

Biomass and Carbon Stock Capacity of Robinia pseudoacacia Plantations at Different Densities on the Loess Plateau


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
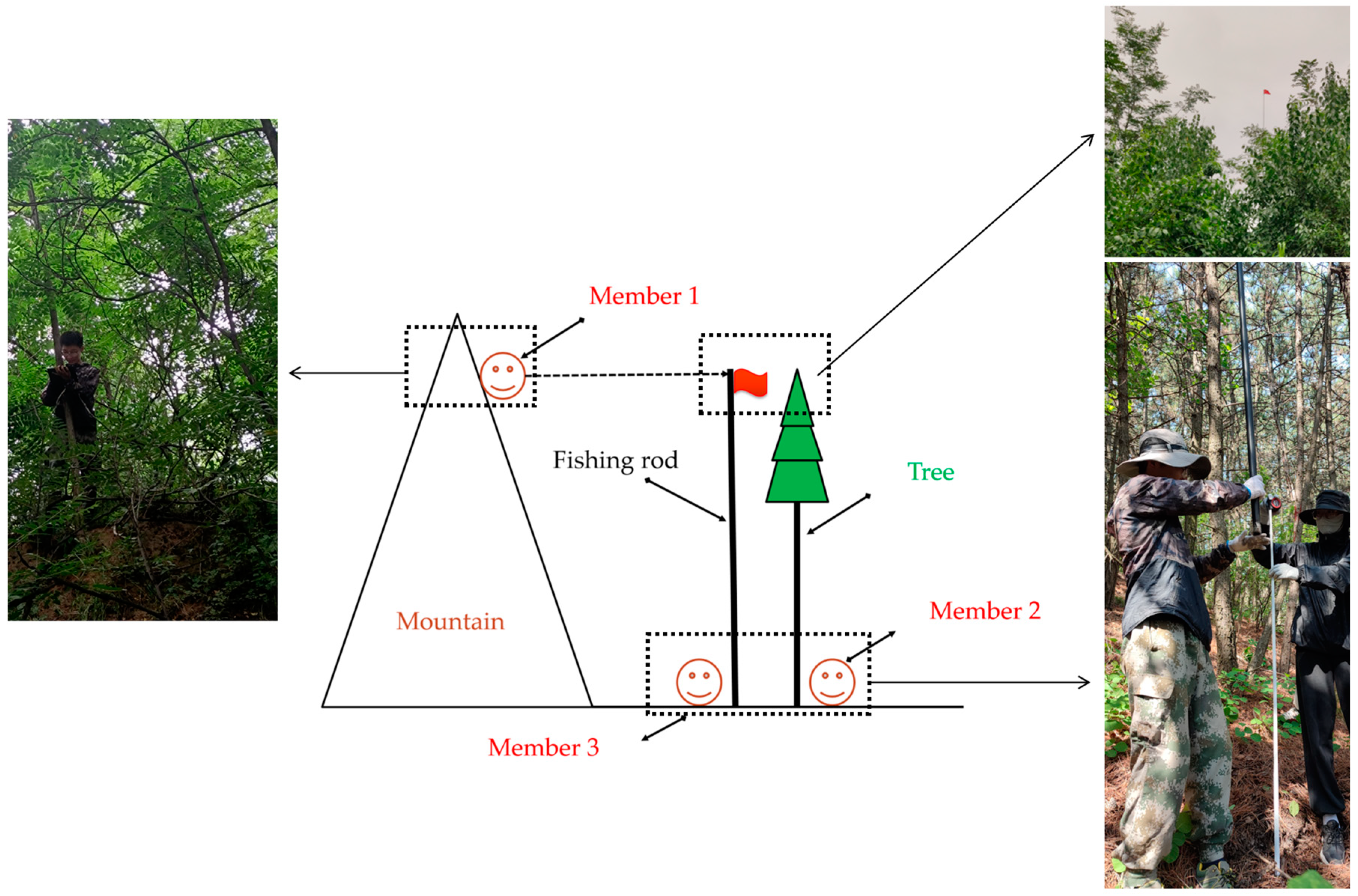
2.2. Selection of Experimental Sites
2.3. Methods for Estimating Aboveground Biomass
2.4. Methods for Estimating Belowground Biomass
2.5. Fitting of Biomass Relationships
2.6. Soil Sample Collection and Determination
2.7. Carbon Stocks Estimation
2.8. Statistical Analysis
3. Results
3.1. Fitting of Biomass Relationships
3.2. Biomass and Contributions in R. pseudoacacia of Different Densities
The Biomass of Individual Trees of R. pseudoacacia
3.3. Carbon Stock in R. pseudoacacia of Different Densities
4. Discussion
4.1. Comparison of Biomass Models
4.2. Biomass Production
4.3. Total Carbon Stocks
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luintel, H.; Scheller, R.M.; Bluffstone, R.A. Assessments of Biodiversity, Carbon, and Their Relationships in Nepalese Forest Commons: Implications for Global Climate Initiatives. For. Sci. 2018, 64, 418–428. [Google Scholar] [CrossRef]
- IPCC. 2023: Sections. In Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; pp. 35–115. [Google Scholar] [CrossRef]
- Cao, L.; Kong, F.; Xu, C. Exploring Ecosystem Carbon Storage Change and Scenario Simulation in the Qiantang River Source Region of China. Sci. Prog. 2022, 105, 003685042211131. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Chen, X. Land Use Transitions and the Associated Impacts on Carbon Storage in the Poyang Lake Basin, China. Remote Sens. 2023, 15, 2703. [Google Scholar] [CrossRef]
- Zhou, Y.; Williams, C.A.; Hasler, N.; Gu, H.; Kennedy, R. Beyond biomass to carbon fluxes: Application and evaluation of a comprehensive forest carbon monitoring system. Environ. Res. Lett. 2021, 16, 055026. [Google Scholar] [CrossRef]
- Carvalhais, N.; Forkel, M.; Khomik, M. Global covariation of carbon turnover times with climate in terrestrial ecosystems. Nature 2014, 514, 213–217. [Google Scholar] [CrossRef]
- Kozlowski, G.; Song, Y.G. Importance, Tools, and Challenges of Protecting Trees. Sustainability 2022, 14, 13107. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Gauthier, S. Young and Old Forest in the Boreal: Critical Stages of Ecosystem Dynamics and Management under Global Change. For. Ecosyst. 2018, 5, 26. [Google Scholar] [CrossRef]
- Węgiel, A.; Polowy, K. Aboveground Carbon Content and Storage in Mature Scots Pine Stands of Different Densities. Forests 2020, 11, 240. [Google Scholar] [CrossRef]
- Poorter, L.; Van Der Sande, M.T.; Arets, E.J.M.M.; Ascarrunz, N.; Enquist, B.J.; Finegan, B.; Licona, J.C.; Martínez-Ramos, M.; Mazzei, L.; Meave, J.A.; et al. Biodiversity and Climate Determine the Functioning of Neotropical Forests. Glob. Ecol. Biogeogr. 2017, 26, 1423–1434. [Google Scholar] [CrossRef]
- Zeng, W.-S. Development of Monitoring and Assessment of Forest Biomass and Carbon Storage in China. For. Ecosyst. 2014, 1, 20. [Google Scholar] [CrossRef]
- Zhang, C.; Ju, W.; Chen, J.M.; Wang, X.; Yang, L.; Zheng, G. Disturbance-Induced Reduction of Biomass Carbon Sinks of China’s Forests in Recent Years. Environ. Res. Lett. 2015, 10, 114021. [Google Scholar] [CrossRef]
- Fang, J.; Yu, G.; Liu, L.; Hu, S.; Chapin, F.S. Climate Change, Human Impacts, and Carbon Sequestration in China. Proc. Natl. Acad. Sci. USA 2018, 115, 4015–4020. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yoon, T.K.; Han, S.; Kim, S.; Yi, M.J.; Park, G.S.; Kim, C.; Son, Y.M.; Kim, R.; Son, Y. Estimating the Carbon Dynamics of South Korean Forests from 1954 to 2012. Biogeosciences 2014, 11, 4637–4650. [Google Scholar] [CrossRef]
- Hérault, B.; Piponiot, C. Key Drivers of Ecosystem Recovery after Disturbance in a Neotropical Forest: Long-Term Lessons from the Paracou Experiment, French Guiana. For. Ecosyst. 2018, 5, 2. [Google Scholar] [CrossRef]
- Lafleur, B.; Fenton, N.J.; Simard, M.; Leduc, A.; Paré, D.; Valeria, O.; Bergeron, Y. Ecosystem Management in Paludified Boreal Forests: Enhancing Wood Production, Biodiversity, and Carbon Sequestration at the Landscape Level. For. Ecosyst. 2018, 5, 27. [Google Scholar] [CrossRef]
- Tian, H.; Lu, C.; Yang, J.; Banger, K.; Huntzinger, D.N.; Schwalm, C.R.; Michalak, A.M.; Cook, R.; Ciais, P.; Hayes, D.; et al. Global Patterns and Controls of Soil Organic Carbon Dynamics as Simulated by Multiple Terrestrial Biosphere Models: Current Status and Future Directions. Glob. Biogeochem. Cycles 2015, 29, 775–792. [Google Scholar] [CrossRef] [PubMed]
- Vanguelova, E.I.; Bonifacio, E.; De Vos, B.; Hoosbeek, M.R.; Berger, T.W.; Vesterdal, L.; Armolaitis, K.; Celi, L.; Dinca, L.; Kjønaas, O.J.; et al. Sources of Errors and Uncertainties in the Assessment of Forest Soil Carbon Stocks at Different Scales—Review and Recommendations. Environ. Monit. Assess. 2016, 188, 630. [Google Scholar] [CrossRef] [PubMed]
- Domke, G.M.; Perry, C.H.; Walters, B.F.; Woodall, C.W.; Russell, M.B.; Smith, J.E. Estimating Litter Carbon Stocks on Forest Land in the United States. Sci. Total Environ. 2016, 557–558, 469–478. [Google Scholar] [CrossRef]
- Filipchuk, A.; Moiseev, B.; Malysheva, N.; Strakhov, V. Russian Forests: A New Approach to the Assessment of Carbon Stocks and Sequestration Capacity. Environ. Dev. 2018, 26, 68–75. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, J.; Zhou, L.; Li, F.; Fu, S. Effects of Prescribed Burning on Carbon Accumulation in Two Paired Vegetation Sites in Subtropical China. For. Ecosyst. 2019, 6, 26. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Wang, Y.; Yu, P.; Guo, J. Impacts of Site Conditions and Stand Structure on the Biomass Allocation of Single Trees in Larch Plantations of Liupan Mountains of Northwest China. Forests 2022, 13, 177. [Google Scholar] [CrossRef]
- Le Toan, T.; Quegan, S.; Davidson, M.W.J.; Balzter, H.; Paillou, P.; Papathanassiou, K.; Plummer, S.; Rocca, F.; Saatchi, S.; Shugart, H.; et al. The BIOMASS Mission: Mapping Global Forest Biomass to Better Understand the Terrestrial Carbon Cycle. Remote Sens. Environ. 2011, 115, 2850–2860. [Google Scholar] [CrossRef]
- Adhikari, D.; Singh, P.P.; Tiwary, R.; Barik, S.K. Forest carbon stock-based bioeconomy: Mixed models improve accuracy of tree biomass estimates. Biomass Bioenergy 2024, 183, 107142. [Google Scholar] [CrossRef]
- Disney, M.I.; Boni Vicari, M.; Burt, A.; Calders, K.; Lewis, S.L.; Raumonen, P.; Wilkes, P. Weighing Trees with Lasers: Advances, Challenges and Opportunities. Interface Focus 2018, 8, 20170048. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.L.; Cao, M.K.; Li, K.R. Temporal-spatial dynamics of carbon storage of forest vegetation in China. Prog. Geogr. 2007, 26, 1–10. [Google Scholar]
- Sun, W.; Liu, X. Review on Carbon Storage Estimation of Forest Ecosystem and Applications in China. For. Ecosyst. 2020, 7, 4. [Google Scholar] [CrossRef]
- Pfeiffer, B.; Fender, A.-C.; Lasota, S.; Hertel, D.; Jungkunst, H.F.; Daniel, R. Leaf Litter Is the Main Driver for Changes in Bacterial Community Structures in the Rhizosphere of Ash and Beech. Appl. Soil Ecol. 2013, 72, 150–160. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Ruiz-Peinado, R.; Onrubia, R.; Del Río, M. Thinning Alters the Early-Decomposition Rate and Nutrient Immobilization-Release Pattern of Foliar Litter in Mediterranean Oak-Pine Mixed Stands. For. Ecol. Manag. 2017, 391, 309–320. [Google Scholar] [CrossRef]
- Pretzsch, H. Density and Growth of Forest Stands Revisited. Effect of the Temporal Scale of Observation, Site Quality, and Thinning. For. Ecol. Manag. 2020, 460, 117879. [Google Scholar] [CrossRef]
- Hepner, H.; Lutter, R.; Tullus, A.; Kanal, A.; Tullus, T.; Tullus, H. Effect of Early Thinning Treatments on Above-Ground Growth, Biomass Production, Leaf Area Index and Leaf Growth Efficiency in a Hybrid Aspen Coppice Stand. BioEnergy Res. 2020, 13, 197–209. [Google Scholar] [CrossRef]
- Zhao, M.; Sun, Y.; Liu, S.; Li, Y.; Chen, Y. Effects of Stand Density on the Structure of Soil Microbial Functional Groups in Robinia pseudoacacia Plantations in the Hilly and Gully Region of the Loess Plateau, China. Sci. Total Environ. 2024, 912, 169337. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Du, H.; Duan, A.; Zhang, J. Effect of Stand Density and Soil Layer on Soil Nutrients of a 37-Year-Old Cunninghamia lanceolata Plantation in Naxi, Sichuan Province, China. Sustainability 2019, 11, 5410. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, J.; Hu, Y.; Li, Y.; Tang, P.; Gusarov, A.V.; Yu, Y. Effects of Land Uses and Rainfall Regimes on Surface Runoff and Sediment Yield in a Nested Watershed of the Loess Plateau, China. J. Hydrol. Reg. Stud. 2022, 44, 101277. [Google Scholar] [CrossRef]
- Jurjević, L.; Liang, X.; Gašparović, M.; Balenović, I. Is Field-Measured Tree Height as Reliable as Believed—Part II, A Comparison Study of Tree Height Estimates from Conventional Field Measurement and Low-Cost Close-Range Remote Sensing in a Deciduous Forest. ISPRS J. Photogramm. Remote Sens. 2020, 169, 227–241. [Google Scholar] [CrossRef]
- Byambadorj, S.O.; Park, B.B.; Lkhagvasuren, S.; Tsedensodnom, E.; Byambasuren, Q.; Khajid, A.; Chiatate, D.; Nyam-Osor, B. Influence of Irrigation on Biomass Partitioning in Above-and Belowground Organs of Trees Planted in Desert Sites of Mongolia. Forests 2023, 15, 46. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; He, S. Effect of Vegetation Growth on Morphological Traits of Vegetation and Biomechanical Features of Roots. Plant Soil 2023, 494, 395–411. [Google Scholar] [CrossRef]
- Cabal, C.; Rodríguez-Torres, L.; Marí-Mena, N.; Más-Barreiro, A.; Vizcaíno, A.; Vierna, J.; Valladares, F.; Pacala, S.W. Comparing Two Field Protocols to Measure Individual Shrubs’ Root Density Distribution. Plant Soil 2022, 481, 691–699. [Google Scholar] [CrossRef]
- Li, T.; Wu, X.-C.; Wu, Y.; Li, M.-Y. Forest Carbon Density Estimation Using Tree Species Diversity and Stand Spatial Structure Indices. Forests 2023, 14, 1105. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Zhang, W.; Chen, B.; Yang, L.; Li, M.; Zhu, J.; Cai, Q. The Effects of Thinning on Tree Growth and Stand Biomass in a Chronosequence of Pinus tabulaeformis Plantations in the Loess Plateau of China. Forests 2023, 14, 1620. [Google Scholar] [CrossRef]
- Hu, Y.W.; Shi, Z.L.; Liu, C.; Xu, Q.T.; Zhang, J.J. Effects of stand densities on understory vegetation diversity and soil physicochemical properties of Robinia pseudoacacia forest in loess region of western Shanxi Province. Chin. J. Ecol. 2023, 42, 2072–2080. [Google Scholar]
- Sun, L.N. Study on Spatial Pattern and Driving Factors of Forests Biomass Carbon Density in Shanxi Province, China. Ph.D. Thesis, Shanxi University, Taiyuan, China, 2020. [Google Scholar]
- Yadav, R.P.; Gupta, B.; Bhutia, P.L.; Bisht, J.K.; Pattanayak, A. Biomass and Carbon Budgeting of Land Use Types along Elevation Gradient in Central Himalayas. J. Clean. Prod. 2019, 211, 1284–1298. [Google Scholar] [CrossRef]
- Kralicek, K.; Huy, B.; Poudel, K.P.; Temesgen, H.; Salas, C. Simultaneous Estimation of Above- and Below-Ground Biomass in Tropical Forests of Viet Nam. For. Ecol. Manag. 2017, 390, 147–156. [Google Scholar] [CrossRef]
- Bolat, İ. The Effect of Thinning on Microbial Biomass C, N and Basal Respiration in Black Pine Forest Soils in Mudurnu, Turkey. Eur. J. For. Res. 2014, 133, 131–139. [Google Scholar] [CrossRef]
- Xue, L.; Pan, L.; Zhang, R.; Xu, P. Density Effects on the Growth of Self-Thinning Eucalyptus urophylla Stands. Trees 2011, 25, 1021–1031. [Google Scholar] [CrossRef]
- Dai, L.L.; Zhou, L.L.; Wu, L.H.; Liu, L.; Huang, Y.; Peng, T.T.; Qiu, J.; He, Z.M.; Cao, G.M. Carbon density and vertical spatial distribution characteristics of Cunninghamia lanceolata forest ecosystem with different stand density. Acta Ecol. Sin. 2022, 42, 710–719. [Google Scholar] [CrossRef]
- Lado-Monserrat, L.; Lidón, A.; Bautista, I. Erratum to: Litterfall, Litter Decomposition and Associated Nutrient Fluxes in Pinus halepensis: Influence of Tree Removal Intensity in a Mediterranean Forest. Eur. J. For. Res. 2016, 135, 203–214. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, S.; Sun, Y.; Chen, Y. Does Stand Density Affect Understory Vegetation and Soil Properties of Differently Aged Robinia pseudoacacia Plantations? For. Ecol. Manag. 2023, 548, 121444. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Wang, Y.H.; Tian, A.; Liu, Z.B.; Guo, J.B.; Yu, P.T.; Wang, X.; Yu, Y.P. Spatiotemporal characteristics and environmental response of vegetation carbon densities of Larix principis-rupprechtii plantations in the Liupan mountains of Ningxia, China. Sci. Silv. Sin. 2023, 59, 32–45. [Google Scholar]
- Li, C.; Zha, T.; Liu, J.; Jia, X. Carbon and Nitrogen Distribution across a Chronosequence of Secondary Lacebark Pine in China. For. Chron. 2013, 89, 192–198. [Google Scholar] [CrossRef]
- Kim, C.; Jeong, J.; Cho, H.-S.; Son, Y. Carbon and Nitrogen Status of Litterfall, Litter Decomposition and Soil in Even-Aged Larch, Red Pine and Rigitaeda Pine Plantations. J. Plant Res. 2010, 123, 403–409. [Google Scholar] [CrossRef]
- Liu, S. Storage and Key Soil Cycling Processes of Carbon and Nitrogen among Typical Forest Ecosystems in the Subalpine Area of Western Sichuan. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2019. [Google Scholar]
- Xu, S.; Su, F.; Sayer, E.J.; Lam, S.K.; Lu, X.; Liu, C.; Lai, D.Y.F. Fine Root Litter Quality Regulates Soil Carbon Storage Efficiency in Subtropical Forest Soils. Soil Ecol. Lett. 2023, 5, 230182. [Google Scholar] [CrossRef]
- Zhao, M.; Li, Y.; Wang, Y.; Sun, Y.; Chen, Y. High Stand Density Promotes Soil Organic Carbon Sequestration in Robinia pseudoacacia Plantations in the Hilly and Gully Region of the Loess Plateau in China. Agric. Ecosyst. Environ. 2023, 343, 108256. [Google Scholar] [CrossRef]
- Wang, H.; Liu, W.; Wang, W.; Zu, Y. Influence of Long-Term Thinning on the Biomass Carbon and Soil Respiration in a Larch (Larix gmelinii) Forest in Northeastern China. Sci. World J. 2013, 2013, 865645. [Google Scholar] [CrossRef]
- Noh, N.J.; Kim, C.; Bae, S.W.; Lee, W.K.; Yoon, T.K.; Muraoka, H.; Son, Y. Carbon and Nitrogen Dynamics in a Pinus densiflora Forest with Low and High Stand Densities. J. Plant Ecol. 2013, 6, 368–379. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Density Gradient /(trees ha−1) | Current Density /(trees ha−1) | DBH /(cm) | H /(m) | Slope /(°) | Age /(a) | Orientation /(°) |
|---|---|---|---|---|---|---|
| D1 (n = 11) | 900−1400 | 13.22 ± 1.92 a | 10.30 ± 0.22 a | 20–30 | 30 | NW-NE |
| D2 (n = 15) | 1401−1900 | 11.62 ± 1.23 ab | 9.75 ± 0.29 a | 20–30 | 30 | NW-NE |
| D3 (n = 13) | 1901−2400 | 11.40 ± 1.29 ab | 9.12 ± 0.31 a | 20–30 | 30 | NW-NE |
| D4 (n = 8) | 2401−2900 | 10.59 ± 0.83 ab | 9.01 ± 0.27 a | 20–30 | 30 | NW-NE |
| D5 (n = 2) | 2901−3400 | 9.79 ± 0.74 b | 8.87 ± 0.79 a | 20–30 | 30 | NW-NE |
| Type | Organ | Fitted Model | Adjusted R2 | Number of Fitted Samples | Number of Samples for Verification | Error |
|---|---|---|---|---|---|---|
| I | Leaf | W = 0.0011(DBH2H))1.0080 | 0.70 | 34 | 15 | 21.46% |
| Branch | W = 0.01(DBH2H))0.84 | 0.72 | 34 | 15 | 26.15% | |
| Stem | W = 0.19(DBH2H))0.69 | 0.79 | 34 | 15 | 11.71% | |
| Root | W = 0.03(DBH2H))0.78 | 0.81 | 34 | 15 | 23.50% | |
| Total | W = 0.28(DBH2H))0.70 | 0.90 | 34 | 15 | 10.55% | |
| II | Leaf | W = 0.0014(1 + DBH)2.7957 | 0.73 | 34 | 15 | 21.62% |
| Branch | W = 0.02(1 + DBH)2.35 | 0.75 | 34 | 15 | 24.34% | |
| Stem | W = 0.25(1 + DBH)1.88 | 0.76 | 34 | 15 | 13.37% | |
| Root | W = 0.13(1 + DBH)1.72 | 0.84 | 34 | 15 | 21.65% | |
| Total | W = 0.35(1 + DBH)1.94 | 0.89 | 34 | 15 | 10.59% |
| Density Gradient /(trees ha−1) | Current Density /(trees ha−1) | Leaf /(kg) | Branch /(kg) | Stem /(kg) | The Total Aboveground /(kg) |
|---|---|---|---|---|---|
| D1 (n = 11) | 900–1400 | 2.97 ± 0.37 a | 11.32 ± 1.08 a | 38.66 ± 3.30 a | 52.95 ± 4.53 a |
| D2 (n = 15) | 1401–1900 | 2.09 ± 0.15 ab | 8.68 ± 0.54 ab | 30.15 ± 1.57 ab | 40.92 ± 2.26 ab |
| D3 (n = 13) | 1901–2400 | 1.76 ± 0.16 b | 7.30 ± 0.47 bc | 26.58 ± 1.74 bc | 35.63 ± 2.36 bc |
| D4 (n = 8) | 2401–2900 | 1.37 ± 0.10 b | 5.96 ± 0.35 bc | 21.97 ± 1.06 bc | 29.29 ± 1.50 bc |
| D5 (n = 2) | 2901–3400 | 1.22 ± 0.41 b | 5.25 ± 1.44 c | 19.40 ± 4.05 c | 25.86 ± 5.90 c |
| Density Gradient /(trees ha−1) | Current Density /(trees ha−1) | Coarse /(kg) | Fine /(kg) | The Total Belowground /(kg) | The Total /(kg) |
|---|---|---|---|---|---|
| D1 (n = 11) | 900–1400 | 10.25 ± 0.99 a | 2.65 ± 0.23 a | 12.89 ± 0.99 a | 62.68 ± 4.32 a |
| D2 (n = 15) | 1401–1900 | 7.73 ± 0.45 ab | 2.90 ± 0.15 a | 10.63 ± 0.49 ab | 51.55 ± 2.73 ab |
| D3 (n = 13) | 1901–2400 | 6.71 ± 0.50 b | 2.07 ± 0.16 ab | 8.78 ± 0.49 bc | 44.41 ± 2.83 bc |
| D4 (n = 8) | 2401–2900 | 5.43 ± 0.30 b | 1.72 ± 0.19 b | 7.16 ± 0.39 c | 36.46 ± 1.85 bc |
| D5 (n = 2) | 2901–3400 | 4.78 ± 1.18 b | 1.38 ± 0.16 b | 6.17 ± 1.35 c | 32.02 ± 7.25 c |
| Density (trees ha−1) | Leaf Biomass (t ha−1) | Branch Biomass (t ha−1) | Stem Biomass (t ha−1) | Coarse Root Biomass (t ha−1) | Fine Root Biomass (t ha−1) | R. pseudoacacia Biomass (t ha−1) |
|---|---|---|---|---|---|---|
| 900–1400 | 3.02 (4.21%) | 12.91 (18.00%) | 41.78 (58.25%) | 10.87 (15.16%) | 3.15 (4.38%) | 71.72 |
| 1401–1900 | 3.08 (3.98%) | 12.88 (16.66%) | 45.28 (58.58%) | 11.53 (14.92%) | 4.53 (5.86%) | 77.30 |
| 1901–2400 | 3.21 (3.83%) | 13.84 (16.51%) | 49.97 (59.60%) | 12.51 (14.92%) | 4.31 (5.14%) | 83.84 |
| 2401–2900 | 3.56 (3.79%) | 15.40 (16.39%) | 56.25 (59.87%) | 13.99 (14.89%) | 4.75 (5.06%) | 93.96 |
| 2901–3400 | 3.79 (3.78%) | 16.38 (16.34%) | 60.75 (60.62%) | 14.95 (14.92%) | 4.35 (4.34%) | 100.22 |
| Density (trees ha−1) | Aboveground or Belowground | Shrub (t ha−1) | Herb (t ha−1) | R. pseudoacacia (t ha−1) | Total (t ha−1) |
|---|---|---|---|---|---|
| 900–1400 | Aboveground biomass | 2.69 (4.30%) | 2.15 (3.44%) | 57.70 (92.26%) | 62.54 |
| Belowground biomass | 0.54 (3.71%) | 14.02 (96.29%) | 14.56 | ||
| Total biomass | 5.38 (6.98%) | 71.72 (93.02%) | 77.10 | ||
| 1401–1900 | Aboveground biomass | 3.20 (4.84%) | 1.60 (2.42%) | 61.24 (92.74%) | 66.04 |
| Belowground biomass | 0.96 (5.64%) | 16.06 (94.36%) | 17.02 | ||
| Total biomass | 5.76 (6.93%) | 77.3 (93.07%) | 83.06 | ||
| 1901–2400 | Aboveground biomass | 1.18 (1.68%) | 1.84 (2.63%) | 67.02 (95.69%) | 70.04 |
| Belowground biomass | 1.03 (5.77%) | 16.83 (94.23%) | 17.86 | ||
| Total biomass | 4.06 (4.62%) | 83.84 (95.38%) | 87.90 | ||
| 2401–2900 | Aboveground biomass | 1.23 (1.58%) | 1.65 (2.11%) | 75.21 (96.31%) | 78.09 |
| Belowground biomass | 0.58 (3.00%) | 18.75 (97.00%) | 19.33 | ||
| Total biomass | 3.47 (3.56%) | 93.95 (96.44%) | 97.42 | ||
| 2901–3400 | Aboveground biomass | 1.05 (1.27%) | 0.57 (0.69%) | 80.91 (98.04%) | 82.53 |
| Belowground biomass | 1.09 (5.34%) | 19.30 (94.66%) | 20.39 | ||
| Total biomass | 2.71 (2.63%) | 100.21 (97.37%) | 102.92 | ||
| Density (trees ha−1) | Vegetation (t ha−1) | Soil Organic Carbon (t ha−1) | Litter (t ha−1) | Total (t ha−1) |
|---|---|---|---|---|
| 900−1400 | 38.37 (45.92%) | 43.23 (51.74%) | 1.96 (2.34%) | 83.56 |
| 1401−1900 | 41.34 (49.60%) | 39.79 (47.74%) | 2.22 (2.66%) | 83.35 |
| 1901−2400 | 43.82 (51.82%) | 38.29 (45.28%) | 2.45 (2.90%) | 84.56 |
| 2401−2900 | 48.60 (53.38%) | 39.78 (43.69%) | 2.67 (2.93%) | 91.05 |
| 2901−3400 | 51.37 (49.78%) | 48.84 (47.34%) | 2.97 (2.88%) | 103.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Zhao, J.; Li, Y.; Tang, P.; Yang, Z.; Zhang, J.; Sun, R. Biomass and Carbon Stock Capacity of Robinia pseudoacacia Plantations at Different Densities on the Loess Plateau. Forests 2024, 15, 1242. https://doi.org/10.3390/f15071242
Hu Y, Zhao J, Li Y, Tang P, Yang Z, Zhang J, Sun R. Biomass and Carbon Stock Capacity of Robinia pseudoacacia Plantations at Different Densities on the Loess Plateau. Forests. 2024; 15(7):1242. https://doi.org/10.3390/f15071242
Chicago/Turabian StyleHu, Yawei, Jiongchang Zhao, Yang Li, Peng Tang, Zhou Yang, Jianjun Zhang, and Ruoxiu Sun. 2024. "Biomass and Carbon Stock Capacity of Robinia pseudoacacia Plantations at Different Densities on the Loess Plateau" Forests 15, no. 7: 1242. https://doi.org/10.3390/f15071242
APA StyleHu, Y., Zhao, J., Li, Y., Tang, P., Yang, Z., Zhang, J., & Sun, R. (2024). Biomass and Carbon Stock Capacity of Robinia pseudoacacia Plantations at Different Densities on the Loess Plateau. Forests, 15(7), 1242. https://doi.org/10.3390/f15071242