
Mechanism of the Effects of Phyllostachys edulis Invasion on the Soil Microbial Community in Quercus acutissima Forests

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Sample Collection and Analytical Methods
2.3.1. Sample Collection
2.3.2. Soil Physicochemical Index Determination
2.3.3. DNA Extraction and PCR Amplification
2.3.4. Statistical Analysis
3. Results
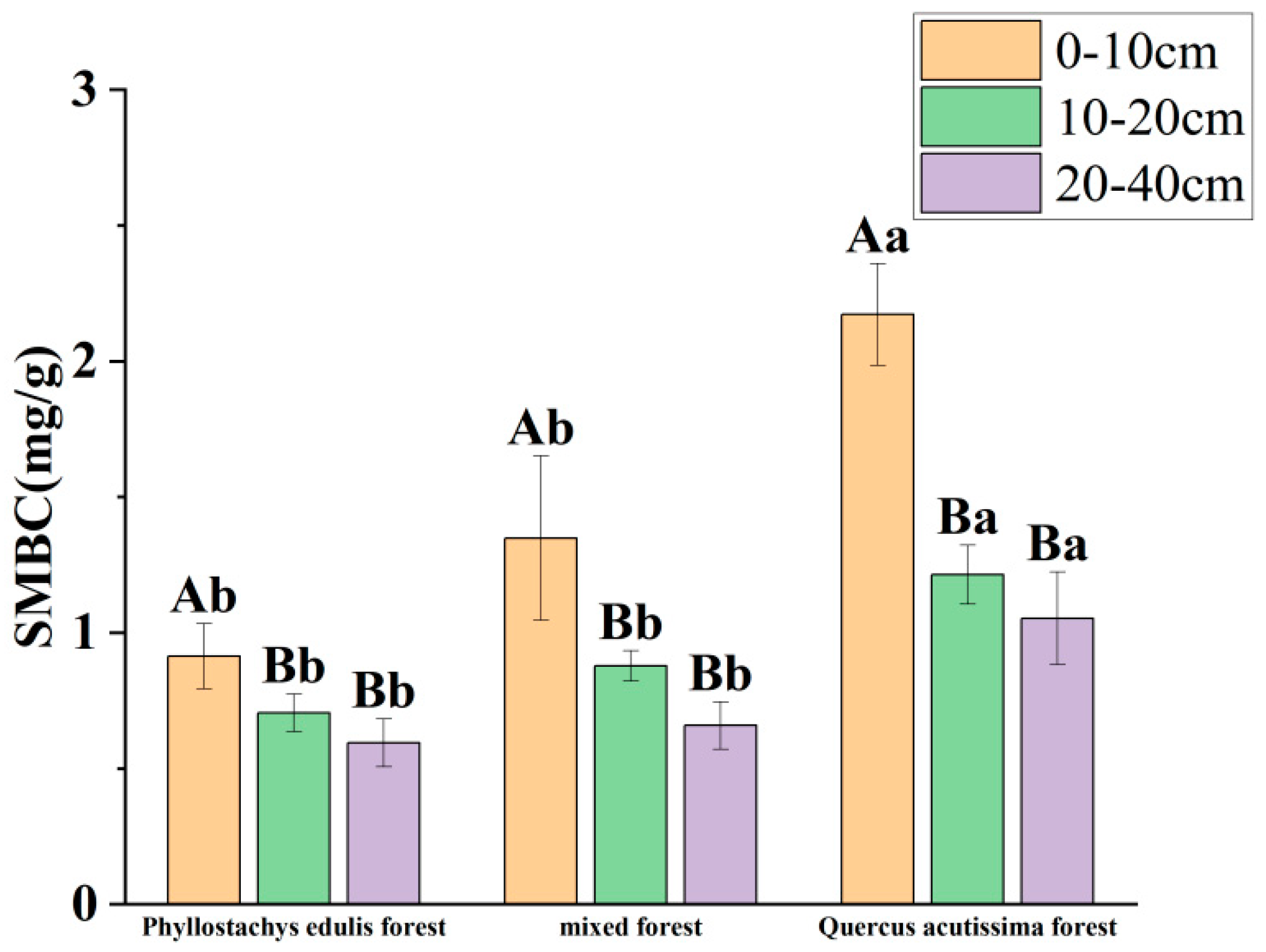
3.1. Effects of Phyllostachys edulis Invasion on Soil Physicochemical Properties
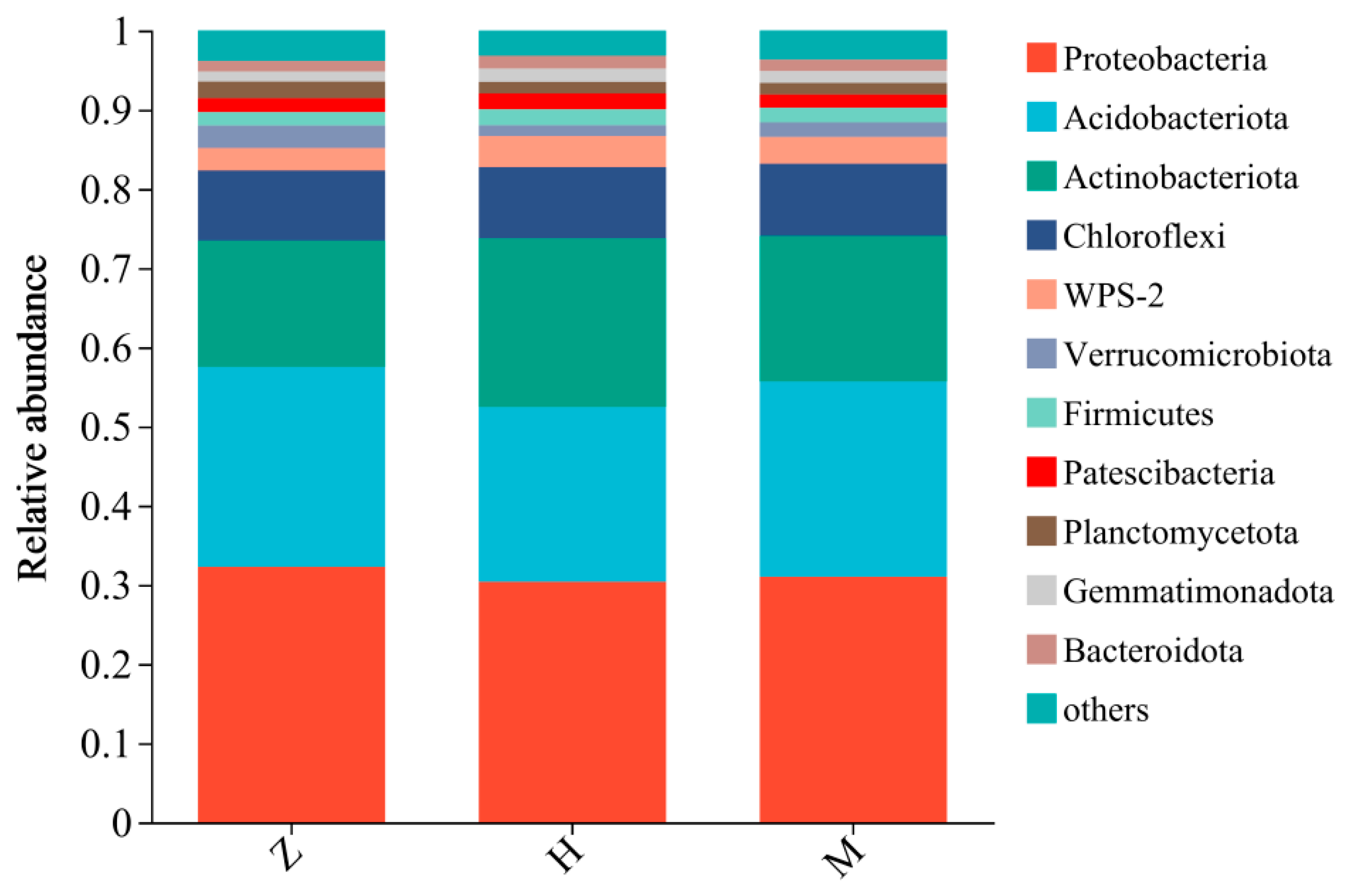
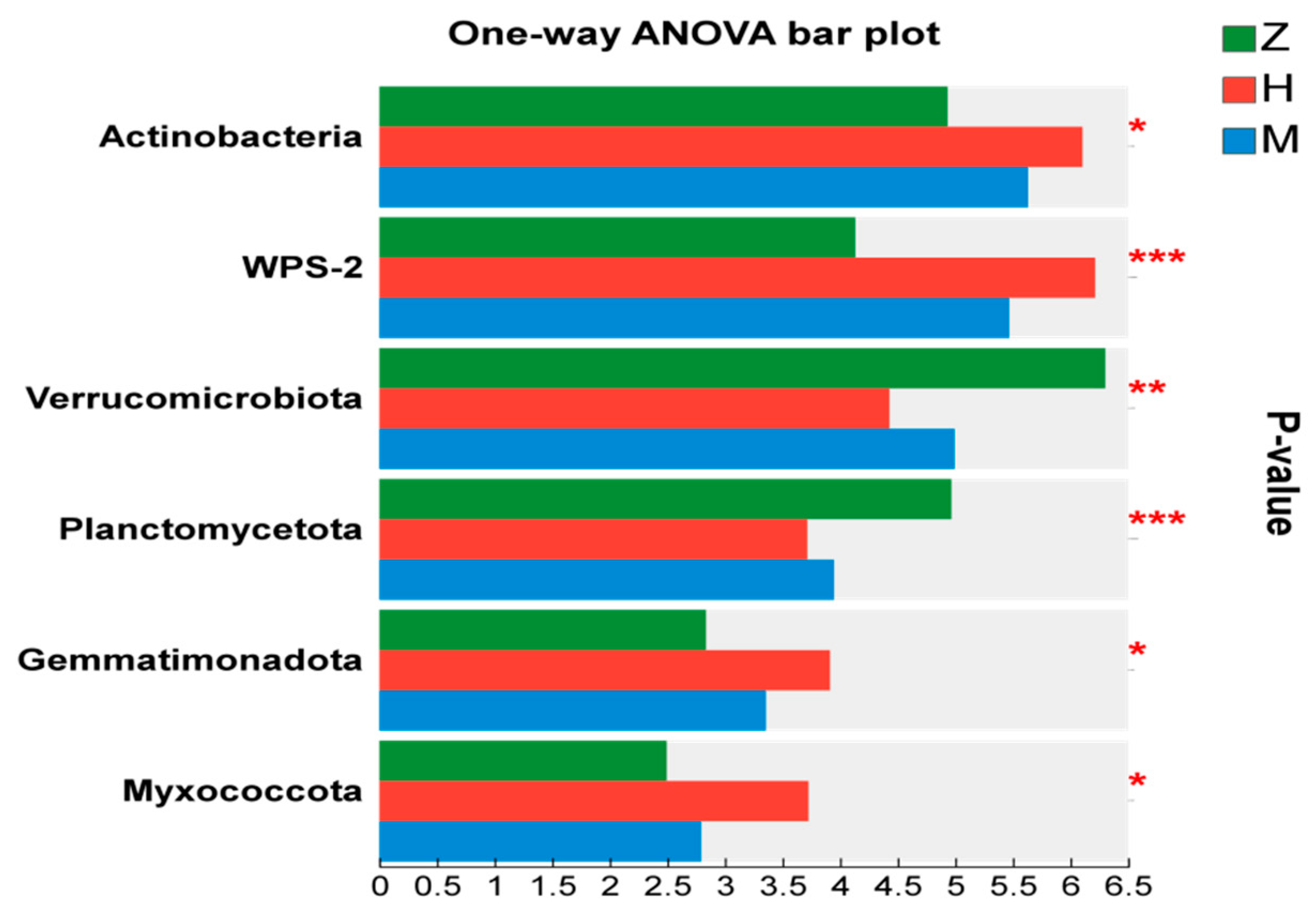
3.2. Effects of Phyllostachys edulis Invasion on Soil Microbial Communities
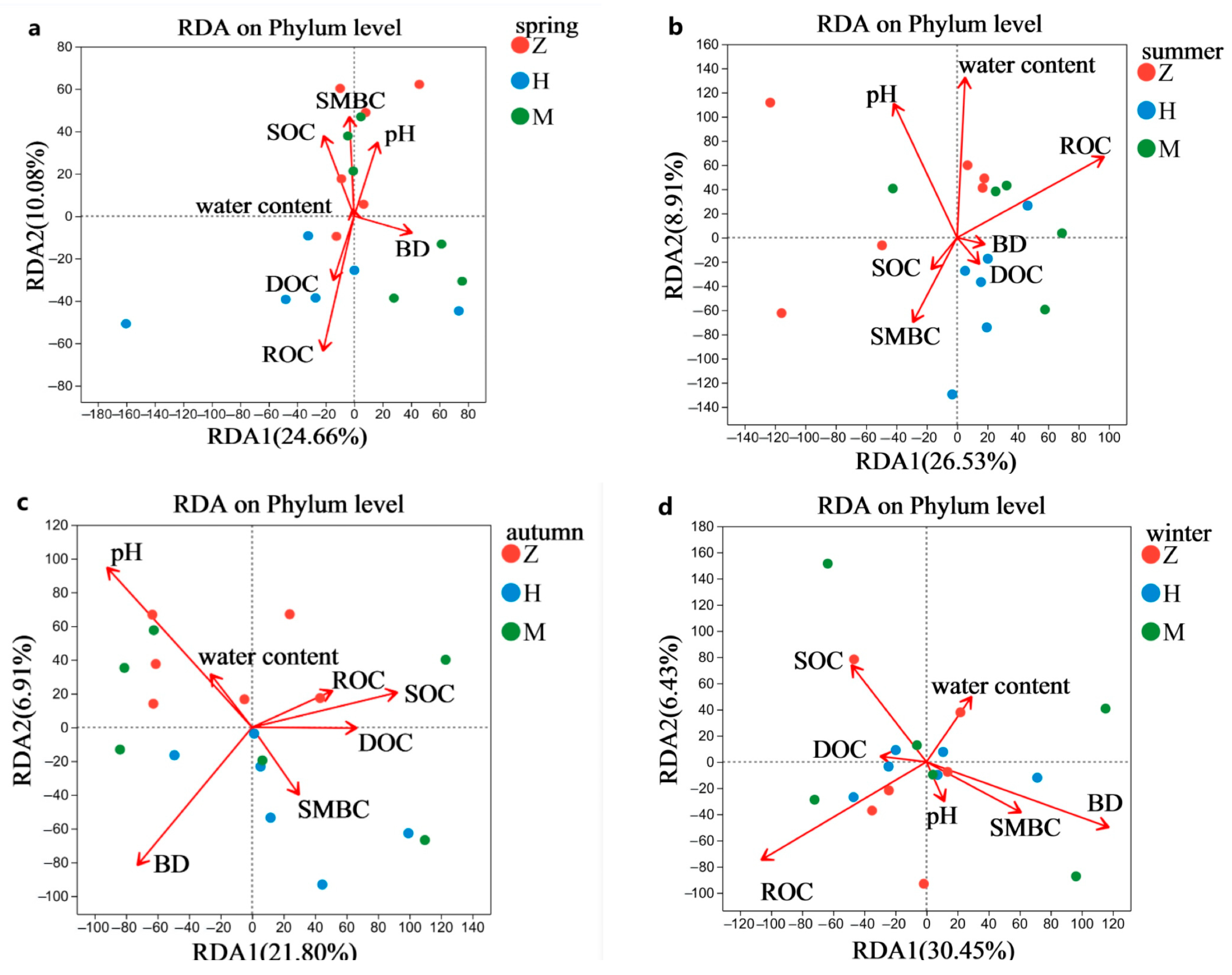
3.3. Association between the Soil Microbial Community and Environmental Factors in Phyllostachys edulis-Invaded Soils
4. Discussion
4.1. Effects of Phyllostachys edulis Invasion on Soil Physicochemical Properties
4.2. Effects of Phyllostachys edulis Invasion on the Soil Microbial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ju, R.T.; Li, H.; Shi, Z.R.; Li, B. Progress of biological invasion research in China in the last decade. Biodiversity 2012, 20, 581–611. [Google Scholar]
- Song, Q.N.; Yang, Q.P.; Liu, J.; Yu, D.K.; Fang, K.; Xu, P.; He, Y.J. Effects of moso bamboo expansion on soil nitrogen mineralization and effectiveness in broad-leaved evergreen forest. J. Appl. Ecol. 2013, 24, 338–344. [Google Scholar]
- Ming, O.Y.; Di, T.; Jiamin, P.; Chen, G.P.; Su, H.J.; Yan, Z.B.; Yang, Q.P.; Ji, C.J.; Tang, Z.Y.; Fang, J.Y. Moso bamboo (Phyllostachys edulis) invasion increases forest soil pH in subtropical China. Catena 2022, 215, 106339. [Google Scholar]
- Wang, S.; Chen, T.H.; Liu, E.U.; Liu, C.P. Accessing the nursing behaviour of Moso bamboo (Phyllostachys edilus) on carbohydrates dynamics and photosystems. Sci. Rep. 2020, 10, 1015. [Google Scholar] [CrossRef] [PubMed]
- Dutta, K.; Reddy, C.S. Geospatial analysis of reed bamboo(Ochlandra travancorica) invasion in Western Ghats, India. J. Indian Soc. Remote Sens. 2016, 44, 699–711. [Google Scholar] [CrossRef]
- Tong, F.G.; Wu, Z.H.; Lin, R.X.; Wu, X.J.; Deng, H.F.; Huang, Z.J.; Li, R.J.; Luan, J.W. Effects of Phyllostachys expansion on soil hopping community characteristics of evergreen broad-leaved forests in South Asia. For. Surv. Plan Cent. South. China 2024, 43, 51–57+70. [Google Scholar]
- Wei, Q.Y.; Yin, R.; Huang, J.H.; Alfried, P.V.; Li, Y.C.; Miao, X.Q.; Paul, K. The diversity of soil mesofauna declines after bamboo invasion in subtropical China. Sci. Total Environ. 2021, 789, 147982. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.L.; Fan, S.H.; Qi, L.H.; Guan, F.Y.; Cai, C.J.; Du, M.Y. Soil respiration and carbon balance in a Moso bamboo(Phyllostachys heterocycla(Carr.) Mitford cx. Pubescens) forest in subtropical China. Iforest-Biogeosciencesand For. 2015, 8, 606–614. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Huang, W.Z. Effects of Exotic Plant Species on Soil Properties in Hardwood Forests of New Jersey. Water Air Soil Pollut. 1998, 105, 493–501. [Google Scholar] [CrossRef]
- Song, C.; Chi, X.C.; Wang, X.Y.; Wang, Y.X.; Ye, J.W.; Zhu, X.T.; Bai, S.B. Impacts of moso bamboo invasion on carbon storage in broad-leaved evergreen forests. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2020, 49, 809–815. [Google Scholar]
- Fahad, M.S.; Zhihao, C.; Caixian, T.; Zhang, S.; Fang, L.; Miao, D.N.; Li, Y.F.; Zhang, Q.Q.; Li, Y.C. Seasonal linkages between soil nitrogen mineralization and the microbial community in broadleaf forests with Moso bamboo (Phyllostachys edulis) invasion. Sci. Total Environ. 2023, 899, 165557. [Google Scholar]
- Li, Y.M.; Feng, P.F. Analysis of bamboo resources in China based on the ninth national forest inventory. World Bamboo Ratt. Newsl. 2019, 17, 45–48. [Google Scholar]
- Liu, C.X.; Zheng, C.Y.; Wang, L.; Zhang, J.; Wang, Q.Z.; Shao, S.; Qin, H.; Xu, Q.F.; Liang, C.F.; Chen, J.H. Moso bamboo invasion changes the assembly process and interactive relationship of soil microbial communities in a subtropical broadleaf forest. For. Ecol. Manag. 2023, 536, 120901. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakagoshi, N. Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions. Ecol. Res. 2008, 23, 641–647. [Google Scholar] [CrossRef]
- Isagi, Y.; Torii, A. Range Expansion and Its Mechanisms in a Naturalized Bamboo Species, Phyllostachys pubescens, in Japan. J. Sustain. For. 1997, 6, 127–141. [Google Scholar] [CrossRef]
- Zhou, G.M.; Jiang, P.K. Carbon density and carbon storage of moso bamboo forest and its spatial distribution. For. Sci. 2004, 40, 20–24. [Google Scholar]
- Wang, X.Y.; Song, X.Z.; Pang, C.M.; Yang, H.; Zhao, M.S.; Xu, L. Influence of stand structure and stand establishment of moso bamboo neighboring forests on their natural expansion. Zhejiang For. Sci. Technol. 2019, 39, 19–24. [Google Scholar]
- Song, Q.N.; Ouyang, M.; Yang, Q.P.; Lu, H.; Yang, G.Y.; Chen, F.S.; Shi, J.M. Degradation of litter quality and decline of soil nitrogenmineralization after moso bamboo (Phyllostachys pubscens) expansion to neighboring broadleavedforest in subtropical China. Plant Soil 2016, 404, 113–124. [Google Scholar] [CrossRef]
- Durga, B.D.; Hiromasa, H. Expansion Characteristics of Bamboo Stand and Sediment Disaster in South Western Japan. Pak. J. Biol. Sci. 2006, 9, 622–631. [Google Scholar]
- Tong, R.; Zhou, B.Z.; Jiang, L.N.; Cao, Y.H.; Ge, X.G.; Yang, Z.Y. Progress of research on the impact of moso bamboo invasion on forest plants and soil. J. Ecol. 2019, 39, 3808–3815. [Google Scholar]
- He, D.H.; Shen, Q.L.; Xu, Q.F.; Chen, J.H.; Cheng, M.; Mao, X.W.; Li, Y.C. Evolution of community structure and abundance of soil nitrogen-fixing bacteria in moso bamboo forests of different ages. Soil J. 2015, 52, 934–942. [Google Scholar]
- Ouyang, M.; Yang, Q.; Chen, X.; Yang, G.Y.; Shi, J.M.; Fang, X.M. Effects of the expansion of Phyllostachys edulis on species composition, structure and diversity of the secondary evergreen broad-leaved forests. Biodivers. Sci. 2016, 24, 649–657. [Google Scholar] [CrossRef]
- Deyn, G.B.; Raaijmakers, C.E.; Ruijven, J.; Berendse, F.; Putten, V.D. Plant species identity and diversity effects on different trophic levels of nematodes in the soil food web. Oikos 2004, 106, 576–586. [Google Scholar] [CrossRef]
- Zeng, J.Y.; Vuong, T.M.D.; Zhang, B.W.; Chen, Y.Z.; Zhang, G.C.; Bi, B. An investigation into whether effect of tree species on soil microbial community is related with deciduous property or leaf shape. Catena 2020, 195, 104699. [Google Scholar] [CrossRef]
- Na, Z.; Weiming, S.; Lihan, H.; Kronzucker, H.J.; Huang, L.; Gu, H.M.; Yang, Q.P.; Deng, G.H.; Yang, G.Y. Superior growth, N uptake and NH4+ tolerance in the giant bamboo Phyllostachys edulis over the broad-leaved tree Castanopsis fargesii at elevated NH4+ may under liecommunity succession and favor the expansion of bamboo. Tree Physiol. 2020, 40, 1606–1622. [Google Scholar]
- Chang, E.; Chiu, C. Changes in soil microbial community structure and activity in a cedar plantation invaded by moso bamboo. Appl. Soil Ecol. 2015, 91, 1–7. [Google Scholar] [CrossRef]
- Wu, J.S.; Jiang, P.K.; Zu, L. Impact of expansion of moso bamboo on soil fertility of forest land in Tianmu Mountain National Nature Reserve. J. Jiangxi Agric. Univ. 2008, 30, 689–692. [Google Scholar]
- Wang, N.N.; Yang, X.; Li, S.L.; Sui, X.; Han, S.J.; Feng, F.J. Distribution patterns of soil fungal diversity in red pine broadleaf forests driven by precipitation changes. J. Appl. Ecol. 2013, 24, 1985–1990. [Google Scholar]
- Xu, Q.F.; Jiang, P.K.; Wu, J.S.; Zhou, G.M.; Shen, R.F.; Fuhrmann, J.J. Bamboo invasion of native broadleaf forest modified soil microbial communities and diversity. Biol. Invasions 2015, 17, 433–444. [Google Scholar] [CrossRef]
- Ming, J.Z. The relationship between soil pH and geochemical components. Environ. Earth Sci. 2024, 83, 402. [Google Scholar]
- He, H.J.; Zhang, Z.H.; Zhang, C.Y.; Hao, M.H.; Yao, J.; Xie, Z.; Gao, H.T.; Zhao, X.H. Short-term effects of cutting intensity on stand growth and species diversity of mixed needle and broad forest in northeast China. For. Sci. 2019, 55, 1–12. [Google Scholar]
- Tu, J.; Yang, X.B.; Wei, B.Y.; Zhao, X.; Wei, L.; Fu, W.; Yang, Y.R.; Wang, H.Y.; Yi, Y.J.; Zeng, L.J. Effects of Bacillus subtilis Y25 on the growth of Longya lily and its inter-root soil microbial community. Jiangsu Agric. Sci. 2024, 52, 214–223. [Google Scholar]
- GB/T 1215-1999; Determination of Forest Soil Water. Physical Properties. China Academy of Forestry, Forest Research Institute: Beijing, China, 1999.
- Shi, Z.Y.; Wang, Y.; Liu, Y.Q. Study on the measurement method of carbon, hydrogen and nitrogen in coal sample by element analyzer. Coal Process. Compr. Util. 2024, 66–70. [Google Scholar]
- Gu, X.J.; Yu, Y.H.; Liu, Y.; Wu, M.J.; Liu, L.Y.; Mo, Q.F. Effects of typical plantations on soil readily oxidized organic carbon in South China. J. For. Environ. 2023, 43, 145–151. [Google Scholar]
- Huang, Y.M.; An, S.S.; Qu, D.; Li, M.J. Comparison of two methods for determination of soil microbial biomass nitrogen. Plant Nutr. Fertil. J. 2005, 11, 830–835. [Google Scholar]
- Liu, C.; Zhao, D.; Ma, W.; Guo, Y.; Wang, A.; Wang, Q.; Lee, D.J. Denitrifying sulfide removal process on high-salinity wastewaters in the presence of Halomonas sp. Appl. Microbiol. Biotechnol. 2016, 100, 1421–1426. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537. [Google Scholar] [CrossRef]
- Wang, Z.; Bi, Y.; Lu, G.; Zhang, X.; Xu, X.Y.; Ning, Y.L.; Du, X.H.; Wang, A.K. Monitoring Forest Diversity under Moso Bamboo Invasion: A Random Forest Approach. Forests 2024, 15, 318. [Google Scholar] [CrossRef]
- Yang, H.Q.; Xiang, Y.Q.; Lv, Q.; Yin, B.R.; Tang, Z.R.; Zhang, Y.; Chen, G.; Lai, J.M.; Fan, C.; Li, X.W. Study on soil fungal community structure at the early stage of afforestation of three mixed forests. J. Ecol. 2024, 44, 1–12. [Google Scholar]
- Qi, C.; Qin, Z.; Yuanqiu, L.; Lan, L.Y.; Gao, Y.Y.; Yuan, X.; Huang, J.H.; Fu, X.B.; Li, L.H.; Deng, W.P. Analysis on the transpiration response of Japanese cedar (Crytomeria fortunei) and influencing factors after expansion of moso bamboo (Phyllostachys edulis). Ecol. Indic. 2023, 154, 110636. [Google Scholar]
- Mo, L.P.; Liang, Z.M. Characteristics of apomorphy and soil water holding in typical coniferous and broad-leaved forests in Guidong. Mod. Agric. Sci. Technol. 2023, 85–89. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, C.L.; Zhao, H.X.; Sun, X.J.; Peng, G.W.; Zhang, J.; Zhao, W.L.; Xia, J.B. Characteristics of photosynthetic physiological parameters of hickory leaves in response to soil moisture. Shandong For. Sci. Technol. 2023, 53, 28–35+40. [Google Scholar]
- Ou, Y.M.; Yang, Q.P.; Chen, X.; Yang, G.Y.; Shi, J.M.; Fang, X.M. Effects of moso bamboo expansion on species composition, structure and diversity of secondary broad-leaved evergreen forest. Biodiversity 2016, 24, 649–657. [Google Scholar]
- Bai, S.B.; Zhou, G.M.; Wang, Y.X.; Yu, S.Q.; Li, Y.H.; Fang, F.Y. Study on the change of stand structure during the dispersal of moso bamboo in Tianmu Mountain National Nature Reserve. West. For. Sci. 2012, 41, 77–82. [Google Scholar]
- Lv, H.; Zhang, J.; Yi, K.X.; Huang, Z.Y.; Zhang, X.P.; Zhong, Z.K.; Yang, C.B. Effects of different management measures on soil physicochemical properties, enzyme activities and organic carbon mineralization characteristics of moso bamboo forests. Bamboo J. 2022, 41, 34–40. [Google Scholar]
- Shinohara, Y.; Otsuki, K. Comparisons of soil-water content between a moso bamboo (Phyllostachys pubescens) forest and an evergreen broadleaved forest in western Japan. Plant Species Biol. 2015, 30, 96–103. [Google Scholar] [CrossRef]
- Cui, C.; Zhang, J.T.; Guo, X.M. Effects of moso bamboo invasion of broadleaf forest on soil physical properties. South. For. Sci. 2020, 48, 1–6. [Google Scholar]
- Zhao, Y.H.; Fan, S.H.; Luo, J.D. Effects of moso bamboo expansion on soil properties of broad-leaved evergreen forest and related analysis. For. Sci. Res. 2017, 30, 354–359. [Google Scholar]
- Bai, S.B.; Zhou, G.M.; Wang, Y.X.; Liang, Q.Q.; Chen, J.; Cheng, Y.Y.; Shen, R. Response and dynamics of forest community plant diversity to moso bamboo invasion in Tianmu Mountain Reserve. Biodiversity 2013, 21, 288–295. [Google Scholar]
- Umemura, M.; Takenaka, C. Changes in chemical characteristics of surface soils in hinoki cypress (Chamaecyparis obtusa) forests induced by the invasion of exotic moso bamboo (Phyllostachys pubescens) in central Japan. Plant Species Biol. 2015, 30, 72–79. [Google Scholar] [CrossRef]
- Xu, D.W. Effects of Expansion of Moso Bamboo to Cedar in Daiyunshan Nature Reserve on Soil Quality of Forest Stands and Its Apomictic Decomposition. Ph.D. Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2018. [Google Scholar]
- Zhang, D.L.; Zhang, L. Soil soluble organic carbon content and its influencing factors in different succession series of broad-leaved red pine forest. For. Eng. 2024, 40, 10–16. [Google Scholar]
- Zhang, Y.; Wu, F.Z.; Ai, L.; Fan, X.B.; Yang, Y.; Zhen, X.P.; Zhu, J.J.; Ni, X.Y. Effects of litter input on soil soluble organic carbon. J. Ecol. 2023, 42, 2107–2112. [Google Scholar]
- Li, W.C.; Sheng, H.Y.; Chen, W.J.; Liu, Y.Y.; Zhang, R.; Wen, X. Changes in soil flora diversity of moso bamboo invading Malus domestica forest. J. Appl. Ecol. 2018, 29, 3969–3976. [Google Scholar]
- Ma, Y.; Ji, X.L.; Zou, B.Y.; Xue, Y.Y.; Wang, Y.K. Effects of expansion of moso bamboo forest on soil biological community structure. Anhui Agric. Bull. 2023, 29, 64–70. [Google Scholar]
- Zhao, Y.Z.; Liang, C.F.; Shao, S.; Chen, J.H.; Qin, H. Linkages of litter and soil C:N:P stoichiometry with soil microbial resource limitation and community structure in a subtropical broadleaf forest invaded by Moso bamboo. Plant Soil 2021, 465, 473–490. [Google Scholar] [CrossRef]
- Xu, Y.M. Response of soil microbial biomass carbon to simulated nitrogen deposition in poplar plantation forest. Jiangsu For. Sci. Technol. 2023, 50, 1–7. [Google Scholar]
- Zhao, L.J.; Chen, S.J.; Luo, L.F.; Zeng, X.L.; Zhang, H.S.; Lin, R. Progress of soil microbiota research in Yunnan Province. J. Intell. Agric. 2024, 4, 37–40. [Google Scholar]
- Ma, X.R.; Zheng, X.L.; Zheng, C.Y.; Hu, Y.T.; Qin, H.; Chen, J.H.; Xu, Q.F.; Liang, C.F. Effects of moso bamboo expansion on soil microbial communities in broad-leaved evergreen forests. J. Appl. Ecol. 2022, 33, 1091–1098. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strip | Sample Plot | Slope (°) | DOS | SP | SD (Plants/hm2) | ATH (m) | ACD (cm) | CD | Species Diversity Index * |
|---|---|---|---|---|---|---|---|---|---|
| strip 1 | Phyllostachys edulis forest | 15 | east | middle | 3600 | 10.11 | 9.33 | 0.9 | 0.88 |
| Mixed forest | 15 | east | middle | 2200 | 13.28 | 16.73 | 0.8 | 1.76 | |
| Quercus acutissima forest | 15 | east | middle | 1000 | 13.77 | 19.32 | 0.8 | 2.02 | |
| strip 2 | Phyllostachys edulis forest | 18 | east | middle | 4400 | 10.83 | 10.10 | 0.9 | 1.73 |
| Mixed forest | 18 | east | middle | 2600 | 13.18 | 18.93 | 0.8 | 1.91 | |
| Quercus acutissima forest | 18 | east | middle | 1600 | 10.85 | 19.67 | 0.8 | 2.19 | |
| strip 3 | Phyllostachys edulis forest | 17 | east | middle | 3500 | 11.01 | 10.17 | 0.9 | 1.41 |
| Mixed forest | 17 | east | middle | 1800 | 13.73 | 17.22 | 0.8 | 1.79 | |
| Quercus acutissima forest | 17 | east | middle | 1100 | 11.96 | 19.03 | 0.8 | 1.93 |
| Stand Type | Soil Depth (cm) | Soil Bulk Density (g/cm3) | Soil Water Content (g/kg) | Capillary Water Capacity (g/kg) |
|---|---|---|---|---|
| Phyllostachys edulis forest | 0–10 | 1.10 ± 0.03 Bb | 289.46 ± 10.08 Aa | 432.78 ± 12.89 Aa |
| 10–20 | 1.27 ± 0.08 Aa | 255.04 ± 14.17 Ba | 357.22 ± 10.11 Ba | |
| 20–40 | 1.35 ± 0.12 Aa | 220.86 ± 10.25 Ca | 328.96 ± 15.23 Ca | |
| Mixed forest | 0–10 | 1.16 ± 0.02 Ba | 265.61 ± 9.45 Ab | 402.27 ± 10.64 Ab |
| 10–20 | 1.29 ± 0.14 ABa | 229.11 ± 10.36 Bb | 324.28 ± 17.48 Bb | |
| 20–40 | 1.37 ± 0.07 Aa | 197.94 ± 10.46 Cb | 287.66 ± 18.14 Cb | |
| Quercus acutissima forest | 0–10 | 1.18 ± 0.04 Ba | 243.83 ± 11.44 Ac | 371.89 ± 12.27 Ac |
| 10–20 | 1.29 ± 0.09 ABa | 210.53 ± 7.73 Bc | 266.88 ± 15.33 Bc | |
| 20–40 | 1.38 ± 0.10 Aa | 173.55 ± 10.27 Cc | 226.29 ± 17.54 Cc |
| Stand Type | Soil Depth (cm) | Capillary Porosity (%) | Noncapillary Porosity (%) | Total Porosity (%) |
|---|---|---|---|---|
| Phyllostachys edulis forest | 0–10 | 47.70 ± 0.83 Aa | 5.65 ± 0.11 Aa | 53.34 ± 0.19 Aa |
| 10–20 | 45.67 ± 0.76 Ba | 3.55 ± 0.57 Ba | 49.22 ± 1.89 Ba | |
| 20–40 | 43.34 ± 1.77 Ba | 2.92 ± 0.14 Ba | 46.26 ± 2.26 Ba | |
| Mixed forest | 0–10 | 43.00 ± 0.54 Ab | 5.02 ± 0.41 Ab | 48.02 ± 1.41 Ab |
| 10–20 | 40.91 ± 0.80 Bb | 4.04 ± 0.36 Ba | 44.95 ± 0.40 Bb | |
| 20–40 | 40.62 ± 0.26 Bb | 3.49 ± 0.33 Ba | 44.11 ± 0.86 Bab | |
| Quercus acutissima forest | 0–10 | 42.69 ± 0.46 Ab | 3.87 ± 0.66 Ab | 46.56 ± 1.59 Ab |
| 10–20 | 39.61 ± 0.19 Bc | 3.80 ± 0.32 Aa | 43.41 ± 1.21 Bbc | |
| 20–40 | 38.71 ± 0.87 Bc | 2.39 ± 0.32 Bb | 41.1 ± 1.98 Bb |
| Stand Type | Soil Depth (cm) | pH | SOC (g/kg) | ROC(g/kg) |
|---|---|---|---|---|
| Phyllostachys edulis forest | 0–10 | 4.56 ± 0.11 Aa | 17.25 ± 0.17 Ab | 0.96 ± 0.10 Aa |
| 10–20 | 4.44 ± 0.12 Aa | 9.04 ± 0.78 Bb | 0.85 ± 0.06 Ba | |
| 20–40 | 4.42 ± 0.08 Aa | 6.57 ± 0.30 Cb | 0.84 ± 0.07 Ba | |
| Mixed forest | 0–10 | 4.38 ± 0.06 Ab | 18.34 ± 0.63 Aa | 0.86 ± 0.08 Ac |
| 10–20 | 4.24 ± 0.05 Ab | 11.00 ± 0.25 Aa | 0.78 ± 0.06 Bb | |
| 20–40 | 4.23 ± 0.09 Ab | 8.80 ± 1.00 Ba | 0.75 ± 0.08 Bb | |
| Quercus acutissima forest | 0–10 | 4.37 ± 0.06 Ab | 20.43 ± 1.70 Aa | 0.92 ± 0.09 Ab |
| 10–20 | 4.22 ± 0.08 Ab | 13.42 ± 2.21 Ba | 0.88 ± 0.15 Ba | |
| 20–40 | 4.21 ± 0.11 Ab | 9.63 ± 1.24 Ba | 0.86 ± 0.12 Ba |
| Stand Type | Chao Index | Shannon Index | Simpson Index |
|---|---|---|---|
| Phyllostachys edulis forest | 2411.00 ± 171.52 a | 6.82 ± 0.19 a | 0.0028 ± 0.0008 a |
| Mixed forest | 2076.04 ± 103.49 b | 6.47 ± 0.12 b | 0.0026 ± 0.0006 a |
| Quercus acutissima forest | 2095.63 ± 135.04 b | 6.47 ± 0.15 b | 0.0028 ± 0.0008 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, L.; Hu, H.; Chen, J.; Gong, Y.; Zhu, Z. Mechanism of the Effects of Phyllostachys edulis Invasion on the Soil Microbial Community in Quercus acutissima Forests. Forests 2024, 15, 1170. https://doi.org/10.3390/f15071170
Fang L, Hu H, Chen J, Gong Y, Zhu Z. Mechanism of the Effects of Phyllostachys edulis Invasion on the Soil Microbial Community in Quercus acutissima Forests. Forests. 2024; 15(7):1170. https://doi.org/10.3390/f15071170
Chicago/Turabian StyleFang, Lu, Haibo Hu, Jianyu Chen, Yuyang Gong, and Ziyi Zhu. 2024. "Mechanism of the Effects of Phyllostachys edulis Invasion on the Soil Microbial Community in Quercus acutissima Forests" Forests 15, no. 7: 1170. https://doi.org/10.3390/f15071170
APA StyleFang, L., Hu, H., Chen, J., Gong, Y., & Zhu, Z. (2024). Mechanism of the Effects of Phyllostachys edulis Invasion on the Soil Microbial Community in Quercus acutissima Forests. Forests, 15(7), 1170. https://doi.org/10.3390/f15071170