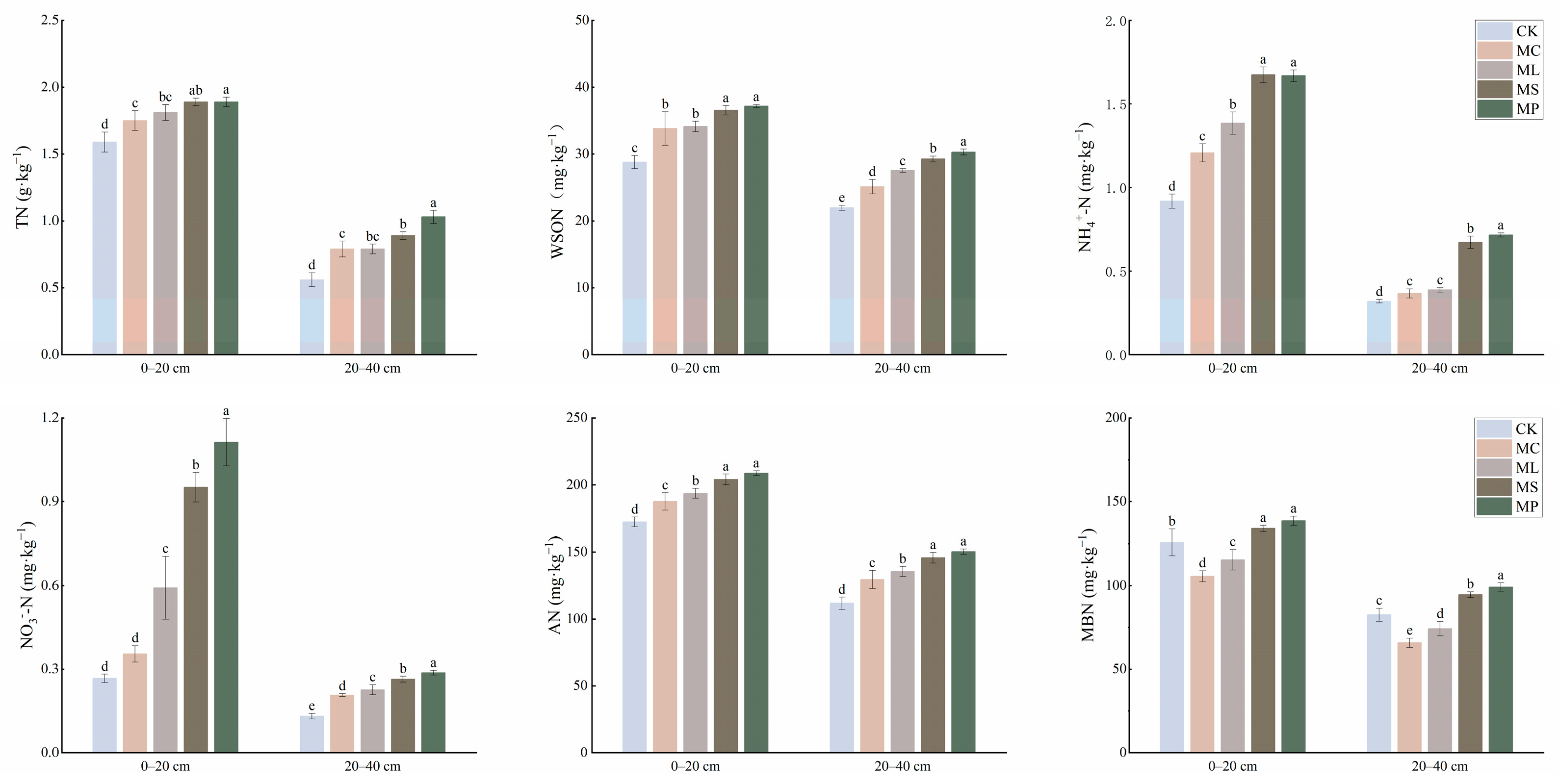
3.2. Soil N Fractions and Content
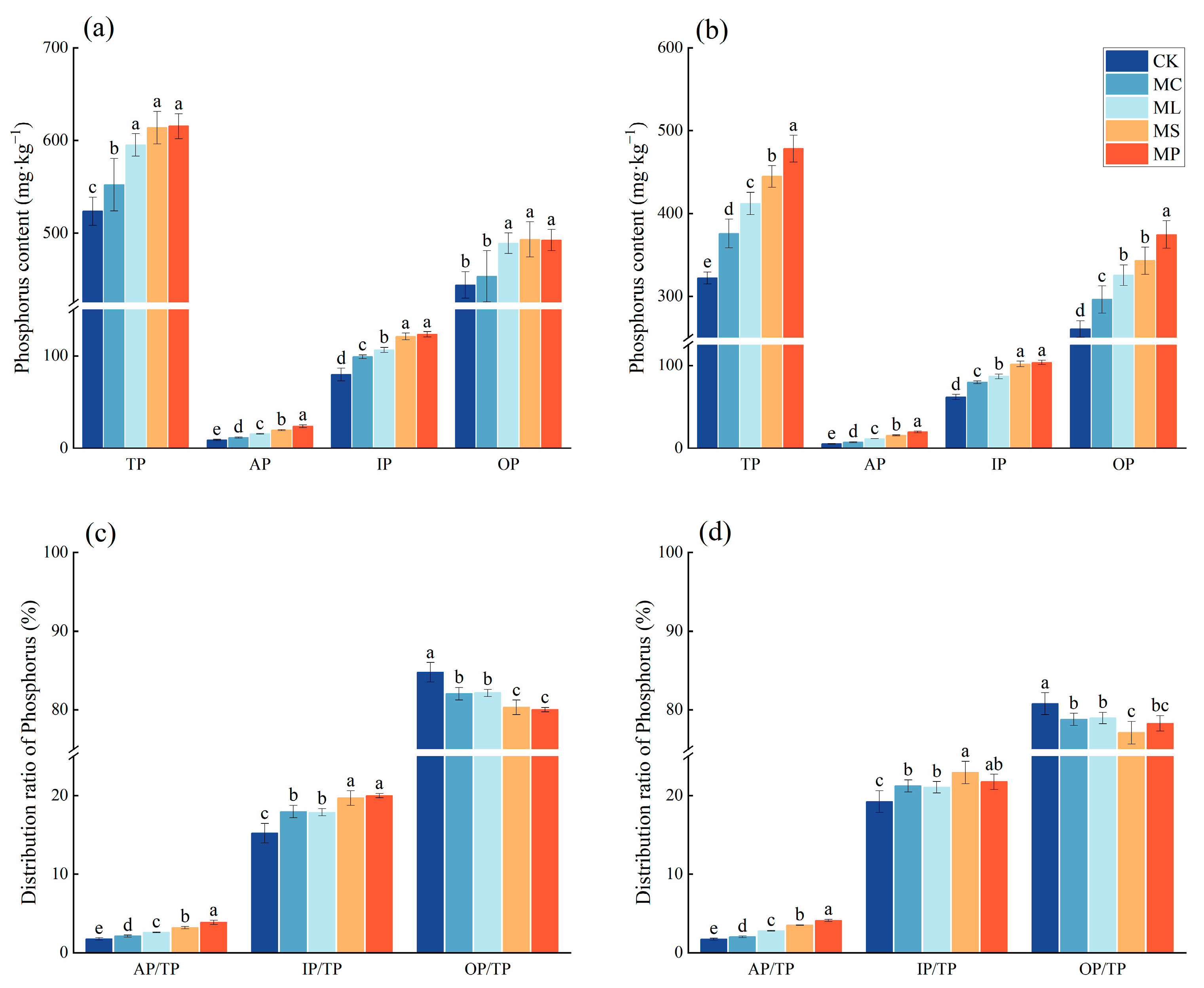
3.3. Soil P Fractions and Content
3.4. Soil Enzyme Activity and Microbial Abundance
3.5. Fine Root Characteristics
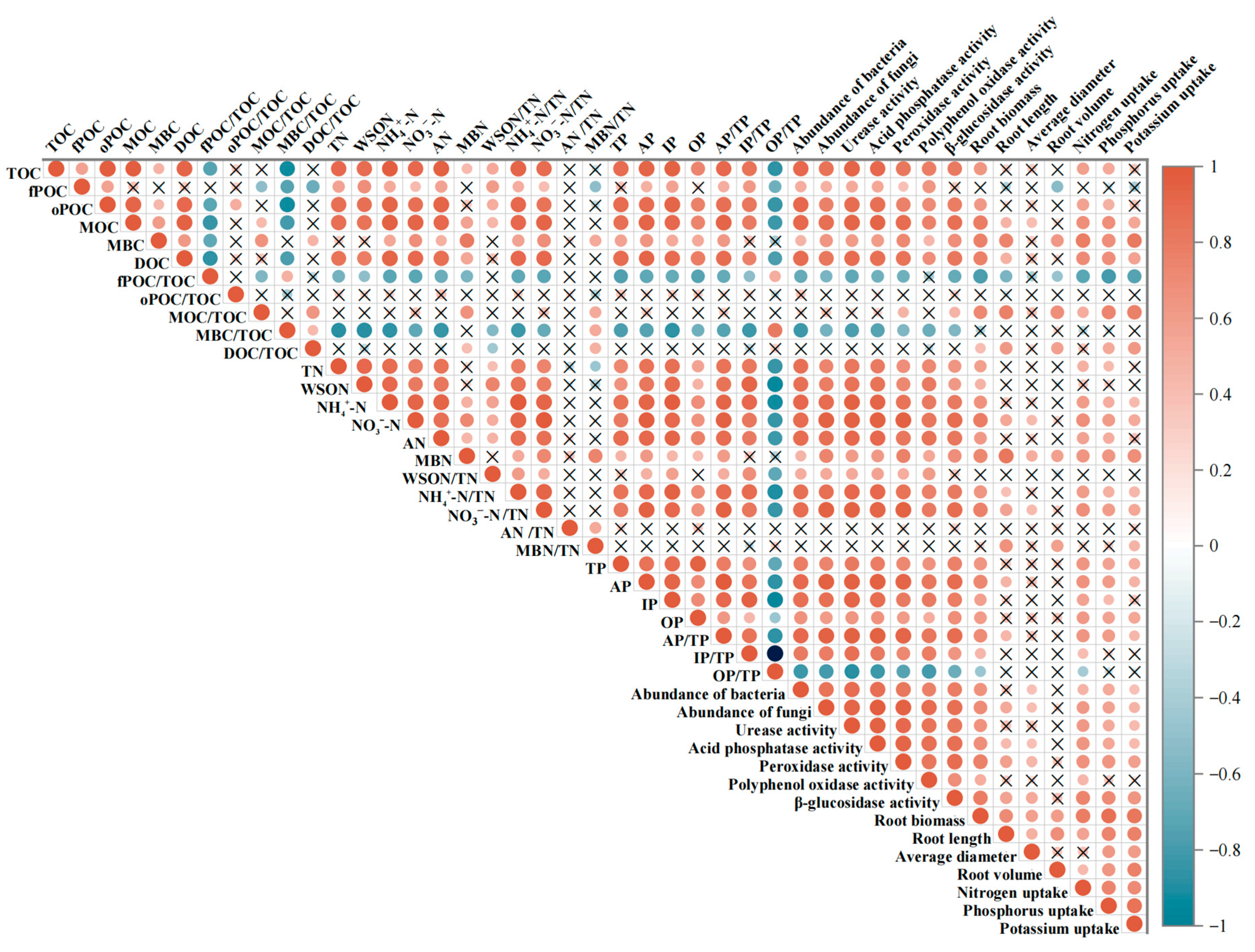
3.6. Relationship between Soil and Fine Root
4. Discussion
4.1. Effects of the Mixed Cultivation of Moso Bamboo with Other Indigenous Tree Species on Soil C, N, and P
4.2. Effects of Cultivating Moso Bamboo with Other Indigenous Tree Species on Soil Microbial Biomass, Abundance, and Enzyme Activity
4.3. Root Growth and Nutrient Uptake
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Song, X.; Zhou, G.; Jiang, H.; Yu, S.; Fu, J.; Li, W.; Wang, W.; Ma, Z.; Peng, C. Carbon Sequestration by Chinese Bamboo Forests and Their Ecological Benefits: Assessment of Potential, Problems, and Future Challenges. Environ. Rev. 2011, 19, 418–428. [Google Scholar] [CrossRef]
- Song, X.; Peng, C.; Ciais, P.; Li, Q.; Xiang, W.; Xiao, W.; Zhou, G.; Deng, L. Nitrogen Addition Increased CO2 Uptake More than Non-CO 2 Greenhouse Gases Emissions in a Moso Bamboo Forest. Sci. Adv. 2020, 6, eaaw5790. [Google Scholar] [CrossRef]
- Li, P.; Zhou, G.; Du, H.; Lu, D.; Mo, L.; Xu, X.; Shi, Y.; Zhou, Y. Current and Potential Carbon Stocks in Moso Bamboo Forests in China. J. Environ. Manag. 2015, 156, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Guo, L.; Ramakrishnan, M.; Fei, Z.; Vinod, K.K.; Ding, Y.; Jiao, C.; Gao, Z.; Zha, R.; Wang, C.; et al. Rapid Growth of Moso Bamboo (Phyllostachys edulis): Cellular Roadmaps, Transcriptome Dynamics, and Environmental Factors. Plant Cell 2022, 34, 3577–3610. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, K.; Jepson, P.; Wu, L.; Ramanuja Rao, I.V.; Jiang, S.; Liese, W.; Lou, Y.; Fu, M. The Potential of Bamboo Is Constrained by Outmoded Policy Frames. Ambio 2011, 40, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Yen, T.-M.; Lee, J.-S. Comparing Aboveground Carbon Sequestration between Moso Bamboo (Phyllostachys heterocycla) and China Fir (Cunninghamia lanceolata) Forests Based on the Allometric Model. For. Ecol. Manag. 2011, 261, 995–1002. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, P.; Wang, H.; Zhou, G.; Wu, J.; Yang, F.; Qian, X. Seasonal Soil CO2 Efflux Dynamics after Land Use Change from a Natural Forest to Moso Bamboo Plantations in Subtropical China. For. Ecol. Manag. 2011, 262, 1131–1137. [Google Scholar] [CrossRef]
- Qin, H.; Chen, J.; Wu, Q.; Niu, L.; Li, Y.; Liang, C.; Shen, Y.; Xu, Q. Intensive Management Decreases Soil Aggregation and Changes the Abundance and Community Compositions of Arbuscular Mycorrhizal Fungi in Moso Bamboo (Phyllostachys pubescens) Forests. For. Ecol. Manag. 2017, 400, 246–255. [Google Scholar] [CrossRef]
- Ni, H.; Su, W.; Fan, S.; Chu, H. Effects of Intensive Management Practices on Rhizosphere Soil Properties, Root Growth, and Nutrient Uptake in Moso Bamboo Plantations in Subtropical China. For. Ecol. Manag. 2021, 493, 119083. [Google Scholar] [CrossRef]
- Li, C.; Shi, Y.; Zhou, G.; Zhou, Y.; Xu, L.; Tong, L.; Liu, X. Effects of Different Management Approaches on Soil Carbon Dynamics in Moso Bamboo Forest Ecosystems. Catena 2018, 169, 59–68. [Google Scholar] [CrossRef]
- Xu, Q.; Jiang, P.; Xu, Z. Soil Microbial Functional Diversity under Intensively Managed Bamboo Plantations in Southern China. J. Soils Sediments 2008, 8, 177–183. [Google Scholar] [CrossRef]
- Li, Q.; Song, X.; Gu, H.; Gao, F. Nitrogen Deposition and Management Practices Increase Soil Microbial Biomass Carbon but Decrease Diversity in Moso Bamboo Plantations. Sci. Rep. 2016, 6, 28235. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Wen, X.; Wu, Z.; Zhong, H.; Zhang, X. Deciphering the Ramet System of a Bamboo Plant in Response to Intensive Management. Forests 2022, 13, 1968. [Google Scholar] [CrossRef]
- Gärtner, S.; Reif, A. The Impact of Forest Transformation on Stand Structure and Ground Vegetation in the Southern Black Forest, Germany. Plant Soil 2004, 264, 35–51. [Google Scholar] [CrossRef]
- Kaitaniemi, P.; Lintunen, A. Neighbor Identity and Competition Influence Tree Growth in Scots Pine, Siberian Larch, and Silver Birch. Ann. For. Sci. 2010, 67, 604. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Huang, S.; Fang, D. Impacts of Moso Bamboo (Phyllostachys pubescens) Invasion on Species Diversity and Aboveground Biomass of Secondary Coniferous and Broad-Leaved Mixed Forest. Front. Plant Sci. 2022, 13, 1001785. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xu, X.; Wu, Z.; Huang, Z.; Gao, G.; Zhang, J.; Zhang, X. Native Bamboo (Indosasa shibataeoides McClure) Invasion of Broadleaved Forests Promotes Soil Organic Carbon Sequestration in South China Karst. Forests 2023, 14, 2135. [Google Scholar] [CrossRef]
- Qin, H.; Niu, L.; Wu, Q.; Chen, J.; Li, Y.; Liang, C.; Xu, Q.; Fuhrmann, J.J.; Shen, Y. Bamboo Forest Expansion Increases Soil Organic Carbon through Its Effect on Soil Arbuscular Mycorrhizal Fungal Community and Abundance. Plant Soil 2017, 420, 407–421. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Fan, S.-H.; Guan, F.-Y.; Yan, X.-R.; Yin, Z.-X. Soil Bacterial Community Structure of Mixed Bamboo and Broad-Leaved Forest Based on Tree Crown Width Ratio. Sci. Rep. 2020, 10, 6522. [Google Scholar] [CrossRef]
- Bai, Y.; Chen, S.; Shi, S.; Qi, M.; Liu, X.; Wang, H.; Wang, Y.; Jiang, C. Effects of Different Management Approaches on the Stoichiometric Characteristics of Soil C, N, and P in a Mature Chinese Fir Plantation. Sci. Total Environ. 2020, 723, 137868. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Wang, M.; Liu, G.; Xue, S.; Yang, X. Disentangling the Direct and Indirect Effects of Cropland Abandonment on Soil Microbial Activity in Grassland Soil at Different Depths. Catena 2020, 194, 104774. [Google Scholar] [CrossRef]
- Cai, X.; Lin, Z.; Penttinen, P.; Li, Y.; Li, Y.; Luo, Y.; Yue, T.; Jiang, P.; Fu, W. Effects of Conversion from a Natural Evergreen Broadleaf Forest to a Moso Bamboo Plantation on the Soil Nutrient Pools, Microbial Biomass and Enzyme Activities in a Subtropical Area. For. Ecol. Manag. 2018, 422, 161–171. [Google Scholar] [CrossRef]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of Changes in Land Use/Cover on Soil Microbial and Enzyme Activities. Catena 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Wang, L.; Pang, X.; Li, N.; Qi, K.; Huang, J.; Yin, C. Effects of Vegetation Type, Fine and Coarse Roots on Soil Microbial Communities and Enzyme Activities in Eastern Tibetan Plateau. Catena 2020, 194, 104694. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Singh, K.P.; Singh, P.K. Temporal Changes in Spatial Pattern of Fine-root Mass and Nutrient Concentrations in Indian Bamboo Savanna. Appl. Veg. Sci. 1999, 2, 229–238. [Google Scholar] [CrossRef]
- Li, J.; Shangguan, Z.; Deng, L. Free Particulate Organic Carbon Plays Critical Roles in Carbon Accumulations during Grassland Succession since Grazing Exclusion. Soil Tillage Res. 2022, 220, 105380. [Google Scholar] [CrossRef]
- Tietz, A.; Kirschner, A.; Langergraber, G.; Sleytr, K.; Haberl, R. Characterisation of Microbial Biocoenosis in Vertical Subsurface Flow Constructed Wetlands. Sci. Total Environ. 2007, 380, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Chang, S.X.; Jiang, P.; Zhou, G.; Shen, Z.; Wu, J.; Lin, L.; Wang, Z.; Shen, M. Converting Native Shrub Forests to Chinese Chestnut Plantations and Subsequent Intensive Management Affected Soil C and N Pools. For. Ecol. Manag. 2014, 312, 161–169. [Google Scholar] [CrossRef]
- Bremner, J.B.; Smith, R.J.; Tarrant, G.J. A Meisenheimer Rearrangement Approach to Bridgehead Hydroxylated Tropane Alkaloid Derivatives. Tetrahedron Lett. 1996, 37, 97–100. [Google Scholar] [CrossRef]
- Li, F.-M.; Song, Q.-H.; Jjemba, P.K.; Shi, Y.-C. Dynamics of Soil Microbial Biomass C and Soil Fertility in Cropland Mulched with Plastic Film in a Semiarid Agro-Ecosystem. Soil Biol. Biochem. 2004, 36, 1893–1902. [Google Scholar] [CrossRef]
- Malik, M.A.; Marschner, P.; Khan, K.S. Addition of Organic and Inorganic P Sources to Soil—Effects on P Pools and Microorganisms. Soil Biol. Biochem. 2012, 49, 106–113. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The Effects of Long Term Nitrogen Deposition on Extracellular Enzyme Activity in an Acer Saccharum Forest Soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Guo, L.B.; Halliday, M.J.; Siakimotu, S.J.M.; Gifford, R.M. Fine Root Production and Litter Input: Its Effects on Soil Carbon. Plant Soil 2005, 272, 1–10. [Google Scholar] [CrossRef]
- Yang, C.; Ni, H.; Zhong, Z.; Zhang, X.; Bian, F. Changes in Soil Carbon Pools and Components Induced by Replacing Secondary Evergreen Broadleaf Forest with Moso Bamboo Plantations in Subtropical China. Catena 2019, 180, 309–319. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, L.; Guan, D.; Chen, Y.; Motelica-Heino, M.; Peng, Y.; Lee, S.Y. The Role of Mangrove Fine Root Production and Decomposition on Soil Organic Carbon Component Ratios. Ecol. Indic. 2021, 125, 107525. [Google Scholar] [CrossRef]
- Sheng, H.; Zhou, P.; Zhang, Y.; Kuzyakov, Y.; Zhou, Q.; Ge, T.; Wang, C. Loss of Labile Organic Carbon from Subsoil Due to Land-Use Changes in Subtropical China. Soil Biol. Biochem. 2015, 88, 148–157. [Google Scholar] [CrossRef]
- Han, X.; Zhao, F.; Tong, X.; Deng, J.; Yang, G.; Chen, L.; Kang, D. Understanding Soil Carbon Sequestration Following the Afforestation of Former Arable Land by Physical Fractionation. Catena 2017, 150, 317–327. [Google Scholar] [CrossRef]
- Budge, K.; Leifeld, J.; Hiltbrunner, E.; Fuhrer, J. Litter Quality and PH Are Strong Drivers of Carbon Turnover and Distribu-Tion in Alpine Grassland Soils. Biogeosciences 2010, 7. [Google Scholar] [CrossRef]
- Tong, X.; Xu, M.; Wang, X.; Bhattacharyya, R.; Zhang, W.; Cong, R. Long-Term Fertilization Effects on Organic Carbon Fractions in a Red Soil of China. Catena 2014, 113, 251–259. [Google Scholar] [CrossRef]
- McGILL, W.B.; Cannon, K.R.; Robertson, J.A.; Cook, F.D. Dynamics of Soil Microbial Biomass and Water-Soluble Organic c in Breton l after 50 Years of Cropping to Two Rotations. Can. J. Soil Sci. 1986, 66, 1–19. [Google Scholar] [CrossRef]
- Naorem, A.; Maverick, J.; Singh, P.; Udayana, S.K. Microbial Community Structure in Organic Farming and Their Management. In Advances in Organic Farming; Elsevier: Amsterdam, The Netherlands, 2021; pp. 47–58. ISBN 9780128223581. [Google Scholar]
- Yang, C.; Wang, A.; Zhu, Z.; Lin, S.; Bi, Y.; Du, X. Impact of Extensive Management System on Soil Properties and Carbon Sequestration under an Age Chronosequence of Moso Bamboo Plantations in Subtropical China. For. Ecol. Manag. 2021, 497, 119535. [Google Scholar] [CrossRef]
- Hansen, E.M.; Munkholm, L.J.; Melander, B.; Olesen, J.E. Can Non-Inversion Tillage and Straw Retainment Reduce N Leaching in Cereal-Based Crop Rotations? Soil Tillage Res. 2010, 109, 1–8. [Google Scholar] [CrossRef]
- Mancinelli, R.; Campiglia, E.; Di Tizio, A.; Marinari, S. Soil Carbon Dioxide Emission and Carbon Content as Affected by Conventional and Organic Cropping Systems in Mediterranean Environment. Appl. Soil Ecol. 2010, 46, 64–72. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global Analysis of Nitrogen and Phosphorus Limitation of Primary Producers in Freshwater, Marine and Terrestrial Ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Conant, R.T.; Zhou, G.; Wang, Y.; Wang, N.; Li, Y.; Zhang, K. Effects of Moso Bamboo Encroachment into Native, Broad-Leaved Forests on Soil Carbon and Nitrogen Pools. Sci. Rep. 2016, 6, 31480. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Mo, Q.; Wang, H.; Zhang, Z.; Huang, G.; Ye, Q.; Zou, Q.; Kong, F.; Liu, Y.; Geoff Wang, G. Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) Invasion Affects Soil Phosphorus Dynamics in Adjacent Coniferous Forests in Subtropical China. Ann. For. Sci. 2018, 75, 24. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Chang, S.X.; Xu, Q.; Li, Y.; Ma, Z.; Qin, H.; Cai, Y. Linking Enhanced Soil Nitrogen Mineralization to Increased Fungal Decomposition Capacity with Moso Bamboo Invasion of Broadleaf Forests. Sci. Total Environ. 2021, 771, 144779. [Google Scholar] [CrossRef] [PubMed]
- Chaer, G.; Fernandes, M.; Myrold, D.; Bottomley, P. Comparative Resistance and Resilience of Soil Microbial Communities and Enzyme Activities in Adjacent Native Forest and Agricultural Soils. Microb. Ecol. 2009, 58, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Li, Q.; Gai, X.; Zhang, X.; Zhong, Z.; Bian, F.; Yang, C. Effects of On- and off-Year Management Practices on the Soil Organic C Fractions and Microbial Community in a Moso Bamboo (Phyllostachys edulis) Forest in Subtropical China. Front. Plant Sci. 2022, 13, 1020344. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil Enzyme Activities, Microbial Communities, and Carbon and Nitrogen Availability in Organic Agroecosystems across an Intensively-Managed Agricultural Landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Luo, M.; Moorhead, D.L.; Ochoa-Hueso, R.; Mueller, C.W.; Ying, S.C.; Chen, J. Nitrogen Loading Enhances Phosphorus Limitation in Terrestrial Ecosystems with Implications for Soil Carbon Cycling. Funct. Ecol. 2022, 36, 2845–2858. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of Extracellular Enzymes to Simple and Complex Nutrient Inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Xing, S.; Chen, C.; Zhou, B.; Zhang, H.; Nang, Z.; Xu, Z. Soil Soluble Organic Nitrogen and Active Microbial Characteristics under Adjacent Coniferous and Broadleaf Plantation Forests. J. Soils Sediments 2010, 10, 748–757. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.-F.; Stokes, A. Root Structure–Function Relationships in 74 Species: Evidence of a Root Economics Spectrum Related to Carbon Economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Ostonen, I.; Helmisaari, H.-S.; Borken, W.; Tedersoo, L.; Kukumägi, M.; Bahram, M.; Lindroos, A.-J.; Nöjd, P.; Uri, V.; Merilä, P.; et al. Fine Root Foraging Strategies in Norway Spruce Forests across a European Climate Gradient. Glob. Change Biol. 2011, 17, 3620–3632. [Google Scholar] [CrossRef]
- Peng, C.; Tu, J.; Yang, M.; Meng, Y.; Li, M.; Ai, W. Root Stoichiometric Dynamics and Homeostasis of Invasive Species Phyllostachys Edulis and Native Species Cunninghamia Lanceolata in a Subtropical Forest in China. J. For. Res. 2021, 32, 2001–2010. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S., III; Mooney, H.A. Resource Limitation in Plants-an Economic Analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Sokol, N.W.; Bradford, M.A. Microbial Formation of Stable Soil Carbon Is More Efficient from Belowground than Aboveground Input. Nat. Geosci. 2019, 12, 46–53. [Google Scholar] [CrossRef]
- Chen, G.-T.; Tu, L.-H.; Peng, Y.; Hu, H.-L.; Hu, T.-X.; Xu, Z.-F.; Liu, L.; Tang, Y. Effect of Nitrogen Additions on Root Morphology and Chemistry in a Subtropical Bamboo Forest. Plant Soil 2017, 412, 441–451. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Uses | WSON/TN | NH4+-N/TN | NO3−-N/TN | AN/TN | MBN/TN |
|---|---|---|---|---|---|
| 0–20 cm | |||||
| CK | 1.815 ± 0.049 b | 0.058 ± 0.002 d | 0.017 ± 0.001 d | 10.863 ± 0.446 a | 7.941 ± 0.795 a |
| MC | 1.93 ± 0.102 a | 0.069 ± 0.004 c | 0.02 ± 0.001 d | 10.732 ± 0.812 a | 6.037 ± 0.387 d |
| ML | 1.883 ± 0.087 ab | 0.076 ± 0.006 b | 0.033 ± 0.007 c | 10.677 ± 0.194 a | 6.372 ± 0.503 cd |
| MS | 1.936 ± 0.035 a | 0.089 ± 0.003 a | 0.05 ± 0.003 b | 10.806 ± 0.154 a | 7.096 ± 0.194 b |
| MP | 1.965 ± 0.034 a | 0.088 ± 0.002 a | 0.059 ± 0.005 a | 11.035 ± 0.302 a | 7.325 ± 0.281 ab |
| 20–40 cm | |||||
| CK | 3.974 ± 0.338 a | 0.058 ± 0.007 c | 0.024 ± 0.003 c | 20.237 ± 1.987 a | 14.973 ± 1.821 a |
| MC | 3.215 ± 0.341 bc | 0.047 ± 0.004 d | 0.026 ± 0.002 bc | 16.513 ± 0.823 b | 8.407 ± 0.664 c |
| ML | 3.331 ± 0.145 b | 0.047 ± 0.003 d | 0.027 ± 0.002 ab | 16.387 ± 0.668 b | 8.98 ± 0.683 c |
| MS | 3.299 ± 0.142 b | 0.076 ± 0.005 a | 0.03 ± 0.001 a | 16.424 ± 0.265 b | 10.661 ± 0.451 b |
| MP | 2.937 ± 0.171 c | 0.069 ± 0.004 b | 0.028 ± 0.001 ab | 14.569 ± 0.721 c | 9.605 ± 0.574 bc |
| Land Uses | Abundance of Bacteria | Abundance of Fungi | Urease Activity | Acid Phosphatase Activity | Peroxidase Activity | Polyphenol Oxidase Activity | β-glucosidase Activity |
|---|---|---|---|---|---|---|---|
| ×108 Copies g−1 Soil | ×105 Copies g−1 Soil | μmol g−1 h−1 | μmol g−1 h−1 | μmol g−1 h−1 | μmol g−1 h−1 | μmol g−1 h−1 | |
| 0–20 cm | |||||||
| CK | 2.5 ± 0.087 d | 2.642 ± 0.145 e | 532.868 ± 19.202 e | 19.596 ± 1.269 e | 52.83 ± 1.007 d | 5.232 ± 0.208 d | 51.08 ± 3.270 d |
| MC | 2.682 ± 0.073 c | 2.998 ± 0.075 d | 581.288 ± 11.427 d | 22.462 ± 0.657 d | 51.754 ± 2.337 d | 6.138 ± 0.155 c | 49.928 ± 4.273 d |
| ML | 2.846 ± 0.047 b | 3.306 ± 0.173 c | 617.306 ± 13.985 c | 24.91 ± 0.864 c | 57.268 ± 1.625 c | 5.506 ± 0.321 d | 56.91 ± 2.662 c |
| MS | 2.918 ± 0.106 b | 5.176 ± 0.202 b | 677.216 ± 17.761 b | 29.562 ± 1.263 b | 63.682 ± 1.606 b | 6.944 ± 0.275 b | 62.092 ± 2.547 b |
| MP | 3.04 ± 0.060 a | 6.23 ± 0.269 a | 723.556 ± 17.620 a | 33.38 ± 1.649 a | 70.652 ± 2.134 a | 7.418 ± 0.260 a | 67.138 ± 1.972 a |
| 20–40 cm | |||||||
| CK | 1.458 ± 0.098 d | 1.528 ± 0.147 e | 454.878 ± 17.426 e | 13.396 ± 1.575 e | 37.298 ± 1.047 d | 3.328 ± 0.300 d | 33.238 ± 1.665 d |
| MC | 1.686 ± 0.082 c | 1.874 ± 0.110 d | 517.298 ± 13.558 d | 16.296 ± 0.472 d | 36.456 ± 1.467 d | 4.208 ± 0.130 c | 32.648 ± 1.994 d |
| ML | 1.842 ± 0.051 b | 2.158 ± 0.123 c | 561.056 ± 15.391 c | 18.83 ± 0.812 c | 41.638 ± 1.689 c | 3.638 ± 0.266 d | 39.706 ± 2.589 c |
| MS | 1.89 ± 0.116 b | 4.012 ± 0.275 b | 617.834 ± 9.140 b | 23.242 ± 1.079 b | 47.872 ± 1.584 b | 5.094 ± 0.209 b | 45.932 ± 2.212 b |
| MP | 2.04 ± 0.048 a | 5.088 ± 0.303 a | 659.382 ± 10.742 a | 26.24 ± 0.527 a | 55.008 ± 1.468 a | 5.508 ± 0.327 a | 50.458 ± 1.163 a |
| Land Uses | Root Biomass (g m−3) | Root Length (cm) | Average Diameter (mm) | Root Volume (cm3) | Nitrogen Uptake (kg ha−1) | Phosphorus Uptake (kg ha−1) | Potassium Uptake (kg ha−1) |
|---|---|---|---|---|---|---|---|
| CK | 11,897.472 ± 1212.803 b | 1339.876 ± 26.894 a | 0.54992 ± 0.044 ab | 3.5672 ± 0.204 a | 59.232 ± 1.777 bc | 6.334 ± 0.649 b | 57.718 ± 1.944 a |
| MC | 10,013.932 ± 832.825 c | 1033.38 ± 58.327 c | 0.5121 ± 0.019 b | 2.648 ± 0.430 b | 55.704 ± 3.547 c | 4.996 ± 0.388 c | 49.098 ± 2.684 b |
| ML | 12,668.54 ± 649.367 b | 1236.21 ± 109.806 b | 0.55006 ± 0.020ab | 3.3688 ± 0.190a | 60.934 ± 1.479 ab | 6.748 ± 0.430 ab | 58.32 ± 1.470 a |
| MS | 14,412.908 ± 523.477 a | 1404.23 ± 71.691 a | 0.56136 ± 0.038 a | 3.383 ± 0.195 a | 63.624 ± 2.834 a | 6.99 ± 0.333 a | 60.318 ± 1.268 a |
| MP | 14,090.13 ± 843.828 a | 1374.714 ± 93.141 a | 0.57158 ± 0.036 a | 3.4554 ± 0.292 a | 64.032 ± 2.469 a | 7.064 ± 0.142 a | 59.03 ± 2.220 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, Y.; Chen, Z.; Gao, H.; Yang, C.; Zhang, X.; Wang, Z.; Wang, A.; Du, X.; Lan, L.; Bi, Y. Introducing Native Tree Species Alter the Soil Organic Carbon, Nitrogen, Phosphorus, and Fine Roots in Moso Bamboo Plantations. Forests 2024, 15, 971. https://doi.org/10.3390/f15060971
Ning Y, Chen Z, Gao H, Yang C, Zhang X, Wang Z, Wang A, Du X, Lan L, Bi Y. Introducing Native Tree Species Alter the Soil Organic Carbon, Nitrogen, Phosphorus, and Fine Roots in Moso Bamboo Plantations. Forests. 2024; 15(6):971. https://doi.org/10.3390/f15060971
Chicago/Turabian StyleNing, Yilin, Zedong Chen, Hongdi Gao, Chuanbao Yang, Xu Zhang, Zijie Wang, Anke Wang, Xuhua Du, Lan Lan, and Yufang Bi. 2024. "Introducing Native Tree Species Alter the Soil Organic Carbon, Nitrogen, Phosphorus, and Fine Roots in Moso Bamboo Plantations" Forests 15, no. 6: 971. https://doi.org/10.3390/f15060971