


Does Soil Acidification Matter? Nutrient Sustainability of Timber Harvesting in Forests on Selected Soils Developed in Sediments of the Early vs. Late Pleistocene

 , ,
, , 
Abstract
:
1. Introduction
- (1)
- Are the soils from sediments of the early Pleistocene more weathered, acidified, and thus poorer or richer in nutrients than soils from sediments of the late Pleistocene? To what extent do the nutrient fluxes differ between the two study areas? In particular, are there differences in weathering and leachate losses between heavily weathered and younger soils? Are there differences in nutrient exports due to timber harvesting because the tree biomass on early Pleistocene till is poorer or richer in nutrients than that on late Pleistocene till?
- (2)
- Is the nutrient sustainability of sites with early Pleistocene substrate more at risk than that of sites with late Pleistocene substrate given comparable timber harvesting?
- (3)
- If this is not the case, what other tools can help practitioners decide on the level of intervention of harvesting-related measures?
- Due to longer soil development, the soils of the early Pleistocene are more weathered and have different nutrient contents than those of the late Pleistocene. As a result, they have a different current weathering rate, different leachate losses, and different nutrient exports through timber harvesting.
- As a consequence, nutrient sustainability is more at risk on early Pleistocene sites than on late Pleistocene sites with comparable timber harvesting intensity.
- If there are no significant differences between early and late Pleistocene sites in terms of nutrient sustainability, acidification parameters such as pH, base saturation, cation exchange capacity, or nutrient stocks in the soil can be used to predict nutrient sustainability in multiple linear models.
2. Materials and Methods
2.1. Study Sites
2.2. Experimental Design
2.3. Sampling and Analyses of Tree Components
- Mixed sample of the bark of the merchantable wood (above diameter 7 cm);
- Brushwood < 5 mm (buds included);
- Thin branches (5–20 mm);
- Thick branches (20–70 mm);
- Stem disk of small merchantable wood (70–120 mm);
- Stem disk of medium merchantable wood (120–250 mm);
- Stem disk of thick merchantable wood (>250 mm).
2.4. Soil Analyses
2.5. Calculation of Nutrient Fluxes and Nutrient Stock in Soil
2.5.1. Deposition
2.5.2. Weathering and Leaching Loss
2.5.3. Nutrient and Organic Carbon Stocks in the Root Zone and the Amount of Clay
2.5.4. CEC for the Root Zone, Mean pH, and BS
2.6. Simulation of Forest Growth and Timber Harvesting
2.7. Assessment of Nutrient Sustainability
2.8. Statistical Analyses
3. Results
3.1. Site Conditions
3.2. Biomass and Nutrient Export by Harvesting
3.3. Dependence of Nutrient Balance on Acidification
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Site Name | Longitude | Lattitude | Altitude [m] | Exposure | Slope [%] | Tree Species | Soil Type | Geological Unit |
|---|---|---|---|---|---|---|---|---|---|
| Bülach | Lärchenischlag | 8.52747 | 47.52564 | 428 | NW | 7 | Bu, BAh, Li | Calcic Luvisol | Late Pleistocene |
| Chengelboden | 8.52879 | 47.53423 | 435 | SE | 3 | Bu, Ta, Fi, BAh | Dystric Cambisol | ||
| Lindi | 8.53340 | 47.54641 | 423 | - | 0 | Bu, Fö, Fi, BAh | Calcic Luvisol | ||
| Marterloch | 8.53648 | 47.53980 | 432 | - | 0 | Bu, Fi, Fö | Calcic Luvisol | ||
| Brengspel | 8.53650 | 47.53054 | 451 | - | 1 | Bu, BAh, Fö, Lä, Ei | Calcaric Cambisol | ||
| Irchel | Hörnli | 8.58262 | 47.55627 | 676 | - | 0 | Bu, Ei, Fi | Haplic Luvisol | Early Pleistocene |
| Schartenflue | 8.59637 | 47.54898 | 668 | - | 2 | Bu, Ei | Calcic Luvisol | ||
| Schaffhuser | 8.60396 | 47.54749 | 669 | NE | 50 | Bu, Fi, Es | Eutric Cambisol | ||
| Obermeser | 8.61025 | 47.53738 | 692 | NW | 2 | Bu, Fi, Dougl | Haplic Luvisol | ||
| Steig | 8.61735 | 47.53603 | 680 | W | 3 | Bu, Fi, Dougl | Haplic Luvisol |
| Site | Pedogenetic Horizon and Its Depth [cm] | Stone Content [%] | Texture | Density of the Fine Earth [Mg·m−3] | Hydromorphic Features | Soil Color |
|---|---|---|---|---|---|---|
| Lärchenischlag | L (1.5–0) | - | - | 0.10 | no | - |
| Ah (0–10) | 11–25 | Sl | 0.67 | no | 10YR 3/3 | |
| (E)AB (10–30) | 26–50 | Sl | 0.72 | no | 10YR 4/4 | |
| B(t) (30–50) | 51–75 | Lts | 0.74 | no | 10YR 4/6 | |
| CBv (50–80) | 51–75 | Sl | 0.78 | no | 10YR 5/6 | |
| Cca1 (80–110) | >75 | Ss | 0.78 | no | 10YR 5/2 | |
| Cca2 (110–135) | >75 | Ss | 0.81 | no | 10YR 5/2 | |
| Chengelboden | L (3–0.5) | - | - | 0.10 | no | - |
| [F] (0.5–0) | - | - | 0.15 | no | - | |
| Ah (0–6) | <2 | Slu | 0.91 | no | 10YR 2/3 | |
| (A)B (6–20) | <2 | Lu | 0.91 | no | 10YR 4/4 | |
| Bv (20–40) | 11–25 | Lu | 1.19 | no | 10YR 5/4 | |
| BSw (40–60) | <2 | Lu | 1.28 | Mn concretions | 10YR 6/3 | |
| Sw (60–100) | <2 | Lu | 1.49 | Mn concretions | 2.5Y 6/4 | |
| IISw (100–140) | <2 | Ls | 1.55 | pale-red colors | 2.5Y 5/4 | |
| IISd (140–195) | <2 | Ls | 1.59 | pale-red colors | 2.5Y 5/3 | |
| IICca (195–220) | >75 | Sl | 1.46 | no | 10YR 5/2 | |
| Lindi | L (3–1) | - | - | 0.10 | no | - |
| F (1–0) | - | - | 0.15 | no | - | |
| Ah (0–4) | <2 | Sl | 0.88 | no | 10YR 3/3 | |
| (EA)B (4–20) | <2 | Ls | 0.88 | no | 10YR 4/4 | |
| (E)B (20–40) | 2–10 | Ls | 1.17 | no | 10YR 5/6 | |
| B(t) (40–55) | 2–10 | Ls | 0.87 | no | 10YR 5/6 | |
| CB(t) (55–80) | 26–50 | Lts | 0.87 | no | 10YR 4/6 | |
| B(t) (80–100) | <2 | Sl | 1.33 | no | 10YR 5/6 | |
| Cca (100–155) | >75 | Sl | 1.20 | no | 10YR 6/3 | |
| Marterloch | L (1–0) | - | - | 0.10 | no | - |
| Ah (0–5) | 2–10 | Sl | 0.91 | no | 10YR 2/3 | |
| (EA)B (5–25) | 11–25 | Ls | 0.91 | no | 10YR 4/3 | |
| (E)B(cn) (25–60) | 11–25 | Lu | 1.09 | Mn concretions | 10YR 5/6 | |
| B(t,cn) (60–80) | 26–50 | Lts | 1.10 | Mn concretions | 10YR 4/4 | |
| CB(t) (80–105) | 51–75 | Lts | 0.95 | no | 10YR 4/4 | |
| Cca (105–140) | >75 | St | 0.95 | no | 10YR 6/1 | |
| Brengspel | L (1–0) | - | - | 0.10 | no | - |
| Ah (0–15) | 11–25 | Sl | 0.67 | no | 10YR 3/2 | |
| (A)B (15–30) | 26–50 | Sl | 0.72 | no | 10YR 4/4 | |
| AB (30–40) | >75 | Ls | 0.79 | no | 10YR 4/6 | |
| Cca1 (40–70) | >75 | Sl | 0.71 | no | 10YR 5/4 | |
| Cb,ca (70–90) | <2 | Sl | 1.15 | no | 10YR 6/2 |
| Site | Pedogenetic Horizon and Its Depth [cm] | Stone Content [%] | Texture | Density of the Fine Earth [Mg·m−3] | Hydromorphic Features | Soil Color |
|---|---|---|---|---|---|---|
| Hörnli | L (1–0) | - | - | 0.10 | no | - |
| Ah (0–6) | <2 | Sl | 0.99 | no | 10YR 3/2 | |
| (A)B (6–25) | <2 | Ls | 0.99 | no | 10YR 4/4 | |
| (Sw)B (25–65) | 2–10 | Ls | 1.41 | Mn concretions | 10YR 4/6 | |
| Sd1 (65–90) | 11–25 | Lt | 1.41 | Mn concretions | 10YR 6/8 | |
| Sd2 (90–160) | 26–50 | Lt | 1.44 | Mn concretions | 10YR 5/6 | |
| Sd3 (160–225) | 26–50 | Lt | 1.51 | Mn concretions | 10YR 5/6 | |
| Schartenflue | L (2–0) | - | - | 0.10 | no | - |
| Ah (0–6) | 26–50 | Sl | 0.59 | no | 10YR 3/2 | |
| (E)AB (6–15) | 26–50 | Sl | 0.59 | no | 10YR 4/3 | |
| (E)Bv (15–40) | 26–50 | Sl | 0.85 | no | 10YR 5/6 | |
| (E)CBv (40–60) | 51–75 | Sl | 1.08 | no | 10YR 4/6 | |
| (Bt)C (60–100) | >75 | Lts | 1.08 | no | 10YR 4/6 | |
| Cca (100–140) | >75 | St | 1.13 | no | 10YR 5/2 | |
| Schaffhuser | L (2–0) | - | - | 0.10 | no | - |
| Ah (0–10) | 11–25 | Lts | 0.67 | no | 10YR 3/2 | |
| (E)AB (10–20) | 11–25 | Ls | 0.72 | no | 10YR 4/2 | |
| B(t) (20–40) | 11–25 | Lts | 0.89 | no | 10YR 5/4 | |
| Bcn (40–55) | 11–25 | Lt | 1.06 | Mn concretions | 10YR 6/6 | |
| CB (55–80) | 51–75 | Lts | 0.91 | no | 10YR 5/6 | |
| (Bcn)Cca (80–120) | <2 | Lts | 1.36 | Mn concretions | 10YR 6/4 | |
| Rca (120–130) | >75 | - | 1.47 | no | - | |
| Obermeser | L (1.5–0) | - | - | 0.10 | no | - |
| Ah (0–6) | 11–25 | Sl | 0.72 | no | 10YR 4/2 | |
| (EA)B (6–16) | 11–25 | Ls | 0.72 | no | 10YR 4/4 | |
| (E)Bcn1 (16–35) | 11–25 | Ls | 1.04 | Mn concretions | 10YR 6/6 | |
| (E)Bcn2 (35–50) | 11–25 | Ls | 1.04 | Mn concretions | 10YR 5/4 | |
| B(t),cn (50–65) | 11–25 | Ls | 1.24 | Mn concretions | 10YR 5/6 | |
| Sw (65–95) | 11–25 | Lt | 1.21 | Mn concretions | 10YR 6/6 | |
| Sd (95–130) | 11–25 | Lt | 1.34 | Mn concretions | 10YR 5/6 | |
| SdC (130–220) | 51–75 | Ls | 1.30 | Mn concretions | 10YR 5/8 | |
| Steig | L (2–0.5) | - | - | 0.10 | no | - |
| F (0.5–0) | - | - | 0.15 | no | - | |
| A (0–15) | 11–25 | Ls | 0.98 | no | 10YR 3/2 | |
| ElB (15–40) | 11–25 | Lt | 1.15 | no | 10YR 6/6 | |
| Bt (40–140) | 11–25 | Lts | 1.35 | Mn concretions | 7.5YR 5/6 | |
| BC (140–200) | 11–25 | Lts | 1.43 | Mn concretions | 10YR 4/6 |
| Soil Property | Lärchenischlag | Chengelboden | Lindi | Marterloch | Brengspel | Hörnli | Schartenflue | Schaffhuser | Obermeser | Steig |
|---|---|---|---|---|---|---|---|---|---|---|
| Mean pH-value | 4.05 | 3.76 | 3.72 | 3.70 | 4.96 | 3.72 | 3.83 | 4.59 | 3.61 | 3.77 |
| Mean BS [%] | 62.6 | 53.3 | 29.9 | 29.8 | 96.3 | 34.5 | 38.0 | 96.2 | 31.6 | 33.0 |
| CEC [molc·m−2] | 23.2 | 143.7 | 53.2 | 52.9 | 69.6 | 74.0 | 15.5 | 242.1 | 162.4 | 305.0 |
| Amount of clay [kg·m−2] | 47.1 | 430.2 | 165.2 | 175.3 | 61.4 | 222.8 | 39.2 | 278.0 | 380.6 | 796.8 |
| Stock of Corg [kg·m−2] | 4.7 | 7.4 | 7.2 | 5.5 | 7.1 | 5.7 | 4.1 | 7.7 | 7.6 | 8.3 |
| Caexc [mol·m−2] | 6.8 | 23.2 | 4.7 | 5.4 | 32.9 | 6.3 | 2.4 | 111.9 | 15.5 | 24.9 |
| Mgexc [mol·m−2] | 0.30 | 12.92 | 2.60 | 1.89 | 0.42 | 5.67 | 0.47 | 2.67 | 8.62 | 22.89 |
| Kexc [mol·m−2] | 0.26 | 3.74 | 0.99 | 1.01 | 0.42 | 1.52 | 0.16 | 3.16 | 3.11 | 4.96 |
| Plak [mmol·m−2] | 98.3 | 3746.7 | 960.4 | 582.6 | 279.7 | 49.3 | 43.4 | 152.0 | 441.7 | 290.2 |
| Ntot [mol·m−2] | 22.1 | 73.4 | 39.9 | 33.5 | 33.4 | 29.9 | 16.7 | 44.4 | 47.2 | 76.8 |
| Correlation Matrix | CEC [molc·m−2] | Depth of Root Zone [m] | Amount of Clay [kg·m−2] | Stock of Corg [kg·m−2] | Caexc [mol·m−2] | Mgexc [mol·m−2] | Kexc [mol·m−2] |
|---|---|---|---|---|---|---|---|
| CEC [molc·m−2] | 1 | ||||||
| Depth of root zone [m] | 0.82 | 1 | |||||
| Amount of clay [kg·m−2] | 0.88 | 0.87 | 1 | ||||
| Stock of Corg [kg·m−2] | 0.81 | 0.77 | 0.71 | 1 | |||
| Caexc [mol·m−2] | 0.60 | 0.25 | 0.17 | 0.49 | 1 | ||
| Mgexc [mol·m−2] | 0.77 | 0.84 | 0.97 | 0.62 | 4.2 × 10−3 | 1 | |
| Kexc [mol·m−2] | 0.93 | 0.81 | 0.95 | 0.78 | 0.39 | 0.89 | 1 |
| Site | Study Area | Thinning Intensity | Ca [kg·ha−1·y−1] | Mg [kg·ha−1·y−1] | K [kg·ha−1·y−1] | P [kg·ha−1·y−1] |
|---|---|---|---|---|---|---|
| Lärchenischlag | Bülach | HIGH | 12.26 | 1.09 | 3.61 | 0.39 |
| Chengelboden | 13.56 | 19.44 | 69.99 | 4.07 | ||
| Lindi | 2.96 | 4.46 | 13.39 | 0.86 | ||
| Marterloch | 3.17 | 5.89 | 14.71 | 1.00 | ||
| Brengspel | 549.49 | 2.34 | 3.72 | 0.61 | ||
| Hörnli | Irchel | 1.68 | 5.25 | 14.75 | 0.00 | |
| Schartenflue | 0.54 | 0.48 | 1.40 | 0.14 | ||
| Schaffhuser | 624.39 | 8.59 | 10.03 | 0.36 | ||
| Obermeser | 1.90 | 11.14 | 39.81 | 0.00 | ||
| Steig | 3.09 | 19.58 | 64.28 | 0.00 | ||
| Lärchenischlag | Bülach | LOW | 12.25 | 1.08 | 3.56 | 0.39 |
| Chengelboden | 13.54 | 19.42 | 69.88 | 4.07 | ||
| Lindi | 2.96 | 4.46 | 13.42 | 0.86 | ||
| Marterloch | 3.17 | 5.88 | 14.65 | 1.00 | ||
| Brengspel | 549.88 | 2.35 | 3.74 | 0.61 | ||
| Hörnli | Irchel | 1.69 | 5.27 | 14.86 | 0.00 | |
| Schartenflue | 0.54 | 0.48 | 1.40 | 0.14 | ||
| Schaffhuser | 624.09 | 8.59 | 10.01 | 0.36 | ||
| Obermeser | 1.90 | 11.14 | 39.80 | 0.00 | ||
| Steig | 3.09 | 19.60 | 64.38 | 0.00 | ||
| Lärchenischlag | Bülach | BAU | 12.26 | 1.09 | 3.61 | 0.39 |
| Chengelboden | 13.55 | 19.44 | 69.97 | 4.07 | ||
| Lindi | 2.96 | 4.46 | 13.42 | 0.86 | ||
| Marterloch | 3.17 | 5.89 | 14.71 | 1.00 | ||
| Brengspel | 549.57 | 2.35 | 3.73 | 0.61 | ||
| Hörnli | Irchel | 1.68 | 5.26 | 14.80 | 0.00 | |
| Schartenflue | 0.54 | 0.48 | 1.40 | 0.14 | ||
| Schaffhuser | 624.29 | 8.59 | 10.02 | 0.36 | ||
| Obermeser | 1.90 | 11.13 | 39.79 | 0.00 | ||
| Steig | 3.09 | 19.58 | 64.29 | 0.00 |
| Site | Study Area | Ca [kg·ha−1·y−1] | Mg [kg·ha−1·y−1] | K [kg·ha−1·y−1] | P [kg·ha−1·y−1] | N [kg·ha−1·y−1] |
|---|---|---|---|---|---|---|
| Lärchenischlag | Bülach | 6.37 | 0.67 | 2.82 | 0.74 | 25.35 |
| Chengelboden | 6.31 | 0.67 | 2.78 | 0.74 | 22.41 | |
| Lindi | 6.14 | 0.65 | 2.65 | 0.74 | 24.51 | |
| Marterloch | 6.20 | 0.65 | 2.70 | 0.74 | 22.44 | |
| Brengspel | 6.30 | 0.67 | 2.77 | 0.74 | 24.73 | |
| Hörnli | Irchel | 6.18 | 0.65 | 2.64 | 0.73 | 18.61 |
| Schartenflue | 6.18 | 0.65 | 2.64 | 0.73 | 18.85 | |
| Schaffhuser | 6.30 | 0.67 | 2.73 | 0.73 | 20.67 | |
| Obermeser | 6.15 | 0.65 | 2.62 | 0.73 | 21.04 | |
| Steig | 6.21 | 0.66 | 2.67 | 0.73 | 20.88 | |
| Lärchenischlag | Bülach | 6.37 | 0.67 | 2.82 | 0.74 | 25.35 |
| Chengelboden | 6.31 | 0.67 | 2.78 | 0.74 | 22.41 | |
| Lindi | 6.14 | 0.65 | 2.65 | 0.74 | 24.51 | |
| Marterloch | 6.20 | 0.65 | 2.70 | 0.74 | 22.44 | |
| Brengspel | 6.30 | 0.67 | 2.77 | 0.74 | 24.73 | |
| Hörnli | Irchel | 6.18 | 0.65 | 2.64 | 0.73 | 18.61 |
| Schartenflue | 6.18 | 0.65 | 2.64 | 0.73 | 18.85 | |
| Schaffhuser | 6.30 | 0.67 | 2.73 | 0.73 | 20.67 | |
| Obermeser | 6.15 | 0.65 | 2.62 | 0.73 | 21.04 | |
| Steig | 6.21 | 0.66 | 2.67 | 0.73 | 20.88 | |
| Lärchenischlag | Bülach | 6.37 | 0.67 | 2.82 | 0.74 | 25.35 |
| Chengelboden | 6.31 | 0.67 | 2.78 | 0.74 | 22.41 | |
| Lindi | 6.14 | 0.65 | 2.65 | 0.74 | 24.51 | |
| Marterloch | 6.20 | 0.65 | 2.70 | 0.74 | 22.44 | |
| Brengspel | 6.30 | 0.67 | 2.77 | 0.74 | 24.73 | |
| Hörnli | Irchel | 6.18 | 0.65 | 2.64 | 0.73 | 18.61 |
| Schartenflue | 6.18 | 0.65 | 2.64 | 0.73 | 18.85 | |
| Schaffhuser | 6.30 | 0.67 | 2.73 | 0.73 | 20.67 | |
| Obermeser | 6.15 | 0.65 | 2.62 | 0.73 | 21.04 | |
| Steig | 6.21 | 0.66 | 2.67 | 0.73 | 20.88 |
| Site | Study Area | Thinning Intensity | Ca [kg·ha−1·y−1] | Mg [kg·ha−1·y−1] | K [kg·ha−1·y−1] | P [kg·ha−1·y−1] | N [kg·ha−1·y−1] |
|---|---|---|---|---|---|---|---|
| Lärchenischlag | Bülach | HIGH | 1.50 | 0.00 | 0.00 | 0.08 | 0.00 |
| Chengelboden | 0.00 | 15.20 | 55.28 | 0.08 | 0.00 | ||
| Lindi | 0.00 | 1.87 | 4.33 | 0.08 | 0.00 | ||
| Marterloch | 0.00 | 3.01 | 5.90 | 0.08 | 0.00 | ||
| Brengspel | 446.02 | 0.07 | 0.00 | 0.08 | 0.00 | ||
| Hörnli | Irchel | 0.00 | 3.11 | 7.25 | 0.08 | 0.00 | |
| Schartenflue | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 | ||
| Schaffhuser | 606.01 | 5.78 | 2.49 | 0.08 | 0.00 | ||
| Obermeser | 0.00 | 8.57 | 29.57 | 0.08 | 0.00 | ||
| Steig | 0.00 | 17.17 | 55.02 | 0.08 | 0.00 | ||
| Lärchenischlag | Bülach | LOW | 2.39 | 0.00 | 0.00 | 0.08 | 0.04 |
| Chengelboden | 0.00 | 15.63 | 56.69 | 0.08 | 0.00 | ||
| Lindi | 0.00 | 1.66 | 3.50 | 0.08 | 0.00 | ||
| Marterloch | 0.00 | 3.18 | 6.42 | 0.08 | 0.00 | ||
| Brengspel | 443.87 | 0.06 | 0.00 | 0.08 | 0.00 | ||
| Hörnli | Irchel | 0.00 | 2.57 | 5.33 | 0.08 | 0.00 | |
| Schartenflue | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 | ||
| Schaffhuser | 607.29 | 6.00 | 3.22 | 0.08 | 0.00 | ||
| Obermeser | 0.00 | 8.63 | 29.70 | 0.08 | 0.00 | ||
| Steig | 0.00 | 16.78 | 53.69 | 0.08 | 0.00 | ||
| Lärchenischlag | Bülach | BAU | 1.38 | 0.00 | 0.00 | 0.08 | 0.00 |
| Chengelboden | 0.00 | 15.30 | 55.65 | 0.08 | 0.00 | ||
| Lindi | 0.00 | 1.69 | 3.67 | 0.08 | 0.00 | ||
| Marterloch | 0.00 | 3.01 | 5.84 | 0.08 | 0.00 | ||
| Brengspel | 445.62 | 0.07 | 0.00 | 0.08 | 0.00 | ||
| Hörnli | Irchel | 0.00 | 2.90 | 6.52 | 0.08 | 0.00 | |
| Schartenflue | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 | ||
| Schaffhuser | 606.36 | 5.87 | 2.75 | 0.08 | 0.00 | ||
| Obermeser | 0.00 | 8.68 | 29.91 | 0.08 | 0.00 | ||
| Steig | 0.00 | 17.12 | 54.84 | 0.08 | 0.00 |
| Site | Horizon | Depth [m] | qz | fsp | crb | feo | shs | cm | om |
|---|---|---|---|---|---|---|---|---|---|
| Irchel | |||||||||
| Steig | BcnC | 1.6–1.8 | 55.75 | 0.54 | 0 | 5.17 | 4.14 | 34.56 | 0.09 |
| Hörnli | Sd | 1.8–2.25 | 56.65 | 4.5 | 0 | 5.01 | 2.45 | 32.12 | 0.01 |
| Schartenflue | Rcca | 1.1–1.4 | 58.82 | 5.07 | 18.35 | 1.9 | 5.6 | 10.53 | 0.15 |
| Obermeser | SdC | 1.3–1.6 | 66.44 | 1.24 | 0 | 3.28 | 2.98 | 25.57 | 0.29 |
| Schaffhuser | Cca | 0.9–1.1 | 34.49 | 5.92 | 30.22 | 3.6 | 0 | 26.05 | 0.05 |
| Bülach | |||||||||
| Lärchenischlag | Cca2 | 1.2–1.35 | 48.48 | 14.39 | 26.65 | 0.94 | 5.6 | 3.52 | 0.81 |
| Chengelboden | IICca | 1.95–2.2 | 33.05 | 14.06 | 29.6 | 2.62 | 8.01 | 12.5 | 0.2 |
| Brengspel | Cca2 | 1.0–1.2 | 30.76 | 7.15 | 42.4 | 0.67 | 6.21 | 13.21 | 0.09 |
| Marterloch | Cca | 1.2–1.4 | 50.6 | 6.33 | 23.77 | 1.77 | 9.83 | 8.02 | 0.14 |
| Lindi | B | 0.85–0.95 | 57.46 | 17.44 | 0.2 | 2.09 | 10.54 | 13.06 | 0.16 |
| Full vs. Partial Model | Response Mass Balance | Thinning HIGH | Thinning LOW | Timber Harvesting (Stem Only) | Timber Harvesting (Whole Tree) | Depth of Occurrence of Calcite | Depth of the Rooting Zone | Late Pleistocene | Mean CEC | Mean pH |
|---|---|---|---|---|---|---|---|---|---|---|
| Full | Ca | 0.07 | −0.20 | 0.12 | −0.12 | −0.27 | 1.72 | 0.07 | −0.01 | 1.14 |
| Partial | Ca | 0.07 | −0.20 | 0.12 | −0.12 | −0.94 | 2.33 | −0.21 | −0.01 | − |
| Full | N | 0.03 | −0.10 | 0.13 | −0.27 | −0.45 | 0.97 | 0.16 | −0.00 | −0.18 |
| Partial | N | 0.03 | −0.10 | 0.13 | −0.27 | −0.34 | 0.88 | 0.21 | −0.00 | − |
| Full | K | −0.02 | 0.11 | 0.56 | −0.67 | 3.42 | −1.70 | 6.66 | 0.06 | −0.88 |
| Partial | K | −0.02 | 0.11 | 0.56 | −0.67 | 3.94 | −2.17 | 6.88 | 0.06 | − |
| Full | Mg | −0.01 | 0.04 | 0.18 | −0.15 | 1.09 | −1.12 | 2.23 | 0.02 | 0.72 |
| Partial | Mg | −0.01 | 0.04 | 0.18 | −0.15 | 0.66 | −0.74 | 2.05 | 0.02 | − |
| Full | P | −0.02 | −0.01 | −0.23 | 0.37 | 0.11 | −0.30 | −0.14 | 0.00 | −0.10 |
| Partial | P | −0.02 | −0.01 | −0.23 | 0.37 | 0.17 | −0.35 | −0.11 | 0.00 | − |
| p-Values Are Reported by the Summary() Function in R from t-Tests of Model Coefficients | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Full vs. Partial Model | Response Mass Balance | Thinning HIGH | Thinning LOW | Timber Harvesting (Stem Only) | Timber Harvesting (Whole Tree) | Depth of the Occurrence of Calcite | Depth of the Rooting Zone | Late Pleistocene | Mean CEC | Mean pH |
| Full | Ca | () | * | () | () | * | *** | () | *** | *** |
| Partial | Ca | () | . | () | () | *** | *** | () | ** | - |
| Full | N | () | () | * | *** | *** | *** | () | . | . |
| Partial | N | () | () | * | *** | *** | *** | . | () | - |
| Full | K | () | () | () | () | *** | () | *** | ** | () |
| Partial | K | () | () | () | () | *** | () | *** | ** | - |
| Full | Mg | () | () | () | () | *** | . | *** | *** | ** |
| Partial | Mg | () | () | () | () | *** | () | *** | ** | - |
| Full | P | () | () | *** | *** | * | * | * | () | . |
| Partial | P | () | () | *** | *** | *** | ** | . | () | - |
| p-Values Relate to the Distribution | ||||
|---|---|---|---|---|
| Nutrient Mass Balance | F Statistic | p Value | B-H Corrected Threshold (p) | Decision |
| Calcium | 80.91 | 8.99 × 10−14 | 0.01 | Reject |
| Magnesium | 7.33 | 8.31 × 10−3 | 0.0125 | Reject |
| Phosphorus | 3.82 | 5.42 ×10−2 | 0.017 | Do not reject |
| Nitrogen | 3.62 | 6.06 × 10−2 | 0.025 | Do not reject |
| Potassium | 1.02 | 3.20 × 10−1 | 0.05 | Do not reject |
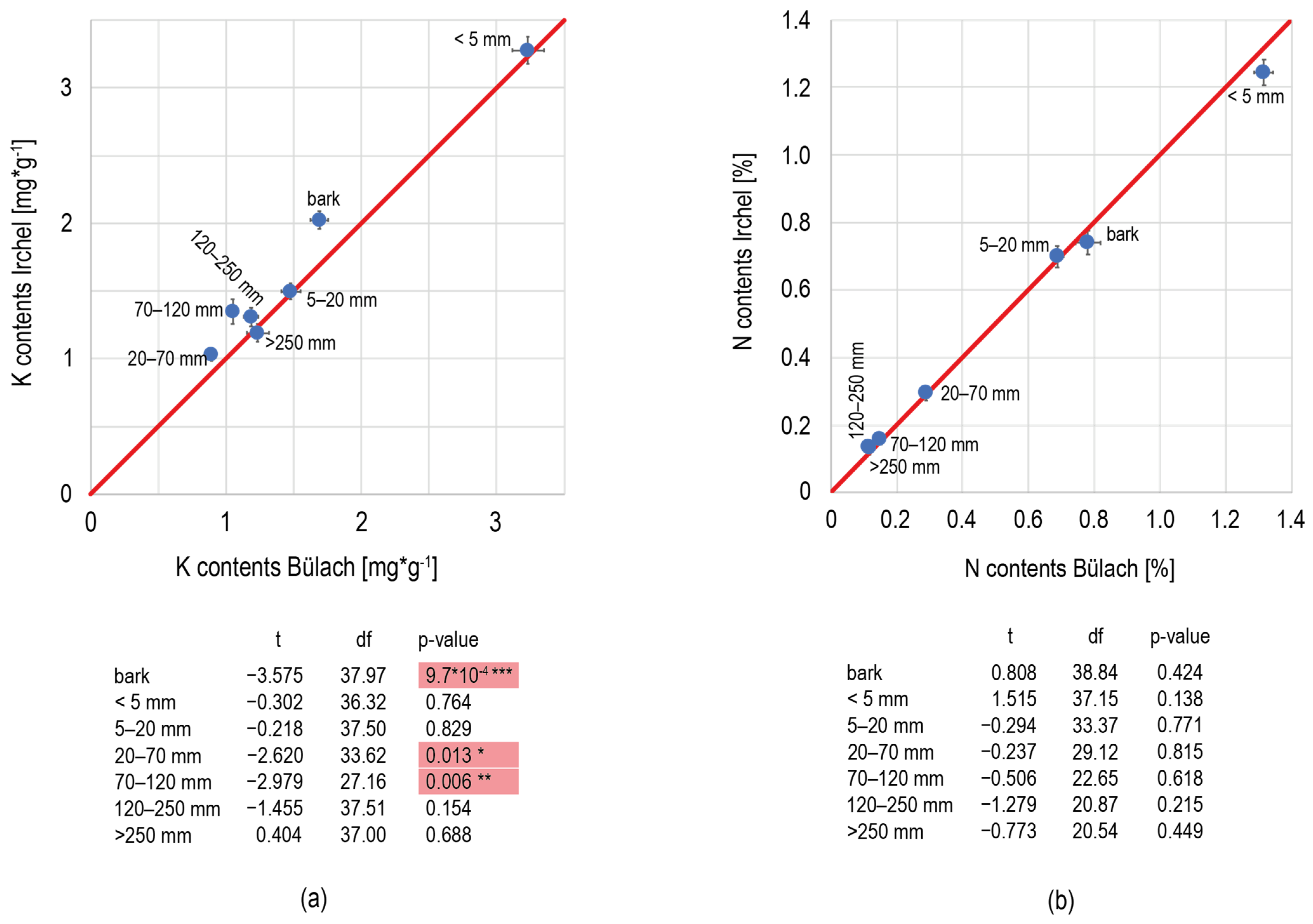
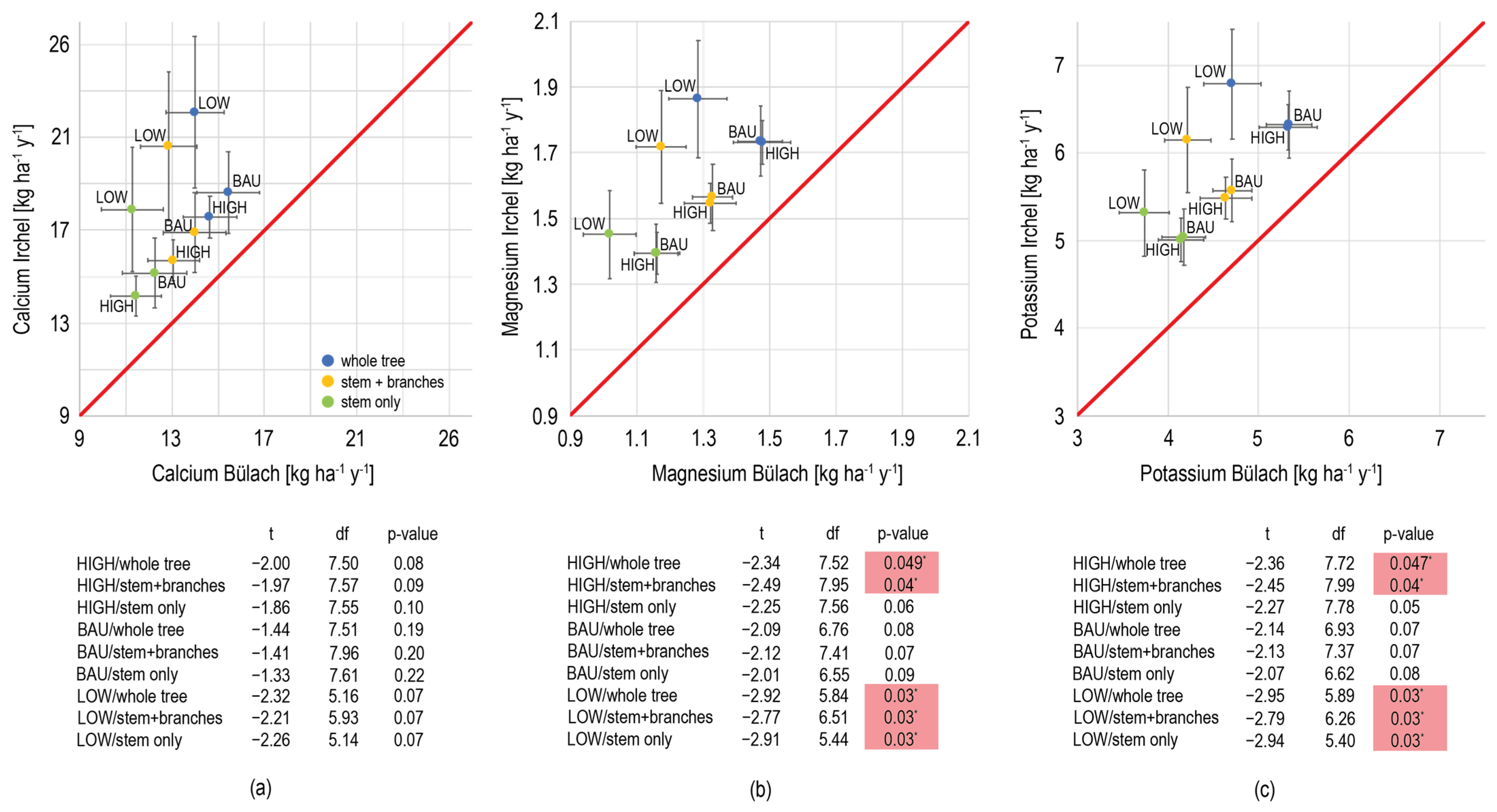

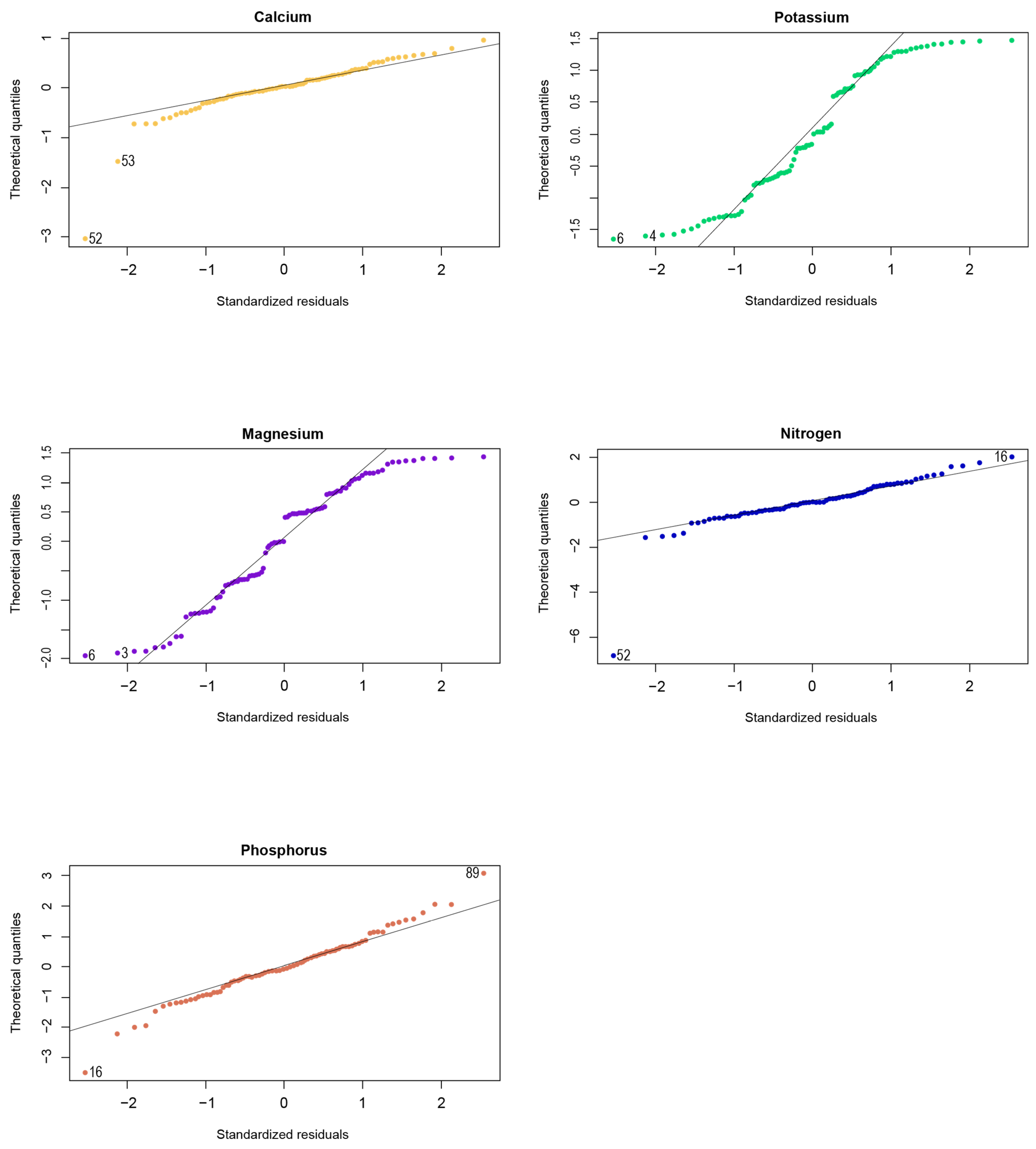
References
- de Jong, A.; de Vries, W.; Kros, H.; Spijker, J. Impacts of harvesting methods on nutrient removal in Dutch forests exposed to high-nitrogen deposition. Ann. For. Sci. 2022, 79, 33. [Google Scholar] [CrossRef]
- Brin, A.; Bouget, C.; Brustel, H.; Jactel, H. Diameter of downed woody debris does matter for saproxylic beetle assemblages in temperate oak and pine forests. J. Insect Conserv. 2011, 15, 653–669. [Google Scholar] [CrossRef]
- Andersson, J.; Hjältén, J.; Dynesius, M. Wood-inhabiting beetles in low stumps, high stumps and logs on boreal clear-cuts: Implications for dead wood management. PLoS ONE 2015, 10, e0118896. [Google Scholar] [CrossRef] [PubMed]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. 2006, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Khanna, P.K.; Fortmann, H.; Meesenburg, H.; Eichhorn, J.; Meiwes, K.J. Biomass and element content of foliage and aboveground litterfall on three long-term experimental beech sites: Dynamics and significance. In Functioning and Management of European Beech Ecosystems; Brumme, R., Khanna, P.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Ecological Studies Volume 208, pp. 183–205. [Google Scholar]
- Rademacher, P.; Khanna, P.K.; Eichhorn, J.; Guericke, M. Tree growth, biomass, and elements in tree components of three beech sites. In Functioning and Management of European Beech Ecosystems; Brumme, R., Khanna, P.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Ecological Studies Volume 208, pp. 105–136. [Google Scholar]
- de Vries, W.; de Jong, A.; Kros, J.; Spijker, J. The use of soil nutrient balances in deriving forest biomass harvesting guidelines specific to region, tree species and soil type in the Netherlands. For. Ecol. Manag. 2021, 479, 118591. [Google Scholar] [CrossRef]
- Kaufmann, G.; Staedeli, M.; Wasser, B. Grundanforderungen an den Naturnahen Waldbau; Projektbericht; Bundesamt für Umwelt (BAFU): Bern, Switzerland, 2010; 42p. [Google Scholar]
- Larsen, J.B.; Angelstam, P.; Bauhus, J.; Carvalho, J.F.; Diaci, J.; Dobrowolska, D.; Gazda, A.; Gustafsson, L.; Krumm, F.; Knoke, T.; et al. Closer-to-Nature Forest Management; European Forest Institute: Joensuu, Finland, 2022; From Science to Policy 12. [Google Scholar] [CrossRef]
- Du, E.; de Vries, W. Nitrogen deposition and its impacts on forest ecosystems: A global perspective. In Atmospheric Nitrogen Deposition to Global Forests. Spatial Variation, Impacts, and Management Implications; Du, E., de Vries, W., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; Chapter 1. [Google Scholar] [CrossRef]
- Braun, S.; Rihm, B.; Tresch, S.; Schindler, C. Long-term risk assessment of uprooting and stem breakage under drought conditions and at high N deposition in beech and Norway spruce. Agric. For. Meteorol. 2023, 341, 109669. [Google Scholar] [CrossRef]
- Turner, B.L.; Laliberté, E. Soil Development and Nutrient Availability Along a 2 Million-Year Coastal Dune Chronosequence Under Species-Rich Mediterranean Shrubland in Southwestern Australia. Ecosystems 2015, 18, 287–309. [Google Scholar] [CrossRef]
- Duchesne, L.; Houle, D. Base cation cycling in a pristine watershed of the Canadian boreal forest. Biogeochemistry 2006, 78, 195–216. [Google Scholar] [CrossRef]
- Haldimann, P.; Graf, H.R.; Jost, J. Geological Atlas of Switzerland, Map No. 151 with Explanatory Notes; Federal Office of Topography, Swiss Geological Survey: Bern, Switzerland, 2017. [Google Scholar]
- Keller, W.; Wohlgemuth, T.; Kuhn, N.; Schütz, M.; Wildi, O. Classification of Swiss forest vegetation with floristic data. Statistically revised version of “Waldgesellschaften und Waldstandorte der Schweiz” by Heinz Ellenberg and Frank Klötzli (1972). Mitt. Eidgenöss. Forsch. Wald Schnee Landsch. 1998, 73, 91–357. [Google Scholar]
- Remund, J. Downscaling CH2018. Calculation of Meteorological and Drought Indices for Forest Research Methods and Results (Version 2); Report for the Federal Office for the Environment; METEOTEST: Bern, Germany, 2020. [Google Scholar]
- Thomas, G.W. Exchangeable Cations. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; Volume 9, pp. 159–165. [Google Scholar]
- Walthert, L.; Graf, U.; Kammer, A.; Luster, J.; Pezzotta, D.; Zimmermann, S.; Hagedorn, F. Determination of organic and inorganic carbon, ∂13C, and nitrogen in soils containing carbonates after acid fumigation with HCl. J. Plant Nutr. Soil Sci. 2010, 173, 207–216. [Google Scholar] [CrossRef]
- Lakanen, E.; Erviö, R. A comparison of eight extractants for the determination of plant available micronutrients in soils. Acta Agral. Fenn. 1971, 123, 223–232. [Google Scholar]
- Posch, M.; Kurz, D. A2M A program to compute all possible mineral modes from geochemical analyses. Comput. Geosci. 2007, 33, 563–572. [Google Scholar] [CrossRef]
- Deer, W.A.; Howie, R.A.; Zussman, J. Rock-Forming Minerals, Vol. 3, SheetSilicates; John Wiley: New York, NY, USA, 1962; 270p. [Google Scholar]
- Deer, W.A.; Howie, R.A.; Zussman, J. An Introduction to the Rock-Forming Minerals; Longman Group Limited: London, UK, 1980; 528p. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle-size Analysis. In Methods of Soil Analysis, Part 1, Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Soil Science Society of America: Madison, WI, USA, 1986; pp. 383–423. [Google Scholar]
- Rihm, B.; Achermann, B. Critical Loads of Nitrogen and Their Exceedances. Swiss Contribution to the Effects-Oriented Work under the Convention on Long-Range Transboundary Air Pollution (UNECE); Federal Office for the Environment: Bern, Switzerland, 2016; 78p, Environmental studies no. 1642. [Google Scholar]
- Rihm, B.; Künzle, T. Mapping Nitrogen Deposition 2015 for Switzerland. Technical Report on the Update of Critial Loads and Exceedance, including the Years 1990, 2000, and 2010; METEOTEST 29.01.2019; Federal Office for the Environment: Bern, Germany, 2019; Available online: https://www.bafu.admin.ch/dam/bafu/en/dokumente/luft/externe-studien-berichte/mapping-nitrogen-deposition-2015-for-switzerland.pdf (accessed on 18 June 2014).
- Thimonier, A.; Schmitt, M.; Waldner, P.; Rihm, B. Atmospheric deposition on Swiss Long-Term Forest Ecosystem Research (LWF) plots. Environ. Monit. Assess. 2005, 104, 81–118. [Google Scholar] [CrossRef] [PubMed]
- NABEL. Technischer Bericht zum Nationalen Beobachtungsnetz für Luftfremdstoffe (NABEL); Federal Office for the Environment, Bern, Materials Science and Technology; EMPA: Bern, Germany, 2023; 200p. [Google Scholar]
- Rihm, B.; Thimonier, A.; Albrecht, S.; Waldner, P. Zwischenbericht Berechnung der Deposition basischer Kationen für Wälder, Provisorische Depositionskarten für Ca, Mg, K, Na und Cl; Internal project report Meteotest/WSL dated 28.5.2013; Federal Office for the Environment, Forest Division: Bern, Switzerland, 2013; 22p. [Google Scholar]
- Posch, M. SWWm A Program to Compute Weathering Rates for a Multilayer Soil Profile. User Manual; Version 2.15; IIASA: Laxenburg, Austria, 2022. [Google Scholar]
- Sverdrup, H.U. The Kinetics of Base Cation Release Due to Chemical Weathering; Lund University Press: Lund, Sweden, 1990; ISBN 0-86238-247-5. [Google Scholar]
- Sverdrup, H.U.; Warfvinge, P. Calculating field weathering rates using a mechanistic geochemical model PROFILE. Appl. Geochem. 1993, 8, 273–283. [Google Scholar] [CrossRef]
- Sverdrup, H.U.; Warfvinge, P. Estimating field weathering rates using laboratory kinetics. In Chemical Weathering of Silicate Minerals; White, A.F., Brantley, S.L., Eds.; Mineralogical Society of America: Chantilly, VA, USA, 1995; Reviews in Mineralogy 31; pp. 485–541. [Google Scholar]
- Alveteg, M. Dynamics of Forest Soil Chemistry. Ph.D. Thesis, Department of Chemical Engineering II, Lund University, Lund, Sweden, 1998. [Google Scholar]
- Posch, M.; De Vries, W.; Sverdrup, H.U. Mass balance models to derive critical loads of nitrogen and acidity for terrestrial and aquatic ecosystems. In Critical Loads and Dynamic Risk Assessments: Nitrogen, Acidity and Metals in Terrestrial and Aquatic Ecosystems; de Vries, W., Hettelingh, J.P., Posch, M., Eds.; Environmental Pollution Series; Springer: Dordrecht, The Netherland, 2015; Volume 25, xxviii+662p, ISBN 978-94-017-9507-4. [Google Scholar] [CrossRef]
- UNECE. Manual on Methodologies and Criteria for Modelling and Mapping Critical Loads and Levels and Air Pollution Effects, Risks and Trends; UNECE Convention on Long-range Transboundary Air Pollution (LRTAP): Geneva, Switzerland, 2004; 266p, Available online: http://icpmapping.org/Mapping_Manual (accessed on 10 June 2024).
- Posch, M.; Reinds, G.J. A very simple dynamic soil acidification model for scenario analyses and target load calculations. Environ. Model. Softw. 2009, 24, 329–340. [Google Scholar] [CrossRef]
- Zell, J. A Climate Sensitive Single Tree Stand Simulator for Switzerland (SwissStandSim); Swiss Federal Institute of Forest, Snow and Landscape Research WSL: Birmensdorf, Switzerland, 2016; 107p, Available online: www.wsl.ch/wald_klima (accessed on 10 June 2024).
- Mey, R.; Stadelmann, G.; Thürig, E.; Bugmann, H.; Zell, J. From small forest samples to generalised uni- and bimodal stand descriptions. Methods Ecol. Evol. 2021, 12, 634–645. [Google Scholar] [CrossRef]
- Didion, M.; Herold, A.; Thürig, E. Whole tree biomass and carbon stock. In Managing Forest Ecosystems Swiss National Forest Inventory Methods and Models of the Fourth Assessment; Fischer, C., Traub, B., Eds.; Springer: Cham, Switzerland, 2019; Volume 35, pp. 243–248. [Google Scholar] [CrossRef]
- Herold, A.; Zell, J.; Rohner, B.; Didion, M.; Thürig, E.; Rösler, E. State and change of forest resources. In Managing Forest Ecosystems Swiss National Forest Inventory Methods and Models of the Fourth Assessment; Fischer, C., Traub, B., Eds.; Springer: Cham, Switzerland, 2019; Volume 35, pp. 205–230. [Google Scholar] [CrossRef]
- Thrippleton, T.; Blattert, C.; Bont, L.G.; Mey, R.; Zell, J.; Thürig, E.; Schweier, J. A multi-criteria decision support system for strategic planning at the Swiss forest enterprise level: Coping with climate change and shifting demands in ecosystem service provisioning. Front. For. Glob. Chang. 2021, 4, 1–18. [Google Scholar] [CrossRef]
- Zell, J.; Nitzsche, J.; Stadelmann, G.; Thürig, E. SwissStandSim: Ein klimasensitives, einzelbaumbasiertes Waldwachstumsmodell. Schweiz. Z. Für Forstwes. 2020, 171, 116–123. [Google Scholar] [CrossRef]
- Forrester, D.I.; Nitzsche, J.; Schmid, H. The Experimental Forest Management project: An Overview and Methodology of the Long-Term Growth and Yield Plot Network; Swiss Federal Institute of Forest, Snow and Landscape Research WSL: Birmensdorf, Switzerland, 2019; 73p, Available online: https://www.wsl.ch/fileadmin/user_upload/WSL/Wald/Waldentwicklung_Monitoring/EFM-EK/forrester_etal_2019_efm_new.pdf (accessed on 10 June 2024).
- Zell, J. SwissStandSim: A Climate Sensitive Single Tree Stand Simulator for Switzerland; Swiss Federal Institute of Forest, Snow and Landscape Research WSL: Birmensdorf, Switzerland, 2018; Schlussbericht im Forschungsprogramm Wald und Klimawandel; Available online: https://www.research-collection.ethz.ch/handle/20.500.11850/311156 (accessed on 10 June 2024).
- Mey, R.; Zell, J.; Thürig, E.; Stadelmann, G.; Bugmann, H.; Temperli, C. Tree species admixture increases ecosystem service provision in simulated spruce- and beech dominated stands. Eur. J. For. Res. 2022, 141, 801–820. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing: Vienna, Austria, 2021. Available online: https://www.R-project.org/(accessed on 10 June 2024).
- Welch, B.L. The generalization of “Student’s” problem when several different population variances are involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Crews, T.E.; Kitayama, K.; Fownes, J.H.; Riley, R.H.; Herbert, D.A.; MuellerDombois, D.; Vitousek, P.M. Changes in Soil Phosphorus Fractions and Ecosystem Dynamics across a Long Chronosequence in Hawaii. Ecology 1995, 76, 1407–1424. [Google Scholar] [CrossRef]
- Richardson, S.J.; Peltzer, D.A.; Allen, R.B.; Mcglone, M.S.; Parfitt, R.L. Rapid development of phosphorus limitation in temperate rainforest along the Franz Josef soil chronosequence. Oecologia 2014, 139, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Huang, L.-M. Changes in soil nutrients and stoichiometric ratios reveal increasing phosphorus deficiency along a tropical soil chronosequence. Catena 2023, 222, 106893. [Google Scholar] [CrossRef]
- Jacobsen, C.; Rademacher, P.; Meesenburg, H.; Meiwes, K.J. Gehalte chemischer Elemente in Baumkompartimenten Literaturstudie und Datensammlung. Research Center Forest Ecosystems, University of Göttingen: Göttingen, Germany, 2003; Volume 69. [Google Scholar]
- Meiwes, K.-J.; Beese, F. Ergebnisse der Untersuchung des Stoffhaushaltes eines Buchenwaldökosystems auf Kalkgestein. Ber. Forschungszentr. Wald. 1988, 9, 1–141. [Google Scholar]
- Göttlein, A.; Baumgarten, M.; Dieler, J. Site Conditions and Tree-Internal Nutrient Partitioning in Mature European Beech and Norway Spruce at the Kranzberger Forest. In Growth and Defense in Plants; Matyssek, R., Schnyder, H., Osswald, W., Ernst, D., Munch, J.C., Pretzsch, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 220, pp. 193–211. [Google Scholar]
- Ulbricht, M.; Göttlein, A.; Biber, P.; Dieler, J.; Pretzsch, H. Variations of nutrient concentrations and contents between summer and autumn within tree compartments of European beech (Fagus sylvatica). J. Plant Nutr. Soil Sci. 2016, 179, 746–757. [Google Scholar] [CrossRef]
- Calvaruso, C.; Kirchen, G.; Saint-André, L.; Redon, P.-O.; Turpault, M.-P. Relationship between soil nutritive resources and the growth and mineral nutrition of a beech (Fagus sylvatica) stand along a soil sequence. Catena 2017, 155, 156–169. [Google Scholar] [CrossRef]
- Diez, C.; Bürgi, A. Wuchsleistung und Qualität von Douglasie (Pseudotsuga menziesii [Mirbel] Franco), Riesen-Lebensbaum (Thuja plicata Donn) und Roteiche (Quercus rubra L.) in der Schweiz. Ber. Eidg. Forschungsanst. WSL 1991, 329, 46. [Google Scholar]
- Thurm, E.A.; Pretzsch, H. Improved productivity and modified tree morphology of mixed versus pure stands of European beech (Fagus sylvatica) and Douglas-fir (Pseudotsuga menziesii) with increasing precipitation and age. Ann. For. Sci. 2016, 73, 1047–1061. [Google Scholar] [CrossRef]
- Thimonier, A.; Graf Pannatier, E.; Schmitt, M.; Waldner, P.; Walthert, L.; Schleppi, P.; Dobbertin, M.; Kräuchi, N. Does exceeding the critical loads for nitrogen alter nitrate leaching, the nutrient status of trees and their crown condition at Swiss Long-term Forest Ecosystem Research (LWF) sites? Eur. J. Forest Res. 2010, 129, 443–461. [Google Scholar] [CrossRef]
- Waldner, P.; Braun, S.; Brunner, I.; Rihm, B.; Reinhard, M.; Hajjar, N.; Meusburger, K.; Schmitt, M.; Thimonier, A. Stickstoff-Deposition in Schweizer Wälder und Nitrataustrag aus Waldböden. Forum für Wissen 2022. Ber. Eidg. Forschungsanst. WSL 2022, 126, 47–55. [Google Scholar] [CrossRef]
- Larsen, J.B. Ecological stability of forests and sustainable silviculture. For. Ecol. Manag. 1995, 73, 85–96. [Google Scholar] [CrossRef]


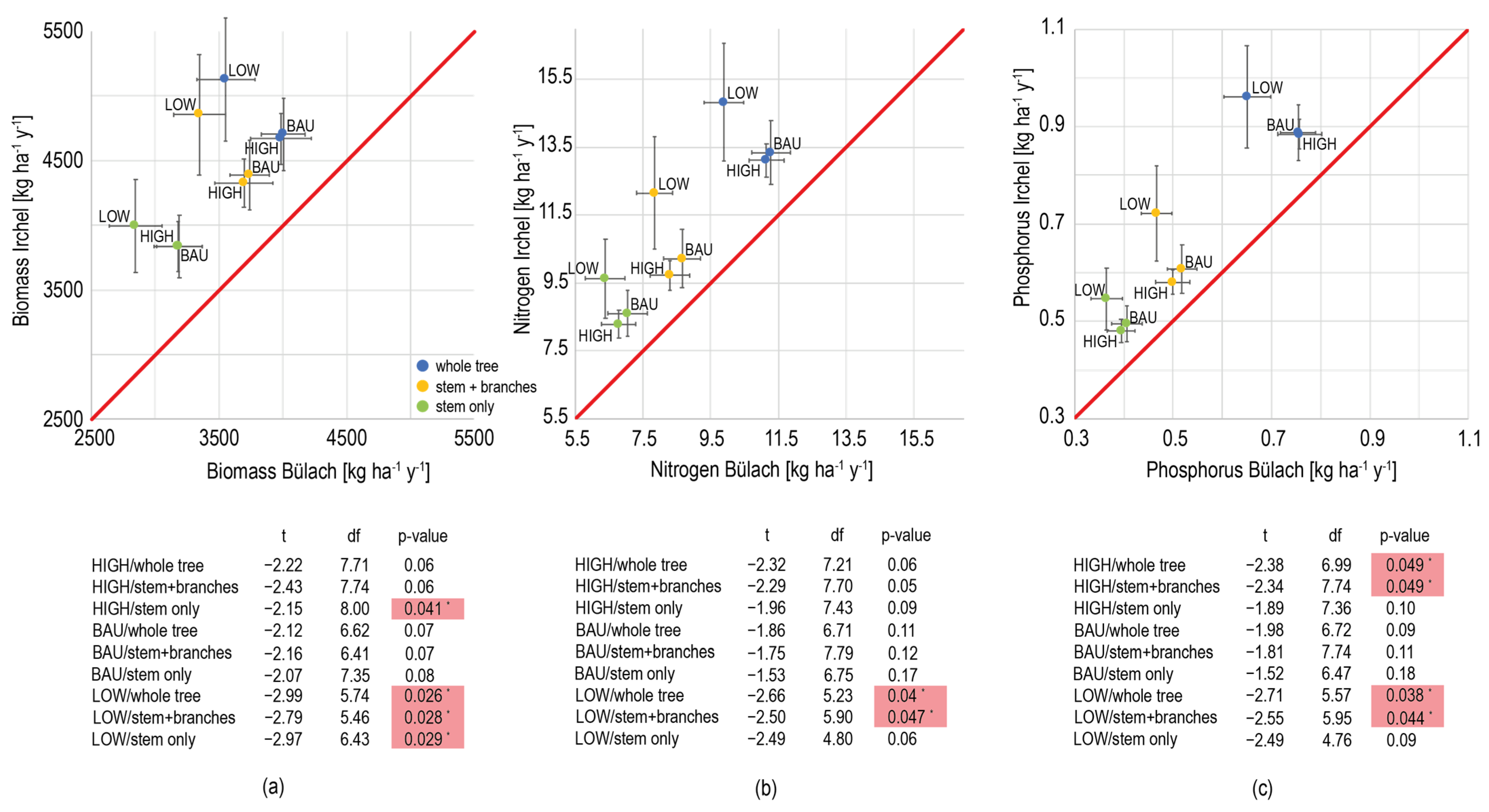


| Geological Unit | Site Name | Depth of Rooting Zone [m] | Depth of the Occurrence of Calcite [m] | Mean Annual Precipitation [mm] | Mean Annual Temperature [°C] | Radiation [µmol photons·m−2·s−1] |
|---|---|---|---|---|---|---|
| Early Pleistocene | Hörnli | 0.90 | 2.45 | 1504 | 8.7 | 690 |
| Schartenflue | 0.60 | 1.00 | 1507 | 8.7 | 689 | |
| Schaffhuser | 1.20 | 0.80 | 1396 | 8.9 | 685 | |
| Obermeser | 1.30 | 1.90 | 1523 | 8.7 | 690 | |
| Steig | 2.00 | 2.20 | 1514 | 8.7 | 689 | |
| Mean ± std. error | 1.2 ± 0.2 | 1.7 ± 0.3 | 1489 ± 19 | 8.7 ± 0.04 | 689 ± 0.7 | |
| Late Pleistocene | Lärchenischlag | 1.10 | 0.80 | 993 | 9.6 | 675 |
| Chengelboden | 1.40 | 1.95 | 1005 | 9.6 | 677 | |
| Lindi | 1.00 | 1.00 | 1001 | 9.6 | 676 | |
| Marterloch | 1.05 | 1.05 | 1008 | 9.6 | 677 | |
| Brengspel | 1.25 | 0.40 | 1004 | 9.6 | 677 | |
| Mean ± std. error | 1.2 ± 0.1 | 1.0 ± 0.3 | 1002 ± 2 | 9.6 ± 0 | 676 ± 0.2 |
| Soil Property | Bülach (Mean ± std. Error) | Irchel (Mean ± std. Error) | t | df | p-Value |
|---|---|---|---|---|---|
| Mean pH value | 4.04 ± 0.24 | 3.90 ± 0.18 | 0.46 | 7.33 | 0.66 |
| Mean BS [%] | 54.4 ± 12.3 | 46.7 ± 12.4 | 0.44 | 8.00 | 0.67 |
| CEC [molc·m−2] | 68.5 ± 20.2 | 159.8 ± 52.9 | −1.61 | 5.14 | 0.17 |
| Clay fraction [kg·m−2] | 175.8 ± 68.7 | 343.5 ± 126.2 | −1.17 | 6.18 | 0.29 |
| Stock of Corg [kg·m−2] | 6.4 ± 0.5 | 6.7 ± 0.8 | −0.29 | 7.09 | 0.78 |
| Caexc [mol·m−2] | 14.6 ± 5.7 | 32.2 ± 20.3 | −0.83 | 4.63 | 0.45 |
| Mgexc [mol·m−2] | 3.6 ± 2.4 | 8.1 ± 4.0 | −0.96 | 6.54 | 0.37 |
| Kexc [mol·m−2] | 1.3 ± 0.6 | 2.6 ± 0.8 | −1.26 | 7.54 | 0.24 |
| Plak [mmol·m−2] | 1133 ± 669 | 195.3 ± 76.1 | 1.39 | 4.10 | 0.23 |
| Ntot [mol·m−2] | 40.4 ± 8.7 | 43.0 ± 10.1 | −0.19 | 7.84 | 0.85 |
| Nutrient Flux | Bülach (Mean ± std. Error) [kg·ha−1·y−1] | Irchel (Mean ± std. Error) [kg·ha−1·y−1] | t | df | p-Value |
|---|---|---|---|---|---|
| Ca_weath | 116.3 ± 57.9 | 126.3 ± 66.5 | −0.11 | 27.48 | 0.91 |
| Mg_weath | 6.6 ± 1.8 | 9 ± 1.7 | −0.96 | 27.97 | 0.34 |
| K_weath | 21.1 ± 6.6 | 26.1 ± 6.1 | −0.55 | 27.83 | 0.59 |
| Ca_leach Mg_leach | 89.4 ± 47.5 | 121.3 ± 64.8 | −0.40 | 25.68 | 0.69 |
| 4 ± 1.5 | 6.9 ± 1.6 | −1.29 | 28.00 | 0.21 | |
| K_leach | 13.2 ± 5.7 | 18.7 ± 5.6 | −0.69 | 27.97 | 0.49 |
| Ca_depo | 6.3 ± 0 | 6.2 ± 0 | 1.27 | 6.74 | 0.25 |
| Mg_depo | 0.7 ± 0 | 0.7 ± 0 | 1.01 | 6.80 | 0.35 |
| K_depo | 2.7 ± 0 | 2.7 ± 0 | 2.34 | 6.74 | 0.053 |
| N_depo | 23.9 ± 0.6 | 20 ± 0.5 | 4.80 | 7.83 | 0.001 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimmermann, S.; Kurz, D.; Thrippleton, T.; Mey, R.; Perry, N.T.; Posch, M.; Schweier, J. Does Soil Acidification Matter? Nutrient Sustainability of Timber Harvesting in Forests on Selected Soils Developed in Sediments of the Early vs. Late Pleistocene. Forests 2024, 15, 1079. https://doi.org/10.3390/f15071079
Zimmermann S, Kurz D, Thrippleton T, Mey R, Perry NT, Posch M, Schweier J. Does Soil Acidification Matter? Nutrient Sustainability of Timber Harvesting in Forests on Selected Soils Developed in Sediments of the Early vs. Late Pleistocene. Forests. 2024; 15(7):1079. https://doi.org/10.3390/f15071079
Chicago/Turabian StyleZimmermann, Stephan, Daniel Kurz, Timothy Thrippleton, Reinhard Mey, Niál Thomas Perry, Maximilian Posch, and Janine Schweier. 2024. "Does Soil Acidification Matter? Nutrient Sustainability of Timber Harvesting in Forests on Selected Soils Developed in Sediments of the Early vs. Late Pleistocene" Forests 15, no. 7: 1079. https://doi.org/10.3390/f15071079
APA StyleZimmermann, S., Kurz, D., Thrippleton, T., Mey, R., Perry, N. T., Posch, M., & Schweier, J. (2024). Does Soil Acidification Matter? Nutrient Sustainability of Timber Harvesting in Forests on Selected Soils Developed in Sediments of the Early vs. Late Pleistocene. Forests, 15(7), 1079. https://doi.org/10.3390/f15071079