Abstract
Fusarium circinatum, the causative agent of pine pitch canker disease, is a pathogenic fungus that poses a significant threat to pine forests globally. It infects various Pinus species, causing resinous cankers, needle discoloration, and tree death. The disease severely impacts forest ecosystems, necessitating cost-effective and environmentally friendly management strategies. Contaminated pine seeds and seedlings are the main pathways for introducing this fungus to disease-free areas. To mitigate this disease and prevent its spread, it is crucial to implement new processes in forest plant production systems that align with the existing conditions of forest nurseries, ensuring effective and sustainable management. With this in mind, a national collaborative study involving 14 Portuguese partners was initiated to develop new prevention and mitigation strategies. In this work, four different treatments—MennoFlorades, Captan, ethanol, and hot water—were tested for their ability to eliminate F. circinatum from contaminated Pinus seeds in vitro. The most effective treatments were selected for further in vitro assays and real-context nursery germination trials to assess their impacts on seed germination, plant production, and certification. MennoFlorades, Captan, and hot water were tested in the nursery, with hot water showing the most promising results due to its negligible impact on seedlings, eco-friendly nature, ease of implementation, and cost-effectiveness. These findings offer promising prospects for preventing pine pitch canker outbreaks in nurseries and, consequently, in forests.
1. Introduction
Fusarium circinatum Nirenberg & O’Donnell, the causal agent of pitch canker disease, is recognized as one of the most severe diseases affecting pine trees, posing a significant threat to forest nurseries and plantations worldwide [1,2,3]. This pathogenic fungus targets Pinus species and Pseudotsuga menziesii (Douglas fir), displaying varying degrees of virulence, depending on the host species and prevailing biotic and abiotic conditions [3,4,5,6,7]. The pathogen’s impact is pervasive, affecting all stages of pine growth. Common symptoms in seedlings include blight and damping-off, while on young and mature trees, dieback of branches and stems is frequent, often accompanied by conspicuous cankers and resin exudation. Additionally, dieback symptoms in the crown can also occur due to water flow obstruction caused by cankers and the saturation of xylem by excessive resin, potentially strangling the treetop and leading to tree death [2,3,8]. This pathogen was first recorded in the US in 1946, and since then, sporadic outbreaks and epidemics have been reported in numerous countries [4,9,10]. In Europe, occurrences have been reported in Portugal, Spain, France, and Italy among various susceptible Pinus species [11,12,13,14,15].
In Portugal, the coniferous forest holds undeniable social, economic, and environmental importance, with native species like maritime pine (Pinus pinaster) and stone pine (Pinus pinea) dominating the territorial landscape. The maritime pine stands as the third most significant forest species in the country, covering 22.1% of the mainland area. However, it has experienced a declining trend, primarily attributed to rural wildfires, surpassing the expansion of new plantations and further compounded by forest pests like pine pitch canker and pine wood nematode (Bursaphelenchus xylophilus), impacting productivity and investment confidence in this species. In recent years, this decrease in area seems to have been slowing down, showcasing the resilience of these ecosystems to disturbances [16]. The maritime pine industry, a major carbon reservoir in the Portuguese forest (90.3 Gg CO2), accounts for 80% of employment and 88% of industrial companies in the forestry sector, contributing to 52% of the Gross Value Added (GVA) and 3.2% of national exports of goods [16,17]. Besides the maritime pine, the stone pine also holds a significant representation in the national forest, ranking as the fifth forest species in Portugal and covering 6% of the mainland area, which corresponds to over 20% of its global distribution [16]. While other species are also naturally present, they have less prominence. Noteworthy about the stone pine is its potential for use in multifunctional agroforestry systems, ensuring a variety of products and the provision of ecosystem services related to soil and water protection, carbon sequestration, and biodiversity maintenance. Similarly to fruit species, it offers the possibility of orchard cultivation in grafted plantations dedicated to pine nut production. With an estimated pine nut production ranging between 70,000 and 120,000 tons annually, equivalent to a value between EUR 75 million and EUR 128.5 million, exports of this product reach approximately EUR 15 million per year [16,17]. The sustainability of these vital economic sectors relies on forest resources and their effective management, including afforestation and reforestation actions, which require access to high-quality plants free from phytosanitary issues.
Efforts to manage the spread of tree pathogens, including F. circinatum, face significant challenges due to the lack of economically viable treatments for large-scale control. Strategies primarily revolve around preventing the pathogen’s introduction, early detection, and eradication of outbreaks in previously disease-free areas. Since 2007, F. circinatum has been categorized as a quarantine fungus within the European Union (EU), which has imposed strict measures to prevent its transmission through infected materials and to curb disease expansion [18,19]. Consequently, a zero-tolerance policy is enforced. According to the pest risk assessment conducted by the European Food and Environment Safety Agency (EFSA), contaminated pine seeds and seedlings represent the primary pathways for potential fungus spread to disease-free areas [20]. Implementing efficient management measures is crucial to minimize the likelihood of pathogen introduction and reduce the associated costs of eradication and control efforts.
In Portugal, since its initial detection in 2008 in nursery plants [15], F. circinatum has been identified in 27 locations across the country, including both nurseries and pine forests, with a higher incidence in the northern and central regions. This has resulted in the destruction of 1.8 million plants and the quarantine of 2500 kg of seeds from host species. The presence of this fungus continues to pose both ecological and economic threats, emphasizing the urgent need to develop mechanisms to prevent or minimize its risk of dispersal. Additionally, as previously mentioned, while current efforts to control the spread of this disease focus heavily on the early reaction to the presence of the fungus to mitigate outbreaks, they should instead be redirected towards preventing infection altogether. In this context, the operational group GO +PrevCRP was established as part of the Rural Development Program PDR2020, bringing together 14 national partners. The action plan, spanning 4 years (2017–2021), aimed to develop new, effective, and environmentally friendly prevention and mitigation strategies across various crucial factors contributing to the spread of the disease within nurseries, such as contaminated irrigation water and substrates, the results of which were recently published [21,22,23,24]. These preventive measures must be grounded in experimental findings, ensuring their feasibility and seamless integration into nursery operations without compromising seed germination rates or the development and quality of the resulting plants. Given that the primary means of pitch canker disease dispersal are contaminated pine seeds and seedlings, the main goals of this study were (i) to assess the effectiveness of certain treatments in eliminating F. circinatum inoculum from seeds; (ii) to evaluate their impacts on seed germination; and (iii) to apply these experimental findings in practical field settings. This included conducting large-scale experiments in nurseries to gather a wider sample size and assess the feasibility of implementing treatment measures in real-world scenarios. For this purpose, several treatments were selected based on their previously reported fungicide activity. To the best of our knowledge, the efficacy of some of these treatments as seed disinfectants against F. circinatum is being tested for the first time, particularly on Pinus species commonly found in Portuguese production and conservation forests.
2. Materials and Methods
2.1. Artificial Seed Inoculation
Seeds from different Pinus species, specifically Pinus pinaster, P. pinea and P. radiata, potential hosts of Fusarium circinatum, were provided by the National Centre for Forest Seeds (CENASEF). Pinus pinaster seeds were collected from 80-year-old parental trees, P. radiata seeds from 27-year-old parental trees, and P. pinea seeds from 53-year-old parental trees. All parental trees were healthy and showed no signs of disease. In a controlled laboratory environment, these seeds underwent artificial inoculation (excluding a portion kept for use as a control in later analyses) and subsequent assessment for successful inoculation following the methodology outlined in the OEPP/EPPO Bulletin (2019) 49 (2), 228–247 [25] for fungal detection in seeds. Given the quarantine status of this fungus in the European Union (EU), this procedure was conducted by the National Institute of Agricultural and Veterinary Research, I.P. (INIAV), a state laboratory designated as the National Reference Laboratory (LNR). Upon confirmation of successful seed inoculation, preliminary seed disinfection treatments were initiated.
2.2. Treatment Selection
The tested treatments included MennoFlorades (Menno Chemie–Vertrieb GMBH, Norderstedt, Germany), Captan 800 WDG (Adama, Lisbon, Portugal), ethanol (Biochem Iberica, Montijo, Portugal) at various concentrations, and hot water treatment at different temperatures (Table 1). This selection was made based on their overall fungicide efficacy, as well as their accessibility and user-friendliness. All products were supplied by Biochem Iberica—Agricultural and Industrial Chemicals, Lda.

Table 1.
Details of the treatments used for disinfecting Pinus seeds.
Action mechanisms of the tested treatments are as follows:
MennoFlorades is a surface disinfectant with proven fungicidal efficacy, including against various Fusarium species. Its active ingredient, benzoic acid (C6H5COOH), acts on glycolysis, specifically on phosphofructokinase, which is inhibited by the acidification of the intracellular content caused by the extracellular accumulation of benzoate. This inhibition triggers a cascade effect inhibiting metabolic pathways and a consequent reduction in the ATP concentration and production [26].
Captan (N-trichloromethylthio]-4-cyclohexene-1,2-dicarboximide) is a widely used agricultural fungicide, acting at the mitochondrial level as an uncoupler of oxidative phosphorylation. Its fungicidal capabilities also extend to its affinity for sulfhydryl groups common in mitochondrial enzymes such as NADH dehydrogenase and β–hydroxybutyrate dehydrogenase, leading to their inhibition and an ability to inhibit other crucial processes in respiration [27].
Ethanol, like other alcohol molecules, serves as a base for many fungicides and exhibits antifungal activity on its own. Its primary target is the fungal cell membrane, inducing stress by interacting at the polar–apolar interface, weakening the hydrophobic barrier, and allowing free exchange of polar molecules, thereby disrupting the membrane structure and function. Additionally, it has other effects, such as protein denaturation and inhibition of nutrient uptake [28].
Hot water treatment is a technique already used to combat various phytopathogenic fungi [29] using high temperatures to inhibit fungal development (e.g., through protein denaturation and cell membrane damage, among other mechanisms), and has shown promising efficacy in eliminating Fusarium circinatum from Pinus radiata seeds [30]. According to Liao et al. [31], the lethal temperatures for conidia and mycelium are reported as 52 °C and 55 °C, respectively.
2.3. Treatment Application
The treatments were prepared and applied as specified in Table 1 provided below.
Sterile distilled water (dH2O) was employed to prepare the treatment solutions, and was used for the hot water treatments. The treatments were conducted in sterile glass containers, with seeds being agitated during the application time, following the conditions as indicated in Table 1. In temperature-dependent treatments, the temperature was adjusted and maintained using magnetic stirring hot plates. After the total treatment time, the seeds were collected, allowed to dry at room temperature for 10 min, and then plated onto Dichloran Chloramphenicol Peptone Agar (DCPA) medium plates supplemented with 0.5 mg/mL of streptomycin following the methodology outlined in the OEPP/EPPO Bulletin (2019) 49 (2), 228–247 for F. circinatum detection in seeds [25]. Additionally, for each Pinus species considered in the study, two controls were included: a positive control (C+), where 100 artificially contaminated seeds (see Section 2.1) were left untreated and were plated under the same conditions; and a negative control (C–), where 100 uncontaminated and untreated seeds were plated under the same conditions (Figure 1).

Figure 1.
Stages of Pinus pinaster seed disinfection: (A) treatment with 1.9 g/L Captan; (B) plating on DCPA media following treatment application; (C) observation of Fusarium circinatum growth on the positive control after 5 days of incubation.
The disinfection percentage was calculated by considering zero contaminated seeds (zero F. circinatum isolates) as achieving 100% successful disinfection and applying the following formula when the count was greater than zero: % of disinfection = (no. of contaminated seeds/total plated seeds) × 100. The results of the disinfection treatment were analyzed via the non-parametric Kruskal–Wallis test and Dunn’s post hoc analysis with p < 0.01 using PAST software, version 4.03 [32].
2.4. Fungal Isolation and Identification
For all treatments, the inoculated media plates were aerobically incubated in the dark at 25 °C for 30 days. All emerging colonies were isolated into axenic cultures in duplicate onto Potato Dextrose Agar (PDA, DifcoTM, Sparks, MD, USA) plates suitable for isolation, and Synthetic Nutrient Agar (SNA) plates suitable for identifying F. circinatum based on morphological features. These cultures were aerobically incubated in the dark at 25 °C for a minimum of 10 days. After this incubation period, the number of F. circinatum isolates and the number of isolates of other fungi were documented, and photographic records were also captured. The identification of F. circinatum was carried out according to the protocol following the EPPO guidelines [OEPP/EPPO Bulletin (2019) 49 (2), 228–247, protocol available at https://gd.eppo.int/taxon/GIBBCI/documents (accessed on 23 April 2024)] [25]. Colonies displaying morphologies significantly different from Fusarium spp. were also identified using a combination of molecular and morphological analyses following the standard procedures described below.
Morphological and Molecular Identification of Non-Fusarium Species
Genomic DNA was extracted from PDA pure cultures with the REDextract–N.Amp™ Plant PCR Kit (Sigma Aldrich, St. Louis, MO, USA), with several modifications. A small portion (~1 mm2) of the colonies was scraped from the agar surface into a PCR–style microtube, submerged in 20 μL of extraction solution, and incubated in the thermocycler using the following protocol: 94 °C for 10 min, followed by 60 °C for 13 min and 10 °C for 15 min. After the incubation, 20 μL of dilution solution was added, and the resulting mixture was vortexed for 2 min [33,34]. The obtained DNA was used for the amplification of the ITS–rDNA region by PCR using the fungal universal primer pair ITS1–F and ITS4 [35,36]. PCR reactions, consisting of a final amplification volume of 25 μL, with 12.5 μL of NZYTaq Green Master Mix (NZYTech™, Lisbon, Portugal), 1 μL of each primer (10 mM), 9.5 μL of ultra-pure water, and 1 μL of template DNA, were performed using an ABI GeneAmp™ 9700 PCR System (Applied Biosystems, Waltham, MA, USA) with the following conditions: initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 52 °C for 30 s, and extension at 72 °C for 1 min, with a final extension at 72 °C for 8 min. Visual confirmation of the overall amplification of the ITS region was performed using agarose gel electrophoresis (1.2%) stained with GreenSafe Premium (NZYTech™, Lisbon, Portugal) and visualized in an Molecular Imager Bio–Rad Gel Doc XR™ (Bio–Rad, Hercules, CA, USA). The obtained amplicons were purified with the EXO/SAP Go PCR Purification Kit (GRISP, Porto, Portugal), following the manufacturer’s recommendations, and sequenced using an ABI 3730xl DNA Analyzer system (96 capillary instruments) at STABVIDA, Portugal.
Sequence reads were quality checked and trimmed at both ends using BioEdit Se–quence Alignment Editor© v.7.2.5 (https://bioedit.software.informer.com/download/ (accessed on 7 January 2019)). The newly generated sequences were deposited in the GenBank database, and their accession numbers are included in Supplementary Table S1. Similarity searches were performed, and sequences were queried against the National Center of Biotechnology Information (NCBI) nucleotide database using a BLASTn search algorithm [37]. For genera like Penicillium and Aspergillus, where the ITS region does not provide sufficient differentiation at the species level, isolates were identified only to the genus level. To ensure accurate species identification, molecular results were verified through comprehensive macroscopic and microscopic analysis of taxonomic traits, resorting to Index Fungorum (www.indexfungorum.org (accessed on 20 January 2019)) and Mycobank (https://www.mycobank.org/ (accessed on 20 January 2019)) [38,39].
2.5. Nursery Germination Assays
The best-performing treatments identified in the previous disinfection phase were chosen for nursery germination trials. Collaborative field trials were conducted in partnership with nurseries involved in the project to accommodate larger-scale assessments of the selected treatments. Field trials exclusively employed P. pinaster seeds. Although both P. pinea and P. radiata were included in the preliminary laboratory testing, their seeds were unavailable during the nursery experimental period due to low seed stocks of these species and the necessity to ensure seed availability for nurseries and private stakeholders outside of this project. Nonetheless, as previously noted, P. pinaster is one of the most prevalent species in Portuguese forests, thus representing the highest interest in field trials. Certified seeds intended for commercial distribution (without inoculation) were distributed by CENACEF to the nurseries designated for the trials, and the procedures were carried out in accordance with the specifications outlined in Table 2. After applying treatments to replicate laboratory procedures, the treated seeds were planted in containers filled with substrate, with one seed per container cell. Regular irrigation was provided, and seed germination was monitored at one and two months post-sowing (Figure 2).
Table 2.
Number of containers per treatment, number of cells per container, and total number of cells per treatment used in the nursery germination assays for Pinus pinaster seeds.

Figure 2.
Records of the different stages of the field trials: (A) preparation of containers for sowing; (B) preparation of treatments; and (C) plant certification process.
Forest reproductive materials (FRM) must possess a genetic quality that enables them to originate stable, adapted, resistant, and resilient forest ecosystems capable of addressing both biotic and abiotic challenges that forests face. General standards applicable to the production and commercialization of FRM are defined in accordance with the applicable Portuguese legislation, Decree–Law No. 205/2003, of 12 September, amended and republished by Decree–Law No. 13/2019, of 21 January. To be deemed marketable, a batch of plants must ensure that 95% of its plants exhibit intact and marketable quality and are devoid of any of the following defects: injuries unrelated to pruning or caused by damage during uprooting; absence of buds with potential for producing a main shoot; presence of multiple stems; malformed root systems; indications of desiccation, overheating, mold, rot, or other harmful organisms; or unbalanced growth. Additional criteria, such as the maximum age and size, must also be considered for plant marketing, with specific limits defined for height (ranging from 7 to 45 cm, depending on the species) and for the minimum root collar diameter (ranging from 2 to 3 mm, also species-dependent). To ensure compliance with the established general standards for FRM production and commercialization of plants produced from seeds subjected to the disinfection treatments, the procedures used by ICNF, I.P., for plant certification, considering the aforementioned parameters, were applied. For that, plants from each trial were observed 7 months after sowing to assess their average height, average collar diameter, and the presence or absence of any deficiencies. Statistical analysis of the data obtained in nursery A for the germination results was undertaken via a t test and via one-way analysis of variance (ANOVA) for nursery B. For the latter, whenever significant differences were found (p < 0.05), post hoc Tukey’s pairwise test was performed to further elucidate differences between treatments at a significance level of α = 0.05 using PAST software, version 4.03 [32].
2.6. In Vitro Germination Assay
Concurrently with the field trials, the top-performing treatments identified in the previous disinfection stage were also subjected to a preliminary germination assay to evaluate their impacts on P. pinaster seed germination. This assessment was conducted by CENASEF in accordance with the standards of the International Seed Testing Association (ISTA, https://www.seedtest.org/en/home.html (accessed on 4 June 2020)). The general conditions of the germination tests were as follows: 200 seeds in sets of 100 seeds each; filter paper substrate soaked in water; temperature set at 20 °C; humidity maintained at 85% inside the germination chamber; photoperiod of 8 h of light followed by 16 h of darkness; and a test duration of 35 days with counts performed every 7 days. Finally, the germination rate was assessed for all treatments and statistically analyzed using one-way analysis of variance (ANOVA).
An important note regarding the field trials and their respective germination tests must be highlighted. Both trials were conducted during the period declared as a pandemic due to the ongoing COVID–19 crisis. Consequently, experiments involving ethanol at various concentrations, previously conducted in the laboratory, could not be reproduced in the field because of limited access to ethanol. Therefore, although promising results were obtained in the laboratory, it was decided to retain this information, despite the inability to conduct further tests. Consequently, no additional conclusions could be drawn regarding its impact on germination or its performance in the field.
3. Results and Discussion
3.1. Effect of Treatments on Fusarium Circinatum-Contaminated Pinus Seeds
The isolation of F. circinatum from seeds was assessed for each Pinus species and treatment. The resulting data, showing the average number of contaminated seeds for each combination, are presented in Figure 3 and in Supplementary Table S2.
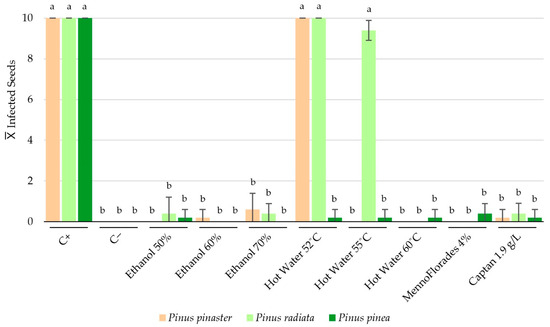
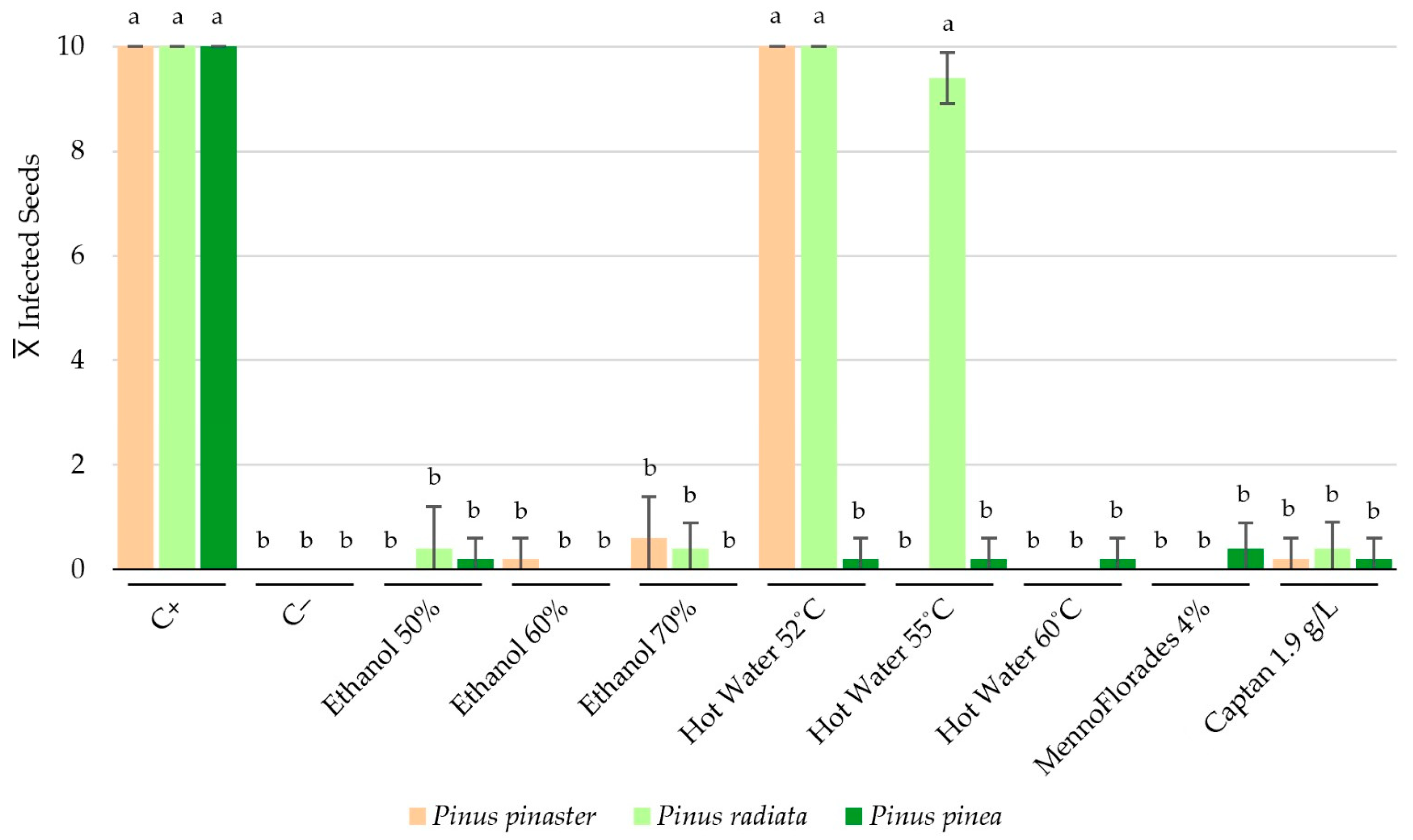
Figure 3.
Average numbers of Pinus pinaster, P. radiata and P. pinea seeds contaminated with F. circinatum after treatment application. Error bars shown in the graph correspond to the standard deviations. For each species, data (n = 10) associated with different letters represent significantly different results in accordance with Dunn’s post hoc test, p < 0.01.
As observed, multiple treatments per species achieved a 100% seed disinfection rate, significantly reducing F. circinatum contamination. For P. pinaster, treatments with 50% ethanol (v/v), hot water at 55 °C and 60 °C, and MennoFlorades at a 4% (v/v) concentration achieved complete disinfection in all replicates. The statistical analysis indicated that there were no significant differences between the following treatments: ethanol at 60% and 70% concentrations, 1.9 g/L Captan, and the negative control. Additionally, significant differences were found between the previous group and the group composed of the positive control and hot water at 52 °C treatment (p < 0.01). A similar trend was seen in P. radiata seeds, where hot water at 60 °C, MennoFlorades at 4% (v/v), and 60% ethanol (v/v) all showed no F. circinatum growth. The data analysis confirmed that there were no significant differences between the following treatments: ethanol at 50% and 70%, 1.9 g/L Captan, and the negative control. Furthermore, significant differences were observed between the previous group and a second data group comprising hot water at 52 °C and 55 °C, and the positive control (p < 0.01). For P. pinea seeds, the statistical analysis identified two significantly different groups: one solely comprising the positive control and another including all evaluated treatments and the negative control (p < 0.01). In the latter group, only treatments with 60% and 70% ethanol (v/v) showed no F. circinatum colonies. An interesting aspect of these results is the apparent importance of seed morphology. Pinus pinea seeds, which are larger and possess smoother surfaces, responded better to all treatments, with each treatment significantly reducing the number of contaminated seeds. In contrast, P. pinaster and P. radiata seeds, which are smaller and have more rugged surfaces, showed varied responses to two of the three tested temperatures for the hot water treatments. For ethanol, Captan, and MennoFlorades treatments, similar results were observed across all concentrations and Pinus species.
Previous studies on disinfection of Pinus seeds contaminated with F. circinatum have explored various methods, including hydrogen peroxide [40], non-thermal plasma [41], commercial fungicides, antagonistic organisms, thermotherapy, and essential oils [42]. Non-thermal plasma seed disinfection, despite showing promising results, was found to negatively affect P. radiata seed germination when exposed for 60 s or more [41]. However, this negative impact was not observed in P. sylvestris seeds contaminated with F. oxysporum [43]. Nevertheless, implementing non-thermal plasma in a nursery setting would be impractical. The use of antagonistic organisms like Trichoderma spp. has proven to be ineffective in seed disinfection, despite their effectiveness in in vitro plate assays [42]. Essential oils have also shown generalized seed germination inhibition, limiting their potential as viable treatments [42].
Among the treatments tested in this study, ethanol, particularly at a 60% concentration, effectively eliminated the F. circinatum inoculum from all three tested Pinus species. This efficacy aligns with previous reports and is attributed to protein denaturation and the disruption of cell membranes [28,41]. The results of hot water treatments were similar to those of previous studies [30,31], but we found that higher temperatures (above 60 °C) yielded the best outcomes. This indicates the potential of hot water treatments for use in Pinus plant production to prevent phytopathogenic fungi infection, similar to their successful application in vineyard production where they effectively eliminate fungal pathogens from grapevine cuttings [29]. The efficacy of Captan, known for its fungicidal activity against F. circinatum [42], was confirmed in our assays. MennoFlorades also demonstrated significant results across all tested seeds, suggesting that its metabolic inhibition, primarily due to benzoic acid [26], effectively suppresses fungal growth, as previously reported [44], including F. circinatum growth.
Across all Pinus species, treatments including 60% ethanol, hot water at 60 °C, MennoFlorades at 4% (v/v), and Captan at 1.9 g/L showed promising results and were selected for further assays to assess their impacts on germination both in vitro and in a nursery context. However, due to an ethanol shortage mentioned in Section 2.6, it was not feasible to continue using it in subsequent germination tests. Therefore, only hot water at 60 °C, MennoFlorades at 4% (v/v), and Captan at 1.9 g/L were utilized for the subsequent steps of the study.
3.2. Isolated and Identified Non-Fusarium Species
On P. pinaster seeds, the identified isolates included Aspergillus sp., Botrytis cinerea, and Leptobacillium chinense. Botrytis cinerea was only detected in the negative control, while L. chinense was found only in the ethanol 50% treatment. Aspergillus sp. was found in all treatments except for the positive control, hot water at 52 °C, and 1.9 g/L Captan. The absence of B. cinerea on treated seeds, despite its presence in the negative control, is noteworthy, as this species can pose a threat to nurseries due to its ability to infect container grown seedlings, potentially leading to their death [45].
Pinus radiata seeds showed higher fungal diversity, with 4% MennoFlorades being the only treatment with no fungal growth. The following species were identified on P. radiata seeds: Echria macrotheca, Irpex laceratus, Chromelosporiopsis carneum, Cladosporium cladosporioides, Neurospora crassa, Peziza varia, Pezizaceae sp., Aequabiliella effusa, Sistotrema sp., Coniochaeta decumbens, Coniochaeta hoffmannii, Coniochaeta acaciae, and Trichodelitschia bisporula. None of these species have been reported as potential Pinus pathogens, with most being endophytes or saprophytes [46,47,48,49,50,51,52,53].
No fungal growth was detected on P. pinea seeds, aside from the F. circinatum colonies and some Trichoderma sp. isolates on the negative control.
3.3. Nursery Germination Assays
To evaluate the impacts of the selected treatments (4% MennoFlorades, hot water at 60 °C, and 1.9 g/L Captan) on Pinus pinaster seed germination and plant quality, large-scale germination assays were conducted in partnering nurseries. The germination rate for each treatment was determined at 1 and 2 months post-sowing, with a control group of non-treated seeds included in each nursery, and these results are presented in Table 3 and in Supplementary Materials Table S3.
Table 3.
Average germination rate (%) after Pinus pinaster seed disinfection with the different treatments in nurseries at 1 and 2 months post-sowing. In each column, data followed by different letters were significantly different according to Tukey’s pairwise test (p < 0.05).
In nursery A, the germination rate for seeds treated with MennoFlorades were not significantly different from the control group in both months. In contrast, in nursery B, seeds treated with 1.9 g/L Captan had a significantly lower germination rate compared to the control group. The hot water treatment, while not significantly different from the control group, showed an average germination rate of 59.03% at two months post-sowing, compared to 55.21% observed in the control group. The overall low germination rate observed across both nurseries, for both treated and non-treated seeds, is likely due to the late sowing in June instead of the usual sowing in March as a result of the COVID-19 pandemic and associated restrictions. This delay, combined with the extremely high temperatures during the summer period, negatively impacted germination rates, a phenomenon also observed in our irrigation water nursery assays [21].
The plant certification process evaluated the P. pinaster plants that resulted from the germination of treated and control seeds in both nurseries. This evaluation took place 7 months post-sowing and assessed compliance with several standards defined by the applicable Portuguese legislation, as mentioned previously in Section 2.5. For P. pinaster plants, the species-specific parameters include a maximum age of 1 year, a minimum height of 7 cm, a maximum height of 30 cm, and a minimum root collar diameter of 2 mm. The results of the plant certification evaluation are presented in Table 4.
Table 4.
Number of certified Pinus pinaster plants and respective parameters per nursery and treatment.
In nursery B, a high percentage of plants met the certification standards across all treatments and the control group. The lowest certification rate was observed in plants from 1.9 g/L Captan-treated seeds, where approximately 86% of the plants fulfilled the certification requirements. The mean height of the plants was significantly above the legal minimum of 7 cm, and the mean diameter was equal to or greater than the legal minimum of 2 mm for all tests. Conversely, in nursery A, the number of certified plants was significantly lower. Only ten plants from the control group met the certification parameters, and no plants from the MennoFlorades treatment did so. Both the mean height and diameter were below the legal minimums in both the treatment and control groups. The late sowing and intense heat during the period are likely reasons for these poor results obtained from this nursery. However, it remains inconclusive whether MennoFlorades would negatively affect the certification of Pinus seedlings under regular conditions.
Unlike a different study that had previously reported on P. radiata seeds treated with hot water, where the germination percentage was as low as 45%–50%, even at lower temperatures (54 °C) [30], our study showed promising results for P. pinaster seeds. These differences could be attributed to the shorter exposure time of seeds to high temperatures, which was one-half to one-third of that tested by Agustí-Brisach et al. [30]. Among all the treatments, hot water at 60 °C for 15 min demonstrated no negative impacts on seed germination or the percentage of certified plants and had a negligible effect on the mean height and diameter of the resulting plants. These findings, combined with its low economic impact for forest reproductive material (FRM) producers and its eco-friendly nature, make this treatment an ideal candidate for application in nurseries as a preventive measure against F. circinatum.
3.4. In Vitro Germination Assays
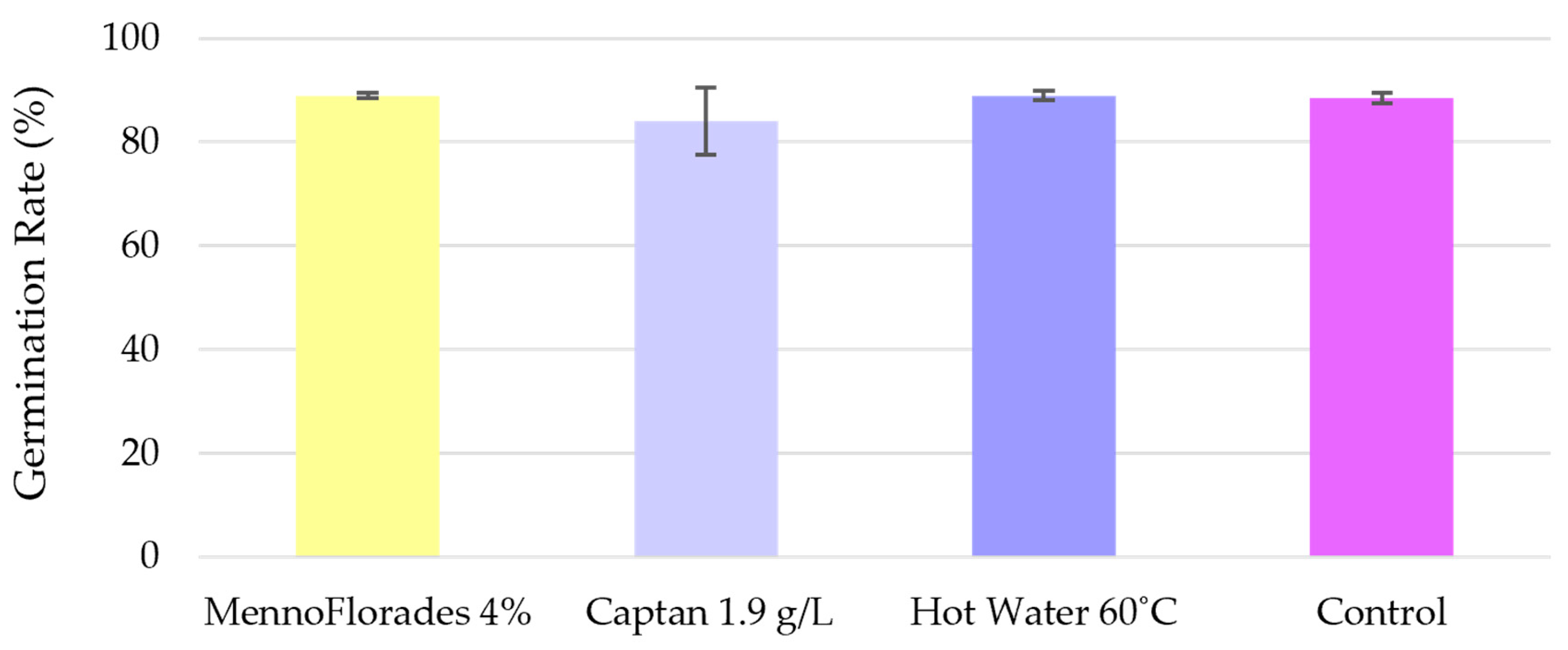
Alongside the nursery trials, in vitro germination assays were conducted by CENASEF in accordance with ISTA standards. The treatments evaluated in these assays were the same as those tested in the nurseries: MennoFlorades at 4% (v/v), Captan at 1.9 g/L, and hot water at 60 °C. The results, shown in Figure 4 and in Supplementary Table S4, indicate that the germination rate was close to 90% for most treatments and the control group, with no significant differences between them (F = 3.87; df = 2.09; p = 0.20), with 1.9 g/L Captan showing an average germination rate of 84% compared to the 88.5% verified for the control group. These findings are consistent with the ones of the nursery assays, where Captan was the only treatment resulting in a pointedly lower germination rate compared to the control. Moreover, hot water treatment did not negatively impact seed germination, corroborating our nursery results. This is further supported by the study of Jones et al. [54], which reported similar outcomes for seeds of another Pinus species, namely, P. palustris. However, extended periods of hot water treatment (30 to 45 min), even at lower temperatures (50–54 °C), have been reported to significantly impact P. radiata seed germination [30].
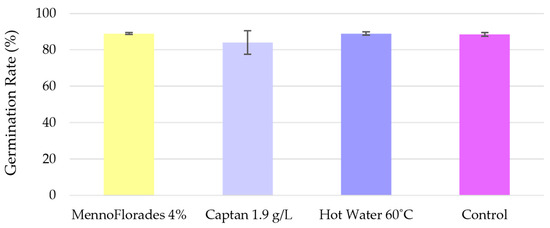
Figure 4.
Average germination rate of Pinus pinaster per treatment from the in vitro CENASEF assays. Error bars shown in the graph correspond to the standard deviations.
Higher germination rates were observed in the in vitro tests compared to the nursery assays, which was expected given that ISTA standards ensure ideal conditions for seed germination, conditions that are not always replicable in a real nursery context.
4. Conclusions
Given the significance of Pinus forests to the Portuguese environment and economy, any strategy that can mitigate threats like potential contamination by Fusarium circinatum is highly valuable. This is particularly true if the strategy is effective, environmentally friendly, and economically viable on a large scale, such as in nurseries where most forest reproductive material (FRM) for reforestation is produced.
Our study took an innovative lab to nursery approach, thus ensuring that the strategies developed were based on scientific data and results, but always working in close collaboration with FRM producers to ensure the viability of said strategies in practical and economical contexts. This approach allowed us to identify several promising treatments for seed disinfection that can be implemented in nurseries to prevent the spread of pine pitch canker. When combined with other measures, such as irrigation water disinfection, alternative substrates, as presented in our previously published work [21,22,23,24], and good nursery practices, these treatments can significantly reduce the spread of F. circinatum in both nurseries and forests. A technical manual based on our findings was composed [55] and distributed to FRM producers’ nationwide, who are already implementing some of the suggested strategies to prevent F. circinatum outbreaks in their nurseries, thus ensuring healthier and more resilient pine populations and contributing to the long-term sustainability of forest ecosystems.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/f15071154/s1, Table S1: Origin and GenBank accession numbers of all non-Fusarium isolates obtained in this study; Table S2: Numbers of Pinus pinaster, P. radiata, and P. pinea seeds contaminated with Fusarim circinatum after disinfection treatment application; Table S3: Germination rate (%) after Pinus pinaster seed disinfection with the different treatments in nurseries at 1 and 2 months post-sowing; Table S4: Germination rate of Pinus pinaster per treatment from the in vitro CENASEF assays.
Author Contributions
Conceptualization, L.F., D.S.P., D.R., L.M., H.B. and A.P.; methodology, L.F., D.S.P., A.C.S., C.F. and A.R.F.; software, L.F. and D.S.P.; validation, L.F., D.S.P., A.C.S., D.R., L.M., H.B. and A.P.; formal analysis, L.F. and D.S.P.; investigation, L.F., D.S.P., A.C.S., C.F. and A.R.F.; resources, D.R., L.M., H.B. and A.P.; data curation, L.F. and D.S.P.; writing—original draft preparation, L.F. and D.S.P.; writing—review and editing, L.F., D.S.P. and A.P.; visualization, L.F. and D.S.P.; supervision, D.R., L.M., H.B. and A.P.; project administration, D.R.; funding acquisition, D.R., L.M., H.B. and A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-funded by PDR2020, Portugal2020 and FEDER under the framework GO +PrevCRP (PDR2020–101–031058, partnership No. 112, initiative No. 237) and PRR—Recovery and Resilience Plan and by the NextGeneration EU European Funds. Diana Paiva was supported by PhD research grant UI/BD/150843/2021 (https://doi.org/10.54499/UI/BD/150843/2021) awarded by the Centre for Functional Ecology—Science for People and the Planet (CFE) and co–funded by Fundação para a Ciência e Tecnologia, I.P. (FCT) through national funding by the Ministério da Ciência, Tecnologia e Ensino Superior (MCTES) from Fundo Social Europeu (FSE). Ana C. Silva was financially supported by PhD research grant 2022.11826.BD. Helena Bragança acknowledges support from UIDB/04551/2020 and UIDP/04551/2020 center grants to GREEN–IT. The authors thank FCT for funding the R&D Unit Centre for Functional Ecology—Science for People and the Planet (CFE) with reference UIDB/04004/2020 and DOI identifier 10.54499/UIDB/04004/2020 (https://doi.org/10.54499/UIDB/04004/2020) and the Associate Laboratory TERRA with reference LA/P/0092/2020 and DOI identifier 10.54499/LA/P/0092/2020 (https://doi.org/10.54499/LA/P/0092/2020), financed by FCT/MCTES through national funds (PIDDAC).
Data Availability Statement
The original contributions presented in the study are included in the article and Supplementary Materials, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank the nurseries APFC-Equipar, Germiplanta, ICNF-Alcácer do Sal, and ICNF-Amarante for their collaboration in this study and for providing the project with the necessary infrastructure and manpower to conduct these assays. The authors also thank Eugénio Diogo from INIAV for his collaboration. Photographs included in Figure 1 and Figure 2 were captured by the article’s authors, Luís Fernandes and Diana Paiva, and by Dina Ribeiro from ICNF, respectively.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gordon, T. Pitch Canker Disease of Pines. Phytopathology 2006, 96, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Bezos, D.; Martinez-Alvarez, P.; Fernandez, M.; Diez, J. Epidemiology and Management of Pine Pitch Canker Disease in Europe—A Review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Drenkhan, R.; Ganley, B.; Martín-García, J.; Vahalík, P.; Adamson, K.; Adamčíková, K.; Ahumada, R.; Blank, L.; Bragança, H.; Capretti, P.; et al. Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker. Forests 2020, 11, 724. [Google Scholar] [CrossRef]
- Schmale, D.G., III; Gordon, T.R. Variation in susceptibility to pitch canker disease, caused by Fusarium circinatum, in native stands of Pinus muricata. Plant Pathol. 2003, 52, 720–725. [Google Scholar] [CrossRef]
- Gordon, T.R.; Kirkpatrick, S.C.; Aegerter, B.J.; Wood, D.L.; Storer, A.J. Susceptibility of Douglas fir (Pseudotsuga menziesii) to pitch canker, caused by Gibberella circinata (anamorph = Fusarium circinatum). Plant Pathol. 2006, 55, 231–237. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- Solel, Z.; Bruck, R.I. Relation between wilt rate and obstruction of water flow in stems of two families of loblolly pine affected by pitch canker. Eur. J. For. Pathol. 1990, 20, 317–320. [Google Scholar] [CrossRef]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of some southern pines. J. For. 1946, 44, 724–744. [Google Scholar]
- Berbegal, M.; Pérez-Sierra, A.; Armengol, J.; Grünwald, N.J. Evidence for Multiple Introductions and Clonality in Spanish Populations of Fusarium circinatum. Phytopathology 2013, 103, 851–861. [Google Scholar] [CrossRef]
- Landeras, E.; Garcia, P.; Fernández, Y.; Braña, M.; Fernández-Alonso, O.; Méndez-Lodos, S.; Pérez-Sierra, A.; Leon, M.; Abad-Campos, P.; Berbegal, M.; et al. Outbreak of Pitch Canker Caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- Dwinell, D. Global Distribution of the Pitch Canker Fungus. In Current and Potential Impacts of Pitch Canker in Radiata Pine; IMPACT Monterey Workshop: Monterey, CA, USA; Citeseer: Princeton, NJ, USA, 1999; Volume 30, pp. 54–57. [Google Scholar]
- European and Mediterranean Plant Protection Organization (EPPO) Global Database. First Report of Gibberella circinata in France; EPPO: Paris, France, 2006; p. 104. [Google Scholar]
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First report of Pitch Canker caused by Fusarium circinatum on Pinus halepensis and P. pinea in Apulia (southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar] [CrossRef]
- Bragança, H.; Diogo, E.; Moniz, F.; Amparo, P. First report of Pitch canker on Pines caused by Fusarium circinatum in Portugal. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef]
- 6º Inventário Florestal Nacional—IFN6. Available online: https://inforcna.pt/Media/Files/201979_Ifn6PrincipaisResultadosJun2019.pdf (accessed on 13 May 2024).
- Instituto Nacional de Estatística—Statistics Portugal, Estatísticas do Comércio Internacional. Available online: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_publicacoes&PUBLICACOESpub_boui=280862956&PUBLICACOESmodo=2 (accessed on 13 May 2024).
- Vettraino, A.; Potting, R.; Raposo, R. EU legislation on forest plant health: An overview with a focus on Fusarium circinatum. Forests 2018, 9, 568. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO) Global Database. EPPO Datasheet: Fusarium circinatum. Available online: https://gd.eppo.int/taxon/GIBBCI/datasheet (accessed on 13 May 2024).
- European Food and Environment Safety Agency (EFSA) Panel on Plant Health (PLH). Risk assessment of Gibberella circinata for the EU territory and identification and evaluation of risk management options. EFSA J. 2010, 8, 1620. [Google Scholar] [CrossRef]
- Fernandes, L.; Paiva, D.; Roxo, I.; Fernandes, A.R.; Ribeiro, D.; Ribeiro, H.; Portugal, A. Development of New Preventive Strategies for Pine Pitch Canker Caused by Fusarium circinatum in Irrigation Water and Evaluation in a Real Nursery Context. Forests 2023, 14, 443. [Google Scholar] [CrossRef]
- Silva, A.C.; Diogo, E.; Bragança, H. Effect of Substrate Solarization for the Control of Fungi: The Case Study of Fusarium circinatum, the Quarantine Agent of Pine Pitch Canker. Silva Lusit. 2021, 29, 161–175. [Google Scholar] [CrossRef]
- Chemetova, C.; Quilhó, T.; Braga, S.; Fabião, A.; Gominho, J.; Ribeiro, H. Aged Acacia melanoxylon bark as an organic peat replacement in container media. J. Clean. Prod. 2019, 232, 1103–1111. [Google Scholar] [CrossRef]
- Chemetova, C.; Mota, D.; Fabião, A.; Gominho, J.; Ribeiro, H. Low-temperature hydrothermally treated Eucalyptus globulus bark: From by-product to horticultural fiber-based growing media viability. J. Clean. Prod. 2021, 319, 128805. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). 91 (2) Fusarium circinatum (formerly Giberella circinata). EPPO Bull. 2019, 49, 228–247. [Google Scholar] [CrossRef]
- Krebs, H.A.; Wigginst, D.; Stubbs, M. Studies on the mechanism of the antifungal action of benzoate. Biochem. J. 1983, 7, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.D. Action of the Fungicides Captan and Folpet on Rat Liver Mitochondria. Biochem. Pharmacol. 1971, 20, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.; Dantigny, P. Control of food spoilage fungi by ethanol. Food Control 2011, 22, 360–368. [Google Scholar] [CrossRef]
- Crous, P.; Swart, L.; Coertze, S. The effect of hot-water treatment on fungi occurring in apparently healthy grapevine cuttings. Phytopathol. Mediterr. 2001, 40, S464–S466. [Google Scholar]
- Agustí-Brisach, C.; Pérez-Sierra, A.; Armengol, J.; García-Jiménez, J.; Berbegal, M. Efficacy of hot water treatment to reduce the incidence of Fusarium circinatum on Pinus radiata seeds. Forestry 2012, 85, 629–635. [Google Scholar] [CrossRef]
- Liao, T.; Ye, J.; Chen, J.; Han, Y.; Wu, C.; An, Y. Biological characteristics of Fusarium circinatum. J. Nanjing For. Univ. 2008, 32, 83–86. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Paiva, D.S.; Fernandes, L.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Uncovering the Fungal Diversity Colonizing Limestone Walls of a Forgotten Monument in the Central Region of Portugal by High-Throughput Sequencing and Culture-Based Methods. Appl. Sci. 2022, 12, 10650. [Google Scholar] [CrossRef]
- Paiva, D.S.; Fernandes, L.; Pereira, E.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal). J. Fungi 2023, 9, 501. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Robert, V.; Stegehuis, G.; Stalpers, J. The MycoBank Engine and Related Databases. 2005. Available online: https://www.MycoBank.org/ (accessed on 13 May 2024).
- Dwinell, L.D. Contamination of Pinus radiata seeds in California by Fusarium circinatum. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, States Department of Agriculture (USDA) Forest Service, San Diego, CA, USA; 1999; pp. 1–4. [Google Scholar]
- Šerá, B.; Zahoranová, A.; Bujdáková, H.; Šerý, M. Disinfection from pine seeds contaminated with Fusarium circinatum Nirenberg & O’Donnell using non-thermal plasma treatment. Rom. Rep. Phys 2019, 71, 701. [Google Scholar]
- Iturritxa, E.; Desprez-Loustau, M.L.; García-Serna, I.; Quintana, E.; Mesanza, N.; Aitken, J. Effect of alternative disinfection treatments against fungal canker in seeds of Pinus radiata. Seed Technol. 2011, 33, 88–110. [Google Scholar]
- Świecimska, M.; Tulik, M.; Šerá, B.; Golińska, P.; Tomeková, J.; Medvecká, V.; Bujdáková, H.; Oszako, T.; Zahoranová, A.; Šerý, M. Non-Thermal Plasma Can Be Used in Disinfection of Scots Pine (Pinus sylvestris L.) Seeds Infected with Fusarium oxysporum. Forests 2020, 11, 837. [Google Scholar] [CrossRef]
- Gioia, T.; Galinski, A.; Lenz, H.; Müller, C.; Lentz, J.; Heinz, K.; Nagel, K.A. GrowScreen-PaGe, a non-invasive, high-throughput phenotyping system based on germination paper to quantify crop phenotypic diversity and plasticity of root traits under varying nutrient supply. Funct. Plant Biol. 2016, 44, 76–93. [Google Scholar] [CrossRef]
- Capieau, K.; Stenlid, J.; Stenström, E. Potential for biological control of Botrytis cinerea in Pinus sylvestris seedlings. Scand. J. For. Res. 2004, 19, 312–319. [Google Scholar] [CrossRef]
- Jiao, Y.; Huang, J. Allelopathic effects of aqueous extracts from uncomposted and composted Mexican devil (Ageratina adenophora) plants on forest fungal growth and soil nitrogen and phosphorus mobilization. Weed Sci. 2024, 72, 76–85. [Google Scholar] [CrossRef]
- Lygis, V.; Vasiliauskaite, I.; Matelis, A.; Pliūra, A.; Vasaitis, R. Fungi in living and dead stems and stumps of Pinus mugo on coastal dunes of the Baltic Sea. Plant Protect. Sci. 2014, 50, 221–226. [Google Scholar] [CrossRef]
- Lygis, V.; Vasiliauskaite, I.; Stenlid, J.; Vasaitis, R. Impact of forest fire on occurrence of Heterobasidion annosum ss root rot and other wood–inhabiting fungi in roots of Pinus mugo. Forestry 2010, 83, 83–92. [Google Scholar] [CrossRef]
- Stępniewska, S.; Mańka, M. Fungi inhabiting Scots pine (Pinus sylvestris) seeds in Wielkopolska region. Phytopathol. Pol. 2004, 31, 73–76. [Google Scholar]
- Sanz–Ros, A.V.; Müller, M.M.; San Martín, R.; Diez, J.J. Fungal endophytic communities on twigs of fast and slow growing Scots pine (Pinus sylvestris L.) in northern Spain. Fungal Biol. 2015, 119, 870–883. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.C.; Hui, S.; Choi, J.; Asiegbu, F.O.; Valkonen, J.P.; Lee, Y.H. Secret lifestyles of Neurospora crassa. Sci. Rep. 2014, 4, 5135. [Google Scholar] [CrossRef] [PubMed]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Garay–Serrano, E.; del Pilar Ortega–Larrocea, M.; Reverchon, F.; Suárez–Quijada, I. Persistence of ecto– and ectendomycorrhizal fungi associated with Pinus montezumae in experimental microcosms. Symbiosis 2018, 74, 67–78. [Google Scholar] [CrossRef]
- Jones, J.P.; Sun, X.; Eckhardt, L.; Webber, A.; Hess, N.; Barnett, J.; McGilvary, J. Longleaf seedling production: Some problems and their solutions. LA Agric. 2002, 45, 4–11. [Google Scholar]
- Fernandes, A.R.; Silva, A.C.; Portugal, A.; Fernandes, C.; Ribeiro, D.; Bragança, H.; Ribeiro, H.; Barbosa, J.N.; Fernandes, L.; Martins, L.; et al. Prevenção do Cancro-Resinoso-do-Pinheiro (Fusarium circinatum)-Manual Técnico para Fornecedores de Materiais Florestais de Reprodução, 1st ed.; Centro PINUS: Viana do Castelo, Portugal, 2021. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).