
The Application of Percolation Theory in Modeling the Vertical Distribution of Soil Organic Carbon in the Changbai Mountains

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Field Sampling
2.2. Data Analysis
2.2.1. Vertical Distribution of SOC in CMMF
2.2.2. Prediction of SOC Profile in CMMF
3. Results
3.1. Vertical Distribution of SOC in Three Forests in CMMF
3.2. Effect of Forest Type on Vertical Distribution of SOC
3.3. Prediction of Vertical Distribution of SOC in CMMF
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Nachtergaele, F.O.; Velthuizen, H.V.; Verelst, L.; Batjes, N.H.; Wiberg, D. The Harmonized World Soil Database; Institut für Bodenforschung, Universität für Bodenkultur: Vienna, Austria, 2010. [Google Scholar]
- Hengl, T.; De Jesus, J.M.; MacMillan, R.A.; Batjes, N.H.; Heuvelink, G.B.M.; Ribeiro, E.; Samuel-Rosa, A.; Kempen, B.; Leenaars, J.G.B.; Walsh, M.G.; et al. SoilGrids1km—Global Soil Information Based on Automated Mapping. PLoS ONE 2014, 9, e105992. [Google Scholar] [CrossRef]
- Akbari, M.; Goudarzi, I.; Tahmoures, M.; Elveny, M.; Bakhshayeshi, I. Predicting Soil Organic Carbon by Integrating Landsat 8 OLI, GIS and Data Mining Techniques in Semi-Arid Region. Earth Sci. Inform. 2021, 14, 2113–2122. [Google Scholar] [CrossRef]
- Arroyo, I.; Tamaríz-Flores, V.; Castelán, R. Mapping Forest Cover and Estimating Soil Organic Matter by GIS-Data and an Empirical Model at the Subnational Level in Mexico. Forests 2023, 14, 539. [Google Scholar] [CrossRef]
- Hobley, E.; Wilson, B.; Wilkie, A.; Gray, J.; Koen, T. Drivers of Soil Organic Carbon Storage and Vertical Distribution in Eastern Australia. Plant Soil 2015, 390, 111–127. [Google Scholar] [CrossRef]
- Ota, M.; Nagai, H.; Koarashi, J. Root and Dissolved Organic Carbon Controls on Subsurface Soil Carbon Dynamics: A Model Approach: CONTROLS ON SUBSURFACE CARBON DYNAMICS. J. Geophys. Res. Biogeosci. 2013, 118, 1646–1659. [Google Scholar] [CrossRef]
- Fröberg, M.; Jardine, P.M.; Hanson, P.J.; Swanston, C.W.; Todd, D.E.; Tarver, J.R.; Garten, C.T. Low Dissolved Organic Carbon Input from Fresh Litter to Deep Mineral Soils. Soil Sci. Soc. Am. J. 2007, 71, 347–354. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Deep Soil Organic Matter—A Key but Poorly Understood Component of Terrestrial C Cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Fröberg, M.; Hanson, P.J.; Trumbore, S.E.; Swanston, C.W.; Todd, D.E. Flux of Carbon from 14C-Enriched Leaf Litter throughout a Forest Soil Mesocosm. Geoderma 2009, 149, 181–188. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Kramer, C.; Trumbore, S.; Fröberg, M.; Cisneros Dozal, L.M.; Zhang, D.; Xu, X.; Santos, G.M.; Hanson, P.J. Recent (<4 Year Old) Leaf Litter Is Not a Major Source of Microbial Carbon in a Temperate Forest Mineral Soil. Soil Biol. Biochem. 2010, 42, 1028–1037. [Google Scholar] [CrossRef]
- Tate, K.R.; Lambie, S.M.; Ross, D.J.; Dando, J. Carbon Transfer from 14C-Labelled Needles to Mineral Soil, and 14C-CO2 Production, in a Young Pinus Radiata Don Stand. Eur. J. Soil Sci. 2011, 62, 127–133. [Google Scholar] [CrossRef]
- Leifeld, J.; KoÈgel-Knabner, I. Organic Carbon and Nitrogen in fine Soil Fractions after Treatment with Hydrogen Peroxide. Soil Biology 2001, 33, 2155–2158. [Google Scholar] [CrossRef]
- Neff, J.C.; Asner, G.P. Dissolved Organic Carbon in Terrestrial Ecosystems: Synthesis and a Model. Ecosystems 2001, 4, 29–48. [Google Scholar] [CrossRef]
- Baisden, W.T.; Parfitt, R.L. Bomb 14C Enrichment Indicates Decadal C Pool in Deep Soil? Biogeochemistry 2007, 85, 59–68. [Google Scholar] [CrossRef]
- Elzein, A.; Balesdent, J. Mechanistic Simulation of Vertical Distribution of Carbon Concentrations and Residence Times in Soils. Soil Sci. Soc. Am. J. 1995, 59, 1328–1335. [Google Scholar] [CrossRef]
- Hiederer, R. Distribution of Organic Carbon in Soil Profile Data; EUR 23980 EN; Office for Official Publications of the European Communities: Luxembourg, 2009; 126p. [Google Scholar]
- Ottoy, S.; Elsen, A.; Van De Vreken, P.; Gobin, A.; Merckx, R.; Hermy, M.; Van Orshoven, J. An Exponential Change Decline Function to Estimate Soil Organic Carbon Stocks and Their Changes from Topsoil Measurements. Eur. J. Soil Sci. 2016, 67, 816–826. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, Q. Organic Carbon Content and Distribution in Soils under Different Land Uses in Tropical and Subtropical China. Plant Soil 2001, 231, 175–185. [Google Scholar]
- Yang, Y.H.; Fang, J.Y.; Guo, D.L.; Ji, C.J.; Ma, W.H. Vertical Patterns of Soil Carbon, Nitrogen and Carbon: Nitrogen Stoichiometry in Tibetan Grasslands. Biogeosci. Discuss. 2010, 7, 1–24. [Google Scholar]
- Robinson, N.; Benke, K. Analysis of Uncertainty in the Depth Profile of Soil Organic Carbon. Environments 2023, 10, 29. [Google Scholar] [CrossRef]
- Yu, F.; Liu, Q.; Fan, C.; Li, S. Modeling the Vertical Distribution of Soil Organic Carbon in Temperate Forest Soils on the Basis of Solute Transport. Front. For. Glob. Change 2023, 6, 1228145. [Google Scholar] [CrossRef]
- Hobley, E.U.; Wilson, B. The Depth Distribution of Organic Carbon in the Soils of Eastern Australia. Ecosphere 2016, 7, e01214. [Google Scholar] [CrossRef]
- Hunt, A.G.; Skinner, T.E. Longitudinal Dispersion of Solutes in Porous Media Solely by Advection. Philos. Mag. 2008, 88, 2921–2944. [Google Scholar] [CrossRef]
- Sheppard, A.P.; Knackstedt, M.A.; Pinczewski, W.V.; Sahimi, M. Invasion Percolation: New Algorithms and Universality Classes. J. Phys. A Math. Gen. 1999, 32, L521–L529. [Google Scholar] [CrossRef]
- Zhou, Y.; Su, J.; Janssens, I.A.; Zhou, G.; Xiao, C. Fine Root and Litterfall Dynamics of Three Korean Pine (Pinus Koraiensis) Forests along an Altitudinal Gradient. Plant Soil 2014, 374, 19–32. [Google Scholar] [CrossRef]
- Chen, X.; Li, B.-L. Change in Soil Carbon and Nutrient Storage after Human Disturbance of a Primary Korean Pine Forest in Northeast China. For. Ecol. Manag. 2003, 186, 197–206. [Google Scholar] [CrossRef]
- Liu, C. Accumulation of Soil Organic Matter and Influential Factors on the North Slope of Changbai Mountain. Master’s Thesis, Northeast Forestry University, Harbin, China, 2004. [Google Scholar]
- Yang, X.; Cheng, J.; Meng, L.; Han, J. Features of Soil Organic Carbon Storage and Vertical Distribution in different Forests. Chin. Agric. Sci. Bull. 2010, 26, 132–135. [Google Scholar]
- Liu, L.; Wang, H.; Dai, W.; Yang, X.; Li, X. Characteristics of Soil Organic Carbon and Nutrients under Four Forest Stands in Eastern Part of Changbai Mountains. Bull. Soil Water Conserv. 2013, 33, 79–85. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.; Dai, W.; Yang, X.; Li, X. Spatial heterogeneity of soil organic carbon and nutrients in low mountain area of Changbai Mountains. Chin. J. Appl. Ecol. 2014, 25, 2460–2468. [Google Scholar] [CrossRef]
- Wei, Y.; Yu, D.; Wang, Q.; Zhou, L.; Zhou, W.; Fang, X.; Gu, X.; Dai, L. Soil organic density and its influencing factors of major forest types in the forest region of northeast China. Chin. J. Appl. Ecol. 2013, 24, 3333–3340. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, H.; Sun, J.; Cui, X. Soil Organic Carbon Density as Affected by Topography and Physical Protection Factors in the Secondary Forest Area of Zhangguangcai Mountains, Northeast China. J. Beijing For. Univ. 2016, 38, 94–106. [Google Scholar]
- Song, Y.; Zhang, Y.; Zhao, Z.; Guan, Q.; Li, Y.; Xu, L. Distribution Characteristics of Soil Organic Carbon and Total Nitrogen of Different Forest Types in the West of Changbai Mountain. J. Northwest For. Univ. 2018, 33, 39–44. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, Y.; Guan, Q.; Xu, L.; Li, Y.; Sui, Z.; Zhao, H. Soil Organic Carbon Content and Its Relations with Soil Physicochemical Properties of Spruce-Fir Mixed Stands in Changbai Mountains. J. Northeast For. Univ. 2019, 47, 70–74. [Google Scholar]
- Sun, Y. Co-Accumulation Characters of Carbon and Nitrogen in Soils under Two Forest Types in Changbai Mountain. Master’s Thesis, Northeast Forestry University, Harbin, China, 2018. [Google Scholar]
- Li, S.; Zhao, H.; Gao, F.; Gao, L.; Wang, M.; Cui, X. Soil organic carbon pools and profile distribution under broad-leaved Korean Pine forest and Betula platyphylla secondary forest in Changbai Mountain. J. Temp. For. Res. 2019, 2, 7–10. [Google Scholar] [CrossRef]
- Zhao, H. Changes of Forest Soil Organic Carbon and Nitrogen Pools as Driven by Secondary Succession in Changbai Mountain. Master’s Thesis, Northeast Forestry University, Harbin, China, 2019. [Google Scholar]
- Yu, D.; Hu, F.; Zhang, K.; Liu, L.; Li, D. Available Water Capacity and Organic Carbon Storage Profiles in Soils Developed from Dark Brown Soil to Boggy Soil in Changbai Mountains, China. Soil Water Res. 2020, 16, 11–21. [Google Scholar] [CrossRef]
- Trumbore, S.; Da Costa, E.S.; Nepstad, D.C.; Barbosa De Camargo, P.; Martinelli, L.A.; Ray, D.; Restom, T.; Silver, W. Dynamics of Fine Root Carbon in Amazonian Tropical Ecosystems and the Contribution of Roots to Soil Respiration: AMAZON FINE ROOT CARBON DYNAMICS. Glob. Change Biol. 2006, 12, 217–229. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A Global Analysis of Root Distributions for Terrestrial Biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.-D. A Global Budget for Fine Root Biomass, Surface Area, and Nutrient Contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef]

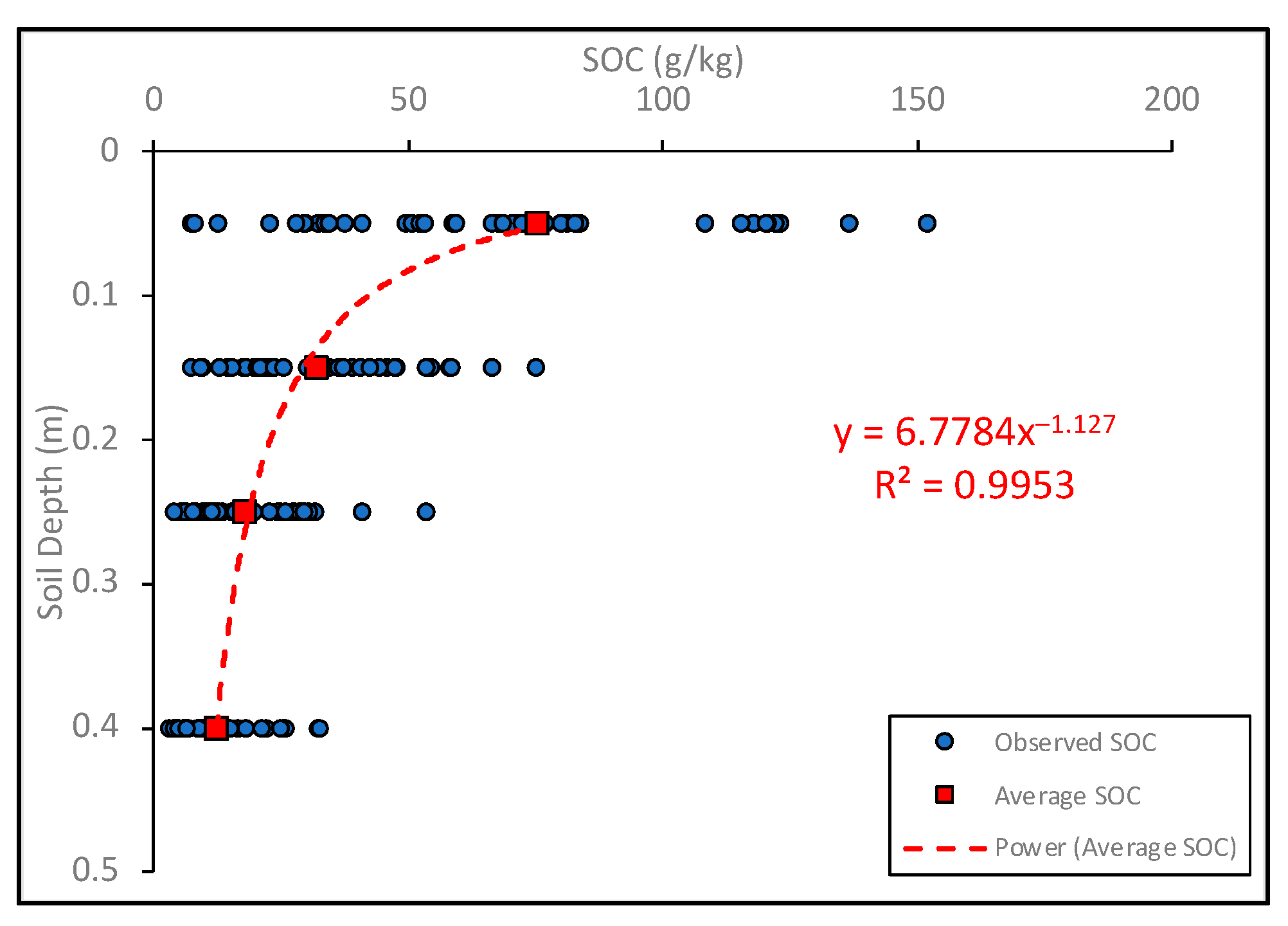
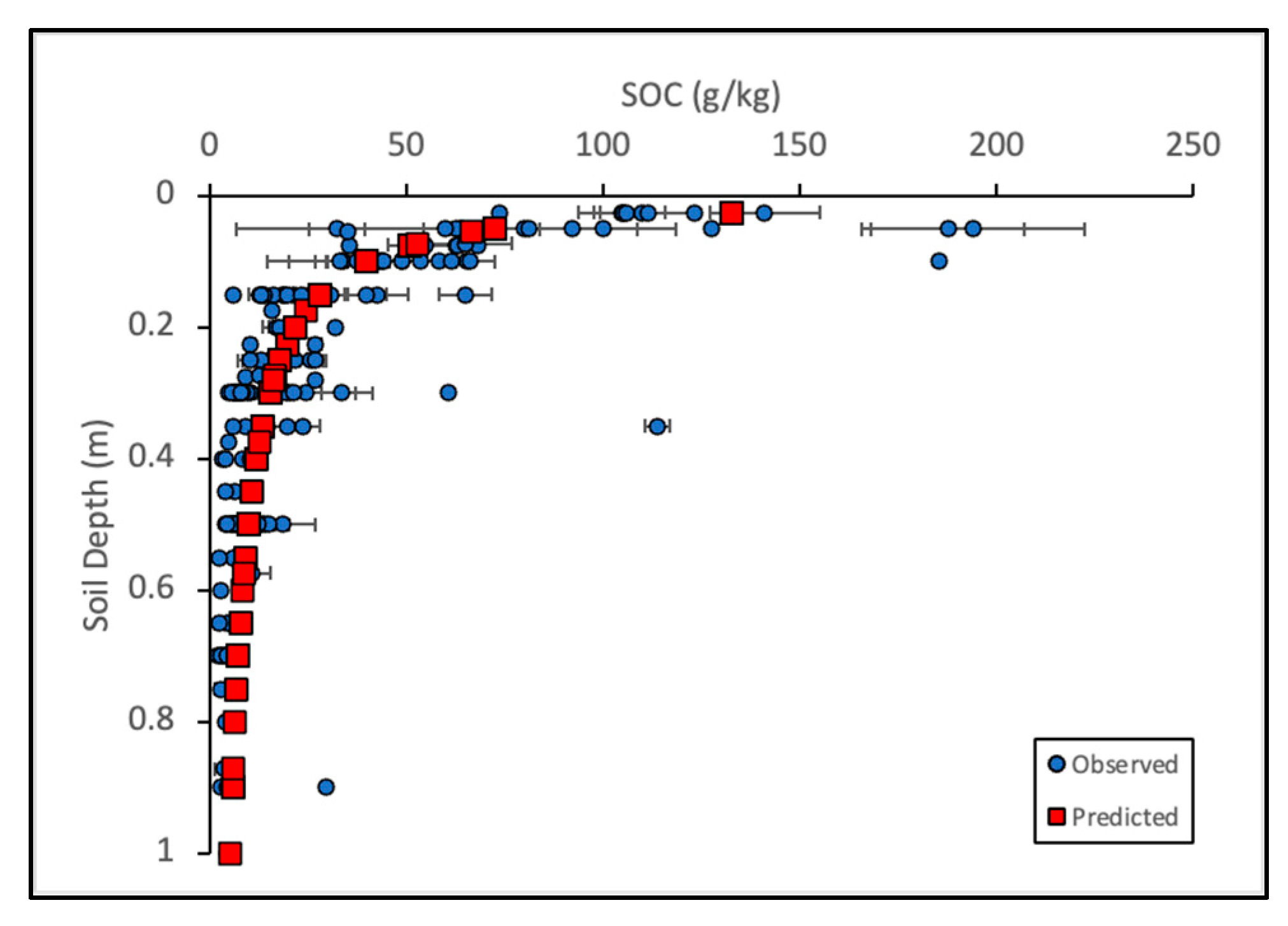


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BF | MBF | BKF | |
|---|---|---|---|
| Location | 42.44 N | 42.40 N | 42.52 N |
| 126.93 E | 126.70 E | 127.01 E | |
| Altitude (m) | 589.6 | 553.0 | 616.4 |
| Slope (°) | <3 | <3 | 4 |
| Mean height (m) | 19.01 | 17.92 | 15.80 |
| Density (stems/ha) | 833 | 967 | 683 |
| Location | Forest | No. of Profiles | |
|---|---|---|---|
| N | E | ||
| 43.63 | 126.03 | Mixed broadleaved | 3 |
| 43.46 | 126.81 | Mixed broadleaved | 3 |
| 44.37 | 126.91 | Mixed broadleaved | 3 |
| 43.66 | 129.97 | Coniferous | 3 |
| 43.48 | 129.35 | Mixed conifer–broadleaved | 3 |
| 43.05 | 129.05 | Mixed broadleaved | 3 |
| 43.33 | 130.13 | Mixed conifer–broadleaved | 3 |
| 42.13 | 128.21 | Mixed conifer–broadleaved | 1 |
| 43.39 | 130.16 | Mixed conifer–broadleaved | 3 |
| 42.40 | 128.46 | Mixed conifer–broadleaved | 3 |
| 43.32 | 130.39 | Mixed broadleaved | 3 |
| 42.49 | 127.77 | Mixed conifer–broadleaved | 3 |
| 43.23 | 130.61 | Mixed conifer–broadleaved | 2 |
| 44.79 | 123.02 | Mixed broadleaved | 2 |
| 42.47 | 127.77 | Mixed conifer–broadleaved | 3 |
| 42.01 | 127.39 | Mixed conifer–broadleaved | 2 |
| Location | Forest | No. of Profiles | Source | |
|---|---|---|---|---|
| N | E | |||
| 41.72–42.43 | 127.70–128.28 | Coniferous, broadleaved Korean pine mixed | 9 | [27] |
| 42.4 | 128.08 | Broadleaved Korean pine mixed, mixed broadleaved | 10 | [28] |
| North Slope of Changbai Mountains | Forest type unavailable from the source | 18 | [29] | |
| 42.06–42.42 | 128.07–128.11 | Erman’s birch, coniferous | 6 | [30] |
| 43.30–43.40 | 130.38–130.62 | Larch, mixed conifer–broadleaved, mixed broadleaved | 16 | [31] |
| 43.30–43.40 | 130.38–130.62 | Larch, mixed conifer–broadleaved, mixed broadleaved | 63 | [32] |
| 43.42–42.65 | 127.75–128.00 | Coniferous, mixed conifer–broadleaved, mixed broadleaved | 69–86 | [33] |
| 44.23–45.60 | 127.90–129.05 | Broadleaved Korean pine mixed, mixed broadleaved | 36 | [34] |
| 43.28–43.42 | 130.08–130.33 | Spruce–fir, mixed conifer–broadleaved | 27 | [35] |
| 43.17–43.85 | 127.33–128.01 | Mixed conifer–broadleaved, mixed broadleaved, larch | 33 | [36] |
| 42.36–42.39 | 127.48–128.00 | Broadleaved Korean pine mixed, poplar–birch | 6 | [37] |
| 42.31–42.50 | 127.83–128.13 | Broadleaved Korean pine mixed, poplar–birch | 20 | [38] |
| 42.32–42.50 | 127.83–128.13 | Broadleaved Korean pine mixed, poplar–birch | 50 | [39] |
| Changbai Mountains | Forest type unavailable from the source | 6 | [40] | |
| Soil Depth (m) | SOC (g/kg) | p-Value | ||
|---|---|---|---|---|
| BF | MBF | BKF | ||
| 0.05 | 89.85 ± 6.75 | 95.26 ± 5.35 | 80.69 ± 9.34 | 0.1237 |
| 0.15 | 42.81 ± 6.15 | 40.22 ± 6.44 | 30.10 ± 4.4 | 0.0750 |
| 0.25 | 22.15 ± 6.42 | 22.63 ± 4.13 | 18.90 ± 2.87 | 0.5991 |
| 0.35 | 11.63 ± 3.17 | 16.68 ± 3.99 | 14.40 ± 1.52 | 0.2108 |
| 0.45 | 4.58 ± 1.58 | 13.35 ± 2.18 | 10.12 ± 1.88 | 0.0037 |
| 0.55 | 2.18 ± 0.75 | 11.09 ± 2.49 | 9.41 ± 2.56 | 0.0045 |
| 0.65 | 2.23 ± 0.4 | 8.87 ± 0.70 | 7.56 ± 0.92 | 0.0001 |
| Forest | Power-Law Exponent a | Fine Root Biomass above 30 cm (%) b | Proportion of Biomass Aboveground (%) c | β d |
|---|---|---|---|---|
| Boreal | −0.905 | 83 | 76 | 0.943 |
| Temperate deciduous | −1.103 | 63 | 85 | 0.967 |
| Temperate evergreen | −1.126 | unavailable | 81 | NA |
| Tropical deciduous | −2.127 | 42 | 75 | 0.982 |
| Tropical evergreen | −1.39 | 57 | 84 | 0.972 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Fan, C. The Application of Percolation Theory in Modeling the Vertical Distribution of Soil Organic Carbon in the Changbai Mountains. Forests 2024, 15, 1155. https://doi.org/10.3390/f15071155
Yu F, Fan C. The Application of Percolation Theory in Modeling the Vertical Distribution of Soil Organic Carbon in the Changbai Mountains. Forests. 2024; 15(7):1155. https://doi.org/10.3390/f15071155
Chicago/Turabian StyleYu, Fang, and Chunnan Fan. 2024. "The Application of Percolation Theory in Modeling the Vertical Distribution of Soil Organic Carbon in the Changbai Mountains" Forests 15, no. 7: 1155. https://doi.org/10.3390/f15071155
APA StyleYu, F., & Fan, C. (2024). The Application of Percolation Theory in Modeling the Vertical Distribution of Soil Organic Carbon in the Changbai Mountains. Forests, 15(7), 1155. https://doi.org/10.3390/f15071155