
Physiological Response to Low-Temperature Stress and Cold Resistance Evaluation of Ziziphus jujuba var. spinosa Clones from Different Provenances


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Treatment
2.3. Experimental Methods
2.3.1. Measurements of Physiological Indices
2.3.2. Observation of Anatomical Structure
2.4. Statistical Analyses
3. Results
3.1. Response of Branch Membrane Stability to Low-Temperature Stress
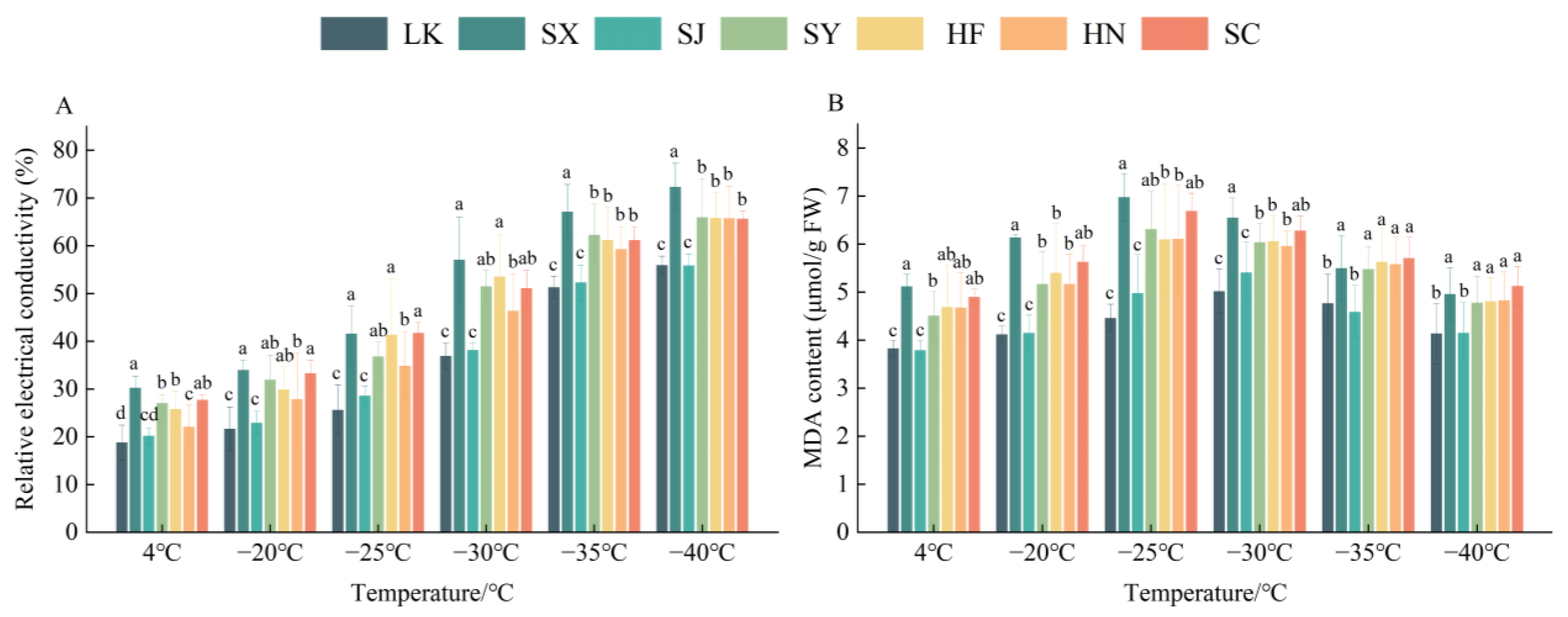
3.1.1. Relative Electrical Conductivity
3.1.2. Malondialdehyde Content
3.2. Response of Branch Water Content to Low-Temperature Stress
3.2.1. Total Water Content
3.2.2. Bound Water/Free Water Ratio
3.3. Response of Antioxidant Enzyme Activities of Branches to Low-Temperature Stress
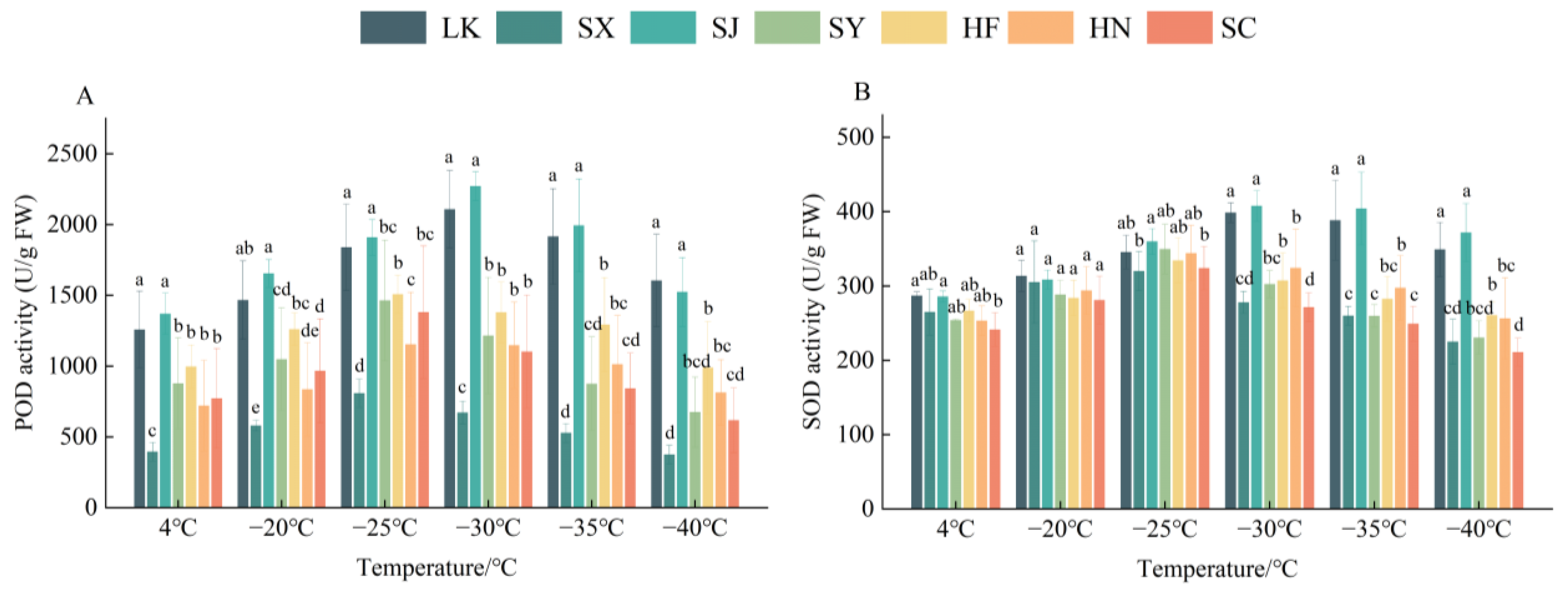
3.3.1. POD Activity
3.3.2. SOD Activity
3.4. Response of Osmotic Regulatory Substances of Branches to Low-Temperature Stress
3.4.1. Soluble Protein Content
3.4.2. Soluble Sugar Content
3.4.3. Proline Content
3.4.4. Starch Content
3.5. Branch Anatomical Structure
3.6. Comprehensive Evaluation of Cold Resistance
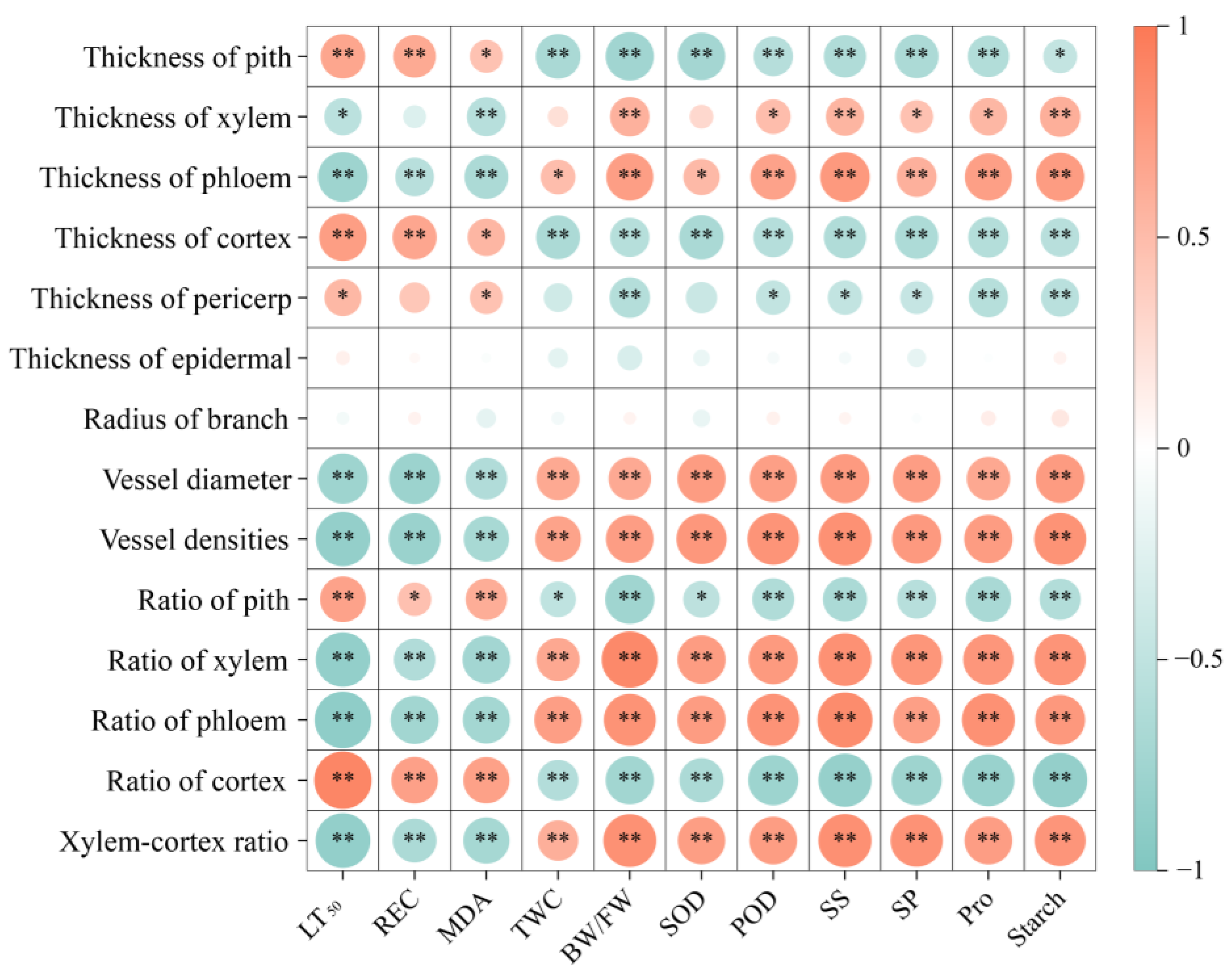
3.6.1. Correlation Analysis
3.6.2. Membership Function Analysis
3.6.3. Cluster Heat Map Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.J.; Li, P.; Yu, G.D. Advance of research and prospect of development of Zizyphus jujuba var. spinosa (Bunge) Hu ex HF Chow. Chin. Wild Plant Resour. 1999, 18, 17–21. [Google Scholar]
- Zhang, Y.; Guo, S.; Zhu, S.Q.; Xu, L.; Yan, H.; Qian, D.W.; Duan, J.A. Effect of different drying methods on nucleoside, amino acids and flavonoids in the leaves of Ziziphus jujuba var. spinosa. Sci. Technol. Food Ind. 2016, 37, 296–303. [Google Scholar]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Gong, M.; Xin, H.; Tang, L.Z.; Dai, D.Q.; Gao, Y.; Liu, C. Effects of chilling stress on the accumulation of soluble sugars and their key enzymes in Jatropha curcas seedlings. Physiol. Mol. Biol. Plants 2018, 24, 857–865. [Google Scholar] [CrossRef]
- Sun, S.H.; Qi, X.J.; Wang, R.; Lin, M.M.; Fang, J.B. Evaluation of freezing tolerance in Actinidia germplasm based on relative electrolyte leakage. Hortic. Environ. Biotechnol. 2020, 61, 755–765. [Google Scholar] [CrossRef]
- Karami, H.; Rezaei, M.; Sarkhosh, A.; Rahemi, M.; Jafari, M. Cold hardiness assessment in seven commercial fig cultivars (Ficus carica L.). Gesunde Pflanz. 2018, 70, 195–203. [Google Scholar] [CrossRef]
- Wang, Y.X.; Hu, Y.; Chen, B.H.; Zhu, Y.F.; Dawuda, M.M.; Svetla, S. Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks. J. Integr. Agric. 2018, 17, 857–866. [Google Scholar] [CrossRef]
- Wang, C.Z.; Gao, J.C.; Li, X.G.; Gao, W.H.; Xin, W.J.; Wu, P. Study on cold hardiness of major jujube cultivars in northwestern China. J. Fruit. Sci. 2011, 28, 898–902. [Google Scholar]
- Gao, M.X.; Tian, X.W.; Zong, J.Y. Research on the Cold Resistance of Different Varietal Jujubes. North. Hortic. 2009, 12, 102–104. [Google Scholar]
- Zhang, Z.P.; Gu, Y.Y.; Mao, Q.X.; Wang, J. Physiological Response to Low Temperature of Four Genotypes of Cyclocarya paliurus and Their Preliminary Evaluation to Cold Resistance. Forests 2023, 14, 1680. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Jiang, C.K.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G.X. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Wang, H.P.; Sun, W.T.; Dong, T.; Niu, J.Q.; Ma, M. Cold resistance evaluation of the shoots of 5 apple rootstocks. J. Fruit. Sci. 2021, 38, 1264–1274. [Google Scholar]
- Lei, M.Y.; Gao, X.F.; Bai, Q.M.; Deng, K.; Zuo, W.F.; Li, Y.Y. Evaluation of Cold Resistance of Pomegranate Branches from Different Varieties. J. Henan Agric. Sci. 2023, 52, 120–130. [Google Scholar]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a multifaceted signalling molecule in plant responses to abiotic stress: Understanding the physiological mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Seydel, C.; Kitashova, A.; Fürtauer, L.; Nägele, T. Temperature-induced dynamics of plant carbohydrate metabolism. Physiol. Plant 2022, 174, e13602. [Google Scholar] [CrossRef] [PubMed]
- Janmohammadi, M.; Zolla, L.; Rinalducci, S. Low temperature tolerance in plants: Changes at the protein level. Phytochemistry 2015, 117, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Lee, Y.M.; Park, J.H.; Nawkar, G.M.; Oh, H.T.; Kim, M.G.; Lee, S.I.; Kim, W.Y.; Yun, D.J.; Lee, S.Y. Ribosomal P3 protein AtP3B of Arabidopsis acts as both protein and RNA chaperone to increase tolerance of heat and cold stresses. Plant Cell Environ. 2016, 39, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Buchner, O.; Neuner, G. Winter frost resistance of Pinus cembra measured in situ at the alpine timberline as affected by temperature conditions. Tree Physiol. 2011, 31, 1217–1227. [Google Scholar] [CrossRef]
- Cheng, J.Y.; Zheng, J.J.; Dou, T.X.; Deng, X.Z.; Wang, W. Summary of Physiological Characteristics of Plant Cold Resistance. Hubei For. Sci. Technol. 2017, 46, 16–20. [Google Scholar]
- Guo, X.M.; Liu, J.Z.; Zhai, J.T.; Xiao, X.; Lv, Y.M.; Li, D.D.; Pei, S.M.; Zhang, L.B. Relationship between leaf anatomical structure and trunk cold resistance of 16 peach cultivars. Sci. Silvae Sin. 2015, 51, 33–43. [Google Scholar]
- Tang, L.H.; Zhao, X.M.; Chai, Y.; Zhao, R.P.; Yang, X.L. Study on the relationship between stem structure and cold resistance of different species of Paeonia rockii. North. Hortic. 2016, 12, 62–64. [Google Scholar]
- Wang, H.; Zhang, J.Z.; Xie, Z.S.; Qi, Y.S. Morphological observation on root vessel elements of different cold resistance grape cultivars. Chin. Agric. Sci. Bull. 2013, 28, 110–114. [Google Scholar]
- Macdonald, M.T.; Lada, R.R.; Veitch, R.S.; Thiagarajan, A.; Adams, A.D. Postharvest needle abscission resistance of balsam fir (Abies balsamea) is modified by harvest date. Can. J. For. Res. 2014, 44, 1394–1401. [Google Scholar] [CrossRef]
- Xu, L.; Tang, Y.; Wang, X.J. Cold Resistance Experiment on Ziziphus jujuba var. spinosa Seedlings with Different Provenances, Tree Ages and Plant Types. Hubei Agric. Sci. 2013, 52, 1325–1329. [Google Scholar]
- Zheng, Q.Q.; Li, P.C.; Chen, Q.L.; Wang, J.J. Comprehensive evaluation of physiological indicators of resistance in Zizphus spinosus Hu from different origins under natural cooling conditions. Jiangsu Agric. Sci. 2019, 47, 154–158. [Google Scholar]
- Zhou, W.J.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 26, 41–47. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Hu, Y.G.; Li, P.P. Consistency of electrical and physiological properties of tea leaves on indicating critical cold temperature. Biosyst. Eng. 2017, 159, 89–96. [Google Scholar] [CrossRef]
- Chen, B.H.; Zhang, B.; Mao, J.; Hao, Y.; Yang, R.; Cai, X.H.; Qi, S.Y. The Relationship between the Changing of Water Content and the Cold Resistance of Grape Branches. Plant Physiol. J. 2014, 50, 535–541. [Google Scholar]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Xiu, W.Y.; Zhu, Y.; Chen, B.; Hu, Y.; Dawuda, M.M. Effects of paclobutrazol on the physiological characteristics of Malus halliana Koehne Seedlings under drought stress via principal component analysis and membership function analysis. Arid. Land. Res. Manag. 2019, 33, 97–113. [Google Scholar] [CrossRef]
- Chen, M.H.; Feng, Y.Q.; Jing, S.C.; Fan, J.B.; Yan, X.B. Evaluation of Semi-lethal Temperature and Cold Toleranceof Bermudagrass (Cynodon dactylon) on Latitude and Longitude Gradients. Chin. J. Grassl. 2022, 44, 49–57. [Google Scholar]
- Yang, Z.X.; Wang, S.Z.; Han, Y.L. Cold Tolerance Variation of Populus cathayana Clones from Different Populations. Forest Res. 1996, 5, 36–41. [Google Scholar]
- Polle, A.; Otter, T.; Seifert, F. Apoplastic peroxidases and lignification in needles of Norway spruce (Picea abies L.). Plant Physiol. 1994, 106, 53–60. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Zhu, Q.Y.; Zhang, M.; Guo, Z.H.; Yang, J.J.; Mo, J.X.; Cui, J.B.; Hu, H.L.; Xu, J. Individual Cryptomeria fortunei Hooibrenk Clones Show Varying Degrees of Chilling Stress Resistance. Forests 2020, 11, 189. [Google Scholar] [CrossRef]
- Chen, L.J.; Xiang, H.Z.; Miao, Y.; Zhang, L.; Guo, Z.F.; Zhao, X.H.; Lin, J.W.; Li, T.L. An Overview of Cold Resistance in plants. J. Agron. Crop Sci. 2014, 200, 237–245. [Google Scholar] [CrossRef]
- Punia Bangar, S.; Trif, M.; Ozogul, F.; Kumar, M.; Chaudhary, V.; Vukic, M.; Tomar, M.; Changan, S. Recent developments in cold plasma-based enzyme activity (browning, cell wall degradation, and antioxidant) in fruits and vegetables. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1958–1978. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y. Injury of Membrane Lipid Peroxidation to Plant Cell. Plant Physiol. Commun. 1991, 2, 84–90. [Google Scholar]
- Guo, Y.L.; Mu, D.S.; Zhao, L.X.; Zhang, Z.M.; Zhang, L.N.; Ma, Z.H. Evaluation of cold resistance of 18 wine grape clones introduced from abroad. J. Arid. Land. Resour. Environ. 2023, 37, 161–168. [Google Scholar]
- Gu, K.; Hou, S.; Chen, J.; Guo, J.; Wang, F.; He, C.; Zou, C.; Xie, X. The physiological response of different tobacco varieties to chilling stress during the vigorous growing period. Sci. Rep. 2021, 11, 22136. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Chang, F.Y.; Zhang, L.Y.; Dong, Q.L.; Luan, H.A.; Jia, P.; Qi, G.H.; Guo, S.P.; Zhang, X.M. The anatomical structure character of raspberry stems is a key factor affecting its cold resistance. Flora 2023, 298, 152196. [Google Scholar] [CrossRef]
- Xu, Y.; Xue, L.; Qu, M. Physiological and Ecological Mechanisms of Plant Adaptation to Low Temperature. Sci. Silvae Sin. 2007, 4, 88–94. [Google Scholar]
- Wang, Z.Y.; Tian, Q.H.; Chang, R.F.; Liu, G.J.; Chen, H.; Li, Y.H. Physiological response and evaluation of different peach varieties under low temperature stress. J. China Agric. Uni. 2022, 27, 66–77. [Google Scholar]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of Reactive Oxygen Species and Hormones in Plant Responses to Temperature Changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef]
- Xu, G.X.; He, M.Q.; Zhao, D.Y.; Lyu, D.G.; Qin, S.J. Physiological and Structural Changes in Apple Tree Branches of Different Varieties during Dormancy. Horticulturae 2023, 9, 947. [Google Scholar] [CrossRef]
- Dey, S.; Biswas, A.; Huang, S.Q.; Li, D.F.; Liu, L.L.; Deng, Y.; Xiao, A.P.; Birhanie, Z.M.; Zhang, J.J.; Li, J.J.; et al. Low Temperature Effect on Different Varieties of Corchorus capsularis and Corchorus olitorius at Seedling Stage. Agronomy 2021, 11, 2547. [Google Scholar] [CrossRef]
- Tang, H.X.; Yang, X.M.; Feng, L.J.; Zhu, F.; Zhou, J.L.; Yin, Y.L. Analysis of Freezing Tolerances and Physiological Differences of Three Pomegranate Cultivars During the Overwintering. Acta Hortic. Sin. 2023, 50, 1563–1573. [Google Scholar]
- Li, M.H.; Jiang, Y.; Wang, A.; Li, X.B.; Zhu, W.; Yan, C.F.; Du, Z.; Shi, Z.; Lei, J.P.; Schönbeck, L.; et al. Active Summer Carbon Storage for Winter Persistence in Trees at the Cold Alpine Treeline. Tree Physiol. 2018, 38, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.Q.; Yang, L.T.; Li, Y.R. Physiological and Biochemical Characteristics Related to Cold Resistance in Sugarcane. Sugar Technol. 2015, 17, 49–58. [Google Scholar] [CrossRef]
- Li, Q.Y.; Lu, B.; Zhao, J.W.; Li, H.; Li, Y.; Miao, S.Y.; Lu, B.S. Physiological response and cold resistance evaluation of different Pyrus calleryan varieties under low temperature stress. J. Northwest A F Univ. 2020, 48, 86–94, 110. [Google Scholar]
- Liu, Z.Y.; Liu, J.L.; Zhu, Y.Y.; Yang, Y.S.; Chen, L.Y.; Zhang, X.M.; Qi, G.H. Research Progress on the Response Mechanism of Woody Plants to low Temperature. J. Northwest For. Univ. 2022, 37, 157–163. [Google Scholar]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Liu, B.B.; Chen, L.N.; Niu, J.; Li, H.X.; Zhang, J.; Cao, S.Y. Selection of methods for evaluation on cold tolerance of six pomegranate varieties. J. Fruit. Sci. 2018, 35, 66–73. [Google Scholar]
- Wang, H.N. Studies on Cold Resistance and Its Related Factor of Young Apple Trees on the SH40 Dwarfing Interstocks. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2014. [Google Scholar]
- Jing, J.L.; Liu, M.X.; Wei, X.; Xu, J.Z.; Li, Z.Y.; Zhang, X.Y.; Zhou, S.S. Evaluation of cold hardiness of several apple interstocks. J. Fruit. Sci. 2022, 39, 970–981. [Google Scholar]
- Zhang, X.X.; Lu, J.G.; Zhu, Y.L. Study on the shoot tissue structure of Chimonanthus varieties in relation to cold resistance. Forest. Ecol. Sci. 2012, 27, 432–434. [Google Scholar]
- Amrina, S.; Vivek, D.; Tejpal, G.; Singh, A.P.; Yelam, S.; Pandey, G.K. Simultaneous over-expression of PaSOD and RaAPX in transgenic Arabidopsis thaliana confers cold stress tolerance through increase in vascular lignifications. PLoS ONE 2014, 9, e110302. [Google Scholar]
- García-Cervigón, A.I.; Fajardo, A.; Caetano-Sánchez, C.; Camarero, J.J.; Olano, J.M. Xylem anatomy needs to change, so that conductivity can stay the same: Xylem adjustments across elevation and latitude in Nothofagus pumilio. Ann. Bot. 2020, 125, 1101–1112. [Google Scholar] [CrossRef]
- Shalaev, E.Y.; Steponkus, P.L. Phase behavior and glass transition of 1, 2-dioleoylphosphatidylethanolamine (DOPE) dehydrated in the presence of sucrose. Biochim. Biophys. Acta 2001, 1514, 100–116. [Google Scholar] [CrossRef]
- Hongbo, S.; Zongsuo, L.; Mingan, S. Osmotic regulation of 10 wheat (Triticum aestivum L.) genotypes at soil water deficits. Colloids Surf. B Biointerfaces 2006, 47, 132–139. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Zhang, L.S.; Chang, R.F.; Liu, G.J.; Han, J.C.; Chen, H. Study on the relationship between tissue structure and cold resistance of peach branches. J. Hebei Agric. Sci. 2014, 18, 29–33. [Google Scholar]
- Liu, R.L.; Miao, Y.; Zhang, Y.C.; Chen, J.H.; Dong, S.J. Cold resistance of introduced Prunus armeniaca var. ansu clones and families. Non-Wood For. Res. 2023, 41, 137–149. [Google Scholar]
- He, Q.F.; Cao, Y.Y.; Peng, Y.H.; Huang, Z.L.; Hao, H.K.; Tan, C.Q. Comprehensive evaluation of cold resistance of individual plants of different Acacia species. Chin. J. Ecol. 2019, 38, 1339–1345. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clones | Provenances | Average Seeding Height /cm | Average Diameter /mm | Longitude /E | Latitude /N | Mean Altitude (m) | Annual Average Temperature/°C |
|---|---|---|---|---|---|---|---|
| 89, 90, 91 | Kazuo, Liaoning (LK) | 74.71 | 9.79 | 119°50′ | 41°24′ | 407.30 | 8.70 |
| 530, 531, 535 | Xiaxian, Shanxi (SX) | 91.16 | 11.42 | 111°12′ | 35°50′ | 539.67 | 12.90 |
| 604, 612, 614 | Jiaxian, Shaanxi (SJ) | 89.91 | 11.38 | 110°29′ | 38°40′ | 507.17 | 10.00 |
| 637, 643, 645 | Yanchuan, Shaanxi (SY) | 83.85 | 9.14 | 110°19′ | 36°52′ | 561.00 | 10.80 |
| 702, 706, 709 | Fuxing, Heibei (HF) | 90.82 | 11.02 | 114°18′ | 36°34′ | 180.62 | 13.50 |
| 750, 752, 753 | Neiqiu, Heibe (HN) | 71.44 | 10.29 | 114°22′ | 37°14′ | 168.13 | 11.50 |
| 853, 859, 862 | Changqing, Shandong (SC) | 85.07 | 9.99 | 116°52′ | 36°29′ | 112.36 | 13.80 |
| Clones | Logistic Equation | R2 | LT50/°C |
|---|---|---|---|
| 89 | y = 100/(1 + 7.6845e−0.0454t) | 0.71 | −44.04 |
| 90 | y = 100/(1 + 4.4696e−0.0380t) | 0.76 | −39.25 |
| 91 | y = 100/(1 + 4.1272e−0.0327t) | 0.75 | −40.39 |
| LK | y = 100/(1 + 5.1074e−0.0392t) | 0.74 | −41.60 |
| 530 | y = 100/(1 + 2.4562e−0.0475t) | 0.73 | −18.92 |
| 531 | y = 100/(1 + 2.7210e−0.0355t) | 0.86 | −24.97 |
| 535 | y = 100/(1 + 2.9868e−0.0356t) | 0.65 | −28.42 |
| SX | y = 100/(1 + 2.7012e−0.0417t) | 0.76 | −23.83 |
| 604 | y = 100/(1 + 4.1148e−0.0321t) | 0.75 | −39.74 |
| 612 | y = 100/(1 + 4.3929e−0.0319t) | 0.76 | −41.33 |
| 614 | y = 100/(1 + 5.3228e−0.0431t) | 0.74 | −39.71 |
| SJ | y = 100/(1 + 4.5814e−0.0375t) | 0.75 | −40.59 |
| 637 | y = 100/(1 + 2.7584e−0.030t) | 0.82 | −32.03 |
| 643 | y = 100/(1 + 2.6461e−0.0317t) | 0.86 | −29.26 |
| 645 | y = 100/(1 + 4.0342e−0.0509t) | 0.68 | −26.57 |
| SY | y = 100/(1 + 3.0716e−0.0388t) | 0.78 | −28.92 |
| 702 | y = 100/(1 + 3.2482e−0.0554t) | 0.83 | −22.27 |
| 706 | y = 100/(1 + 2.4900e−0.0299t) | 0.79 | −29.91 |
| 709 | y = 100/(1 + 4.1103e−0.0361t) | 0.73 | −35.60 |
| HF | y = 100/(1 + 3.1782e−0.0406t) | 0.82 | −28.48 |
| 750 | y = 100/(1 + 4.3649e−0.0477t) | 0.77 | −30.89 |
| 752 | y = 100/(1 + 2.7007e−0.0395t) | 0.89 | −24.71 |
| 753 | y = 100/(1 + 6.0515e−0.0438t) | 0.71 | −37.20 |
| HN | y = 100/(1 + 3.9932e−0.0435t) | 0.81 | −31.83 |
| 853 | y = 100/(1 + 2.7569e−0.0367t) | 0.87 | −27.41 |
| 859 | y = 100/(1 + 2.8537e−0.0307t) | 0.80 | −29.37 |
| 862 | y = 100/(1 + 2.8637e−0.0280t) | 0.82 | −27.05 |
| SC | y = 100/(1 + 2.8474e−0.0373t) | 0.83 | −28.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Zhang, Y.; Kang, Y.; Wang, Y.; Liu, R.; Liu, Q.; Dong, S. Physiological Response to Low-Temperature Stress and Cold Resistance Evaluation of Ziziphus jujuba var. spinosa Clones from Different Provenances. Forests 2024, 15, 1130. https://doi.org/10.3390/f15071130
Li B, Zhang Y, Kang Y, Wang Y, Liu R, Liu Q, Dong S. Physiological Response to Low-Temperature Stress and Cold Resistance Evaluation of Ziziphus jujuba var. spinosa Clones from Different Provenances. Forests. 2024; 15(7):1130. https://doi.org/10.3390/f15071130
Chicago/Turabian StyleLi, Biao, Yuncheng Zhang, Ying Kang, Yijin Wang, Ruilin Liu, Qingbai Liu, and Shengjun Dong. 2024. "Physiological Response to Low-Temperature Stress and Cold Resistance Evaluation of Ziziphus jujuba var. spinosa Clones from Different Provenances" Forests 15, no. 7: 1130. https://doi.org/10.3390/f15071130
APA StyleLi, B., Zhang, Y., Kang, Y., Wang, Y., Liu, R., Liu, Q., & Dong, S. (2024). Physiological Response to Low-Temperature Stress and Cold Resistance Evaluation of Ziziphus jujuba var. spinosa Clones from Different Provenances. Forests, 15(7), 1130. https://doi.org/10.3390/f15071130