
Germplasm Resource Status and Seed Adaptability of Nypa fruticans Wurmb, an Endangered Species in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Methods
2.2.1. Resource Survey
- (1)
- Sample Plot Setting and Investigation
- (2)
- Parameter Analysis of Community Characteristics
2.2.2. Indoor Simulation Experiments
- (1)
- Experimental Design
- (2)
- Parameter Measurements
2.3. Data Statistics and Analysis
3. Results
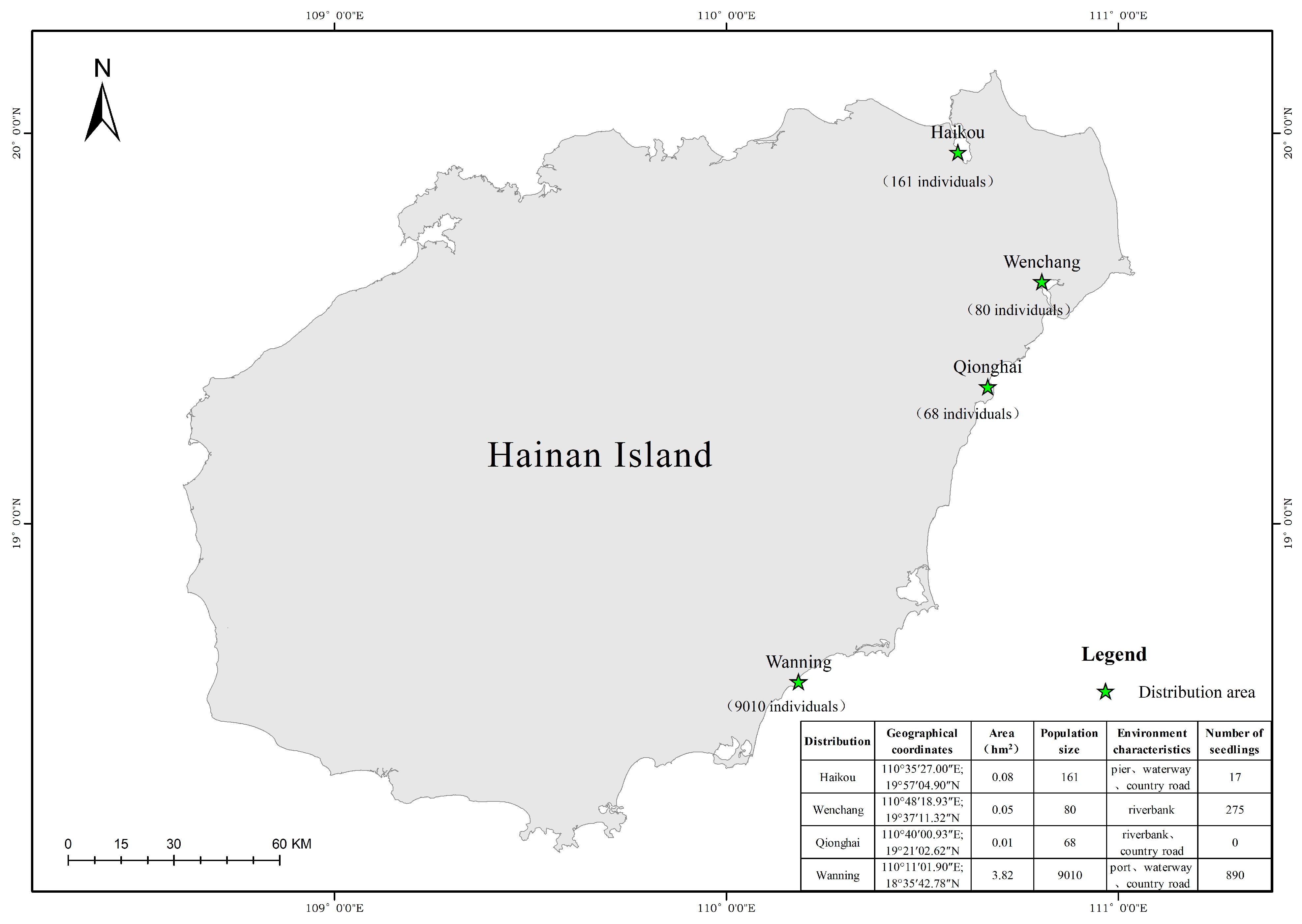
3.1. Geographical Distribution and Resource Status of N. fruticans
3.2. Community Characteristics of N. fruticans
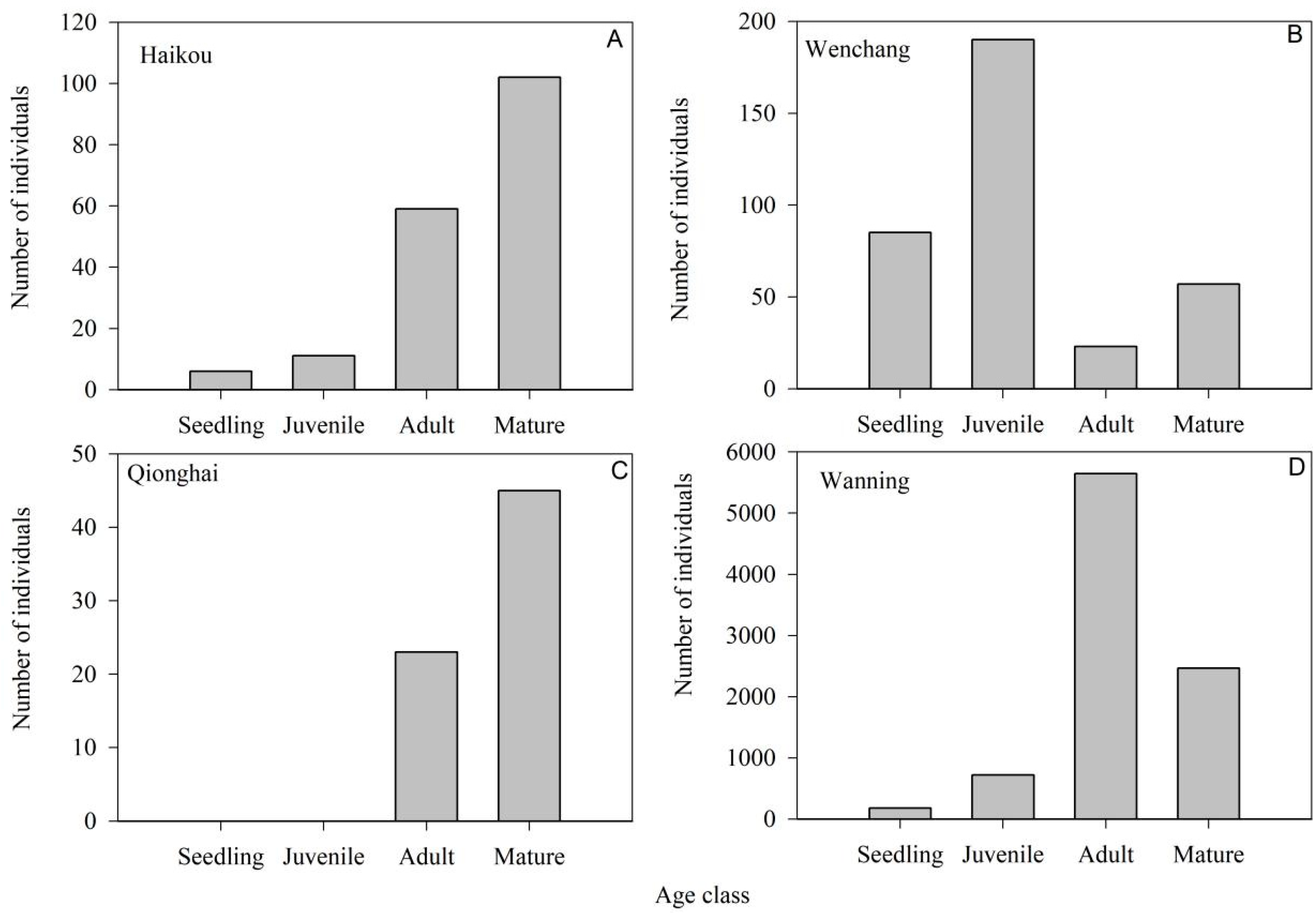
3.3. Population Structure of N. fruticans
3.4. The Effects of Different Environmental Factors on the Seed Germination of N. fruticans
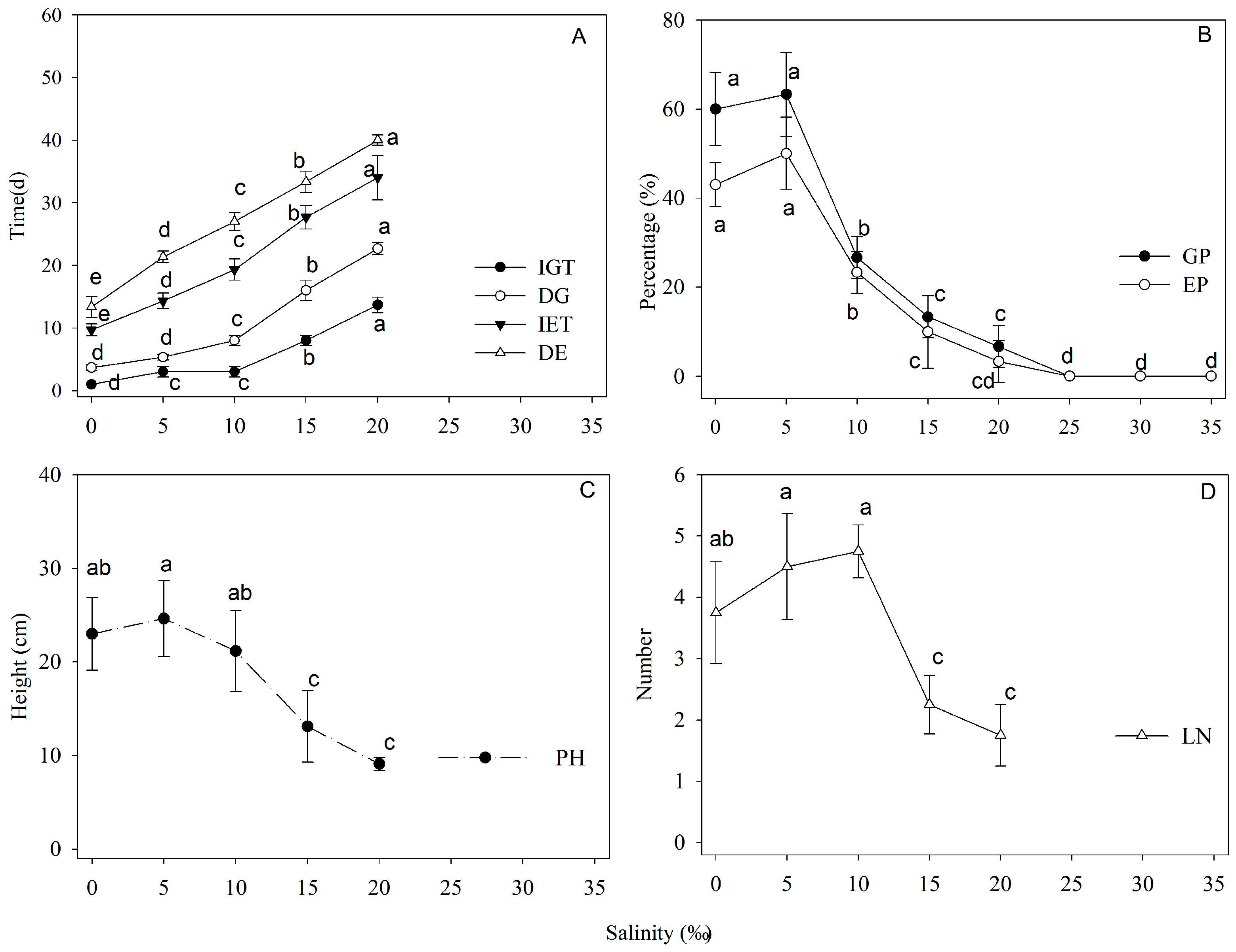
3.4.1. The Effect of Light Intensity on Seed Germination
3.4.2. The Effect of Salinity on Seed Germination
3.4.3. The Effect of Flooding Time on Seed Germination
4. Discussion
4.1. Resource Status and Population Characteristics of N. fruticans
4.2. The Effects of Simulated Environmental Factors on N. fruticans Seed Germination
4.2.1. The Effect of Light Intensity on N. fruticans Seed Germination
4.2.2. The Effect of Salinity on N. fruticans Seed Germination
4.2.3. The Effect of Flooding Time on N. fruticans Seed Germination
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Family | Genus | Species |
|---|---|---|---|
| True mangrove | |||
| 1 | Acanthaceae | Avicennia | Avicennia marina (Forssk.) Vierh |
| 2 | Lythraceae | Sonneratia | Sonneratia alba Sm. in Ress |
| 3 | Rhizophoraceae | Bruguiera | Bruguiera sexangula (Lour.) Poir |
| 4 | Euphorbiaceae | Excoecaria | Excoecaria agallocha L. |
| 5 | Lythraceae | Sonneratia | Sonneratia caseolaris (L.) Engler |
| 6 | Rhizophoraceae | Rhizophora | Rhizophora stylosa Griff |
| 7 | Lythraceae | Sonneratia | Sonneratia apetala Buch.—Ham |
| 8 | Rhizophoraceae | Ceriops | Ceriops tagal (Perr.) C. B. Rob |
| 9 | Combretaceae | Lumnitzera | Lumnitzera racemosa Willd |
| 10 | Combretaceae | Laguncularia | Laguncularia racemosa (L.) C. F. Gaertn |
| 11 | Acanthaceae | Acanthus | Acanthus ilicifolius L. |
| 12 | Pteridaceae | Acrostichum | Acrostichum aureum L. |
| 13 | Lythraceae | Sonneratia | Sonneratia ovata Backer |
| 14 | Meliaceae | Xylocarpus | Xylocarpus granatum J. Koenig |
| 15 | Rhizophoraceae | Bruguiera | Bruguiera gymnorhiza (L.) Savigny |
| 16 | Rhizophoraceae | Kandelia | Kandelia obovata Sheue and al |
| 17 | Arecaceae | Nypa | Nypa fruticans Wurmb |
| 18 | Primulaceae | Aegiceras | Aegiceras corniculatum (L.) Blanco |
| Semi-mangrove | |||
| 19 | Bignoniaceae | Dolichandrone | Dolichandrone spathacea (L. f.) Seem |
| 20 | Fabaceae | Pongamia | Pongamia pinnata (L.) Pierre |
| 21 | Lamiaceae | Volkameria | Volkameria inermis L. |
| 22 | Malvaceae | Heritiera | Heritiera littoralis Dryand |
| 23 | Malvaceae | Talipariti | Talipariti tiliaceum (L.) Fryxell |
References
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Sonia, H.M.; Qian, S.H.; Yang, Y.C.; Ohsawa, M.; Luu, H.T.; et al. Identifying long–term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 5241. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Ohsawa, M. Tertiary relic deciduous forests on a subtropical mountain, Mt. Emei, Sichuan, China. Folia Geobot. 2002, 37, 93–106. [Google Scholar] [CrossRef]
- Tang, C.Q.; Yang, Y.C.; Ohsawa, M.; Arata, M.; Yi, S.R.; Kevin, R.; Song, K.; Zhang, S.Q.; He, L.Y. Community structure and survival of tertiary relict Thuja sutchuenensis(Cupressaceae) in the subtropical Daba Mountains, Southwestern China. PLoS ONE 2015, 10, e0125307. [Google Scholar] [CrossRef] [PubMed]
- He, L.Y.; Tang, C.Q.; Wu, Z.L.; Wang, H.C.; Ohsawa, M.; Yan, K. Forest structure and regeneration of the tertiary relict Taiwania cryptomerioides in the Gaoligong mountains, Yunnan, southwestern China. Phytocoenologia 2015, 45, 135–156. [Google Scholar] [CrossRef]
- Qian, S.; Yang, Y.; Tang, C.Q.; Momohara, A.; Yi, S.; Ohsawa, M. Effective conservation measures are needed for wild Cathaya argyrophylla populations in China: Insights from the population structure and regeneration characteristics. For. Ecol. Manag. 2016, 361, 358–367. [Google Scholar] [CrossRef]
- Lovly, M.S.; Merlee-Teresa, M.V. Nypa palm (Nypa fruticans Wurmb.) A new record from Kerala. Int. J. Adv. Res. 2016, 4, 1051–1055. [Google Scholar]
- Tang, C.Q.; Yang, Y.C.; Ohsawa, M.; Momohara, M.; Hara, M.; Chen, S.L.; Fan, S.H. Population structure of relict Metasequoia glyptostroboides and its habitat fragmentation and degradation in south–central China. Biol. Conserv. 2011, 144, 279–289. [Google Scholar] [CrossRef]
- Xu, Z.H.; Rena, H.; Wei, X.; Ouyang, K.; Li, D.X.; Guo, Y.L.; Wen, S.J.; Long, J.F.; Wang, J.; Hui, D.F. Distribution and conservation status of Camellia longzhouensis (Theaceae), a critically endangered plant species endemic to southern China. Glob. Ecol. Conserv. 2021, 27, e01585. [Google Scholar] [CrossRef]
- Volis, S. How to conserve threatened Chinese plant species with extremely small populations? Plant Divers. 2016, 38, 45–52. [Google Scholar] [CrossRef]
- Sanger, P.; Hegerl, E.J.; Davie, J.D.S. Global status of mangrove ecosystems. Environmentalist 1983, 3, 7–80. [Google Scholar]
- Zhu, J.J. Differences in water and carbon use strategies for photosynthesis in true mangrove and semi-mangrove plants. Plant Physiol. J. 2024, 60, 989–997. [Google Scholar]
- Zakaria, R.M.; Aslezaeim, N.; Sofawi, A.B. Effects of water properties and soil texture on the growth of a mangrove palm; Nypa fruticans on carey Island, Malaysia. Pak. J. Bot. 2017, 49, 33–39. [Google Scholar]
- Moudingo, J.H.; Ajonina, G.; Dibong, D.; Tomedi, M. Chapter 3: Distribution, Devastating Effect, and Drivers of the Exotic Mangrove Nypa fruticans Van Wurmb (Arecaceae) on the Mangroves of West and Central Africa; Academic Press: Pittsburgh, PA, USA, 2020; pp. 24–36. [Google Scholar]
- Rozainah, M.Z.; Aslezaeim, N. A demographic study of a mangrove palm, Nypa fruticans. Sci. Res. Essays 2010, 5, 3896–3902. [Google Scholar]
- Hossain, M.F.; Islam, M.A. Utilization of mangrove forest plant: Nipa Palm (Nypa fruticans Wurmb.). Am. J. Agric. For. 2015, 3, 156–160. [Google Scholar]
- Tan, Y.H.; Chen, Z. Biological characteristics and conservation countermeasures of the rare and endangered Nypa fruticans. Mod. Hortic. 2013, 23, 31–32. [Google Scholar]
- Gee, C.T. The mangrove palm Nypa in the geologic past of the New World. Wetl. Ecol. Manag. 2001, 9, 181–194. [Google Scholar] [CrossRef]
- Lin, Y.R. On the systematics, evolution, floristics and economic uses of Palmae and its sister family, Calamaceae. Bull. Bot. Res. 2002, 3, 341–365. [Google Scholar]
- Jian, S.G.; Ban, J.W.; Ren, H.; Yan, H.F. Low genetic variation detected within the widespread mangrove species Nypa fruticans (Palmae) from Southeast Asia. Aquat. Bot. 2010, 92, 23–27. [Google Scholar] [CrossRef]
- Middeljans, M. The Species Composition of the Mangrove Forest along the Abatan River in Lincod, Maribojoc, Bohol, Philippines and the Mangrove Forest Structure and Its Regeneration Status between Managed and Unmanaged Nipa Palm (Nypa fruticans Wurmb). Bachelor’s Thesis, Van Hall Larenstein University, Leeuwarden, The Netherlands, 2015. [Google Scholar]
- Teo, S.; Kurukulasuriya, B.R. The status and distribution of the nipah palm, Nypa fruticans Wurmb. (Arecaceae) in Singapore. Nat. Singap. 2010, 3, 45–52. [Google Scholar]
- Mondal, S.; Basu, S.K.; Chowdhury, M. Observation on Nypa fruticans. Phytotaxonomy 2018, 17, 39–42. [Google Scholar]
- Chen, L.Z.; Gu, X.X.; Zhong, C.R.; Huang, L.H. The Wetlands of Haikou (Mangroves); Xiamen University Press: Xiamen, China, 2019; pp. 84–102. [Google Scholar]
- Borja, J.A.; Silveira, F.A.O.; Fidelis, A.; Poschlod, P.; Commander, E.L. Seed germination traits can contribute better to plant community ecology. J. Veg. Sci. 2016, 27, 637–645. [Google Scholar]
- Jiang, H.D.; Xie, W.L.; Chai, S.F.; Tang, J.M.; Jiang, Y.S.; Qin, H.Z.; Wei, X. Seed germination characteristics of Keteleeria calcarea, a precious tree species in karst areas. Guihaia 2022, 42, 951–960. [Google Scholar]
- Robert, E.M.; Oste, J.; Vander–Stocken, T.; De–Ryck, D.J.; Quisthoudt, K.; Kairo, J.G.; Dahdouh–Guebas, F.; Koedam, N.; Schmitz, N. Viviparous mangrove propagules of Ceriops tagal and Rhizophora mucronata, where both Rhizophoraceae show different dispersal and establishment strategies. J. Exp. Mar. Biol. Ecol. 2015, 468, 45–54. [Google Scholar] [CrossRef]
- Zhou, X.X.; Cai, L.L.; Fu, M.P.; Hong, L.W.; Shen, Y.J.; Li, Q.S. Progress in the studies of vivipary in mangrove plants. Chin. J. Plant Ecol. 2016, 40, 1328–1343. [Google Scholar]
- Liu, S.S.; Yang, S.; Liu, X.; Chen, Q.H.; Jiang, Z.M. Research progress in the adaptability of mangroves to tidal inundation. World For. Res. 2022, 35, 25–30. [Google Scholar]
- Yan, Z.Z.; Wang, W.Q.; Huang, W.B. Development of the viviparous hypocotyl of mangrove and its adaptation to inter–tidal habitats: A review. Acta Ecol. Sin. 2004, 10, 2317–2323. [Google Scholar]
- Zhou, X.X.; Weng, Y.L.; Su, W.Y.; Ye, C.T.; Qu, H.D.; Li, Q.S. Uninterrupted embryonic growth leading to viviparous propagule formation in mangrove. Front. Plant Sci. 2023, 13, 1061747. [Google Scholar] [CrossRef] [PubMed]
- Duke, N.C.; Watkinson, A.J. Chlorophyll-deficient propagules of Avicennia marina and apparent longer term deterioration of mangrove fitness in oil-polluted sediments. Mar. Pollut. Bull. 2002, 44, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.E.; Liao, B.W. The physio–ecological response of Acanthus ilicifolius seedlings to different degrees of light intensity in tide environment. For. Res. 2013, 26, 192–199. [Google Scholar]
- Ulqodry, T.Z.; Matsumoto, F.; Okimoto, Y.; Nose, A.; Zheng, S.H. Study on photosynthetic responses and chlorophyll fluorescence in Rhizophora mucronata seedlings under shade regimes. Acta Physiol. Plant. 2014, 36, 1903–1917. [Google Scholar] [CrossRef]
- Dangremond, E.M.; Feller, I.C.; Sousa, W.P. Environmental tolerances of rare and common mangroves along light and salinity gradients. Oecologia 2015, 179, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.M.; Tang, H.Y.; Liu, Y.; Zhang, M.Q.; Jiang, S.S.; Yang, F.; Li, X.Y.; Wang, C.Y. Resource status and protection strategies of mangroves in China. J. Coast. Conserv. 2021, 25, 42. [Google Scholar] [CrossRef]
- Fang, J.Y.; Wang, X.P.; Shen, Z.H.; Tang, Z.Y.; He, J.S.; Yu, D.; Jiang, Y.; Wang, Z.H.; Zheng, C.Y.; Zhu, J.L.; et al. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533–548. [Google Scholar]
- Kiruba-Sankar, R.; Krishnan, P.; Roy, S.D.; Angel, J.R.J.; Ramesh, R. Structural complexity and tree species composition of mangrove forests of the Andaman Islands, India. J. Coast. Conserv. 2017, 22, 217–234. [Google Scholar] [CrossRef]
- Hu, G.; Li, J.; Qin, Y.Y.; Hu, B.Q.; Liu, X.; Zhang, Z.H. Population structure and dynamics of mangrove species in Beilun Estuary, Guangxi, southern China. Acta Ecol. Sin. 2018, 38, 3022–3034. [Google Scholar]
- Wijayasinghe, M.M.; Jayasuriya, K.G.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N.; Walck, J.L. Effect of salinity on seed germination of five mangroves from SriLanka: Use of hydrotime modelling for mangrove germination. Seed Sci. Res. 2018, 29, 55–63. [Google Scholar]
- Mo, Z.C.; Fan, H.Q.; He, B. Effects of seawater salinity on hypocotyl growth in two mangrove species. Acta Phytoecol. Sin. 2001, 25, 235–239. [Google Scholar]
- Zhang, Y.; Ye, Y.; Lu, C.Y. Seed germination and seedling growth of mangrove Excoecaria agallocha under different salinities. J. Xiamen Univ. Nat. Sci. 2010, 49, 144–148. [Google Scholar]
- Muller, J. Palynology of the recent Orinoco delta and shelf sediments. Micropaleontology 1959, 5, 1–32. [Google Scholar] [CrossRef]
- Wu, E.H.; Li, D.H.; Yang, X.B.; Zuo, Y.L.; Yang, N. Study on distribution characteristics and population dynamics of wild Cycas Hainanensis in Hainan Island. For. Resour. Manag. 2021, 4, 130–137. [Google Scholar]
- Liu, Q.Q.; Ma, X.Q.; Li, Y.J.; Zhuang, Z.; Du, Z.L.; Xing, X.S.; Liu, B. Response of seed germination and seedling growth of Chinese fir to different light intensities. Chin. J. Appl. Ecol. 2016, 27, 3845–3852. [Google Scholar]
- Zhang, M.W.; Yang, X.B.; Long, W.X.; Li, D.H.; Lv, X.B. Reasons for the extremely small population of putative hybrid Sonneratia × hainanensis W.C.Ko (Lythraceae). Forests 2019, 10, 526. [Google Scholar] [CrossRef]
- Ren, F.Y.; Zhang, F.; Zhang, L.N.; Wang, S.Q. Seed germination characteristics of the endangered mangrove plant Onychophora maritima. Mol. Plant Breed. 2021, 19, 5150–5156. [Google Scholar]
- Zhang, M.W. Comparative Study on Environmental Adaptability of Population Development between Two Mangrove Natural Hybrids and Their Parents in Hainan Island. Ph.D. Thesis, Hainan University, Haikou, China, 2019. [Google Scholar]
- Su, H.; Shen, Y.R.; Cai, J.; Jiang, Z.M.; Yang, C.R.; Xu, X.Q. Germination characteristics of Betula albosinensis seeds from different provenances. J. Northwest For. Acad. 2021, 36, 109–114. [Google Scholar]
- Zhang, M.W.; Zhong, C.R.; Lv, X.B.; Fang, Z.S.; Cheng, C. Studies on the relict mangrove plant of Nypa fruticans Wurmb in the World. World For. Res. 2022, 35, 20–24. [Google Scholar]
- Liu, Q.Q.; Huang, Z.J.; Guo, S.; Wang, D.Y.; Wang, C.H.; Wang, Z.N.; Ma, X.Q.; Liu, B. Responses of seed germination and seedling growth of Cunninghamia lanceolata and Schima superba to different light intensities. Chin. J. Appl. Ecol. 2019, 30, 2955–2963. [Google Scholar]
- Han, D.Y.; Zhang, W.; Nurmati, Y.; Yang, Y.F. Recruitment limitation of plant population regeneration. Chin. J. Plant Ecol. 2021, 45, 1–12. [Google Scholar] [CrossRef]
- Lv, X.B.; Li, D.H.; Yang, X.B.; Zhang, M.W.; Deng, Q. Leaf enzyme and plant productivity responses to environmental stress associated with sea level rise in two Asian mangrove species. Forests 2019, 10, 250–262. [Google Scholar] [CrossRef]
- Orlovsky, N.; Japakova, U.; Zhang, H.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers. 2016, 38, 183–189. [Google Scholar] [CrossRef]
- Saberali, S.F.; Moradi, M. Effect of salinity on germination and seedling growth of Trigonella foenumgraecum, Dracocephalum moldavica, Satureja hortensis and Anethum graveolens. J. Saudi Soc. Agric. Sci. 2019, 18, 316–323. [Google Scholar]
- Luo, M.J.; Zhang, S.G.; Cui, L.J.; Tan, F.L.; Huang, Y.R. Response of growth and biomass allocation of Aegiceras corniculatum to waterlogging stress. J. Zhejiang For. Sci. Technol. 2012, 32, 15–19. [Google Scholar]
- Liao, B.W. The Adaptability of Seedling of Three Mangrove Species to Tide-Flooding and Water Salinity. Ph.D. Thesis, China Academy of Forestry, Beijing, China, 2010. [Google Scholar]
- Dai, J. Adaptation Mechanism of Mangrove Seedlings of Kandelia obovata to Tidal Waterlogging. Master Thesis, East China Normal University, Shanghai, China, 2018. [Google Scholar]
- Zhang, Y. Research on the waterlogging stress impact on Avicennia Marina seed germination and its seedling growth. Trop. For. 2018, 46, 35–38. [Google Scholar]
- Zhao, L.L.; Mo, B.T.; Wang, P.C.; Zhang, Y.; Long, Z.F. Relationship of Sophora davidii seed size to germination, dormancy, and mortality under water stress. S. Afr. J. Bot. 2015, 99, 12–16. [Google Scholar] [CrossRef]


| Factor | Treatment Group | Light Intensity Treatment (%) | Salinity Treatment (‰) | Flooding Time Treatment (h/d) |
|---|---|---|---|---|
| Light intensity | 1 | 20 | 5 | 4 |
| 2 | 40 | 5 | 4 | |
| 3 | 60 | 5 | 4 | |
| 4 | 100 | 5 | 4 | |
| Salinity | 1 | 60 | 0 | 4 |
| 2 | 60 | 5 | 4 | |
| 3 | 60 | 10 | 4 | |
| 4 | 60 | 15 | 4 | |
| 5 | 60 | 20 | 4 | |
| 6 | 60 | 25 | 4 | |
| 7 | 60 | 30 | 4 | |
| 8 | 60 | 35 | 4 | |
| Flooding time | 1 | 60 | 5 | 2 |
| 2 | 60 | 5 | 4 | |
| 3 | 60 | 5 | 6 | |
| 4 | 60 | 5 | 8 | |
| 5 | 60 | 5 | 10 | |
| 6 | 60 | 5 | 12 | |
| 7 | 60 | 5 | 14 | |
| 8 | 60 | 5 | 16 | |
| 9 | 60 | 5 | 18 | |
| 10 | 60 | 5 | 20 | |
| 11 | 60 | 5 | 22 |
| Flooding Time (h) | Injection Water Time | Discharge Water Time |
|---|---|---|
| 2 | 0:00; 12:00 | 1:00; 13:00 |
| 4 | 0:00; 12:00 | 2:00; 14:00 |
| 6 | 0:00; 12:00 | 3:00; 15:00 |
| 8 | 0:00; 12:00 | 4:00; 16:00 |
| 10 | 0:00; 12:00 | 5:00; 17:00 |
| 12 | 0:00; 12:00 | 6:00; 18:00 |
| 14 | 0:00; 12:00 | 7:00; 19:00 |
| 16 | 0:00; 12:00 | 8:00; 20:00 |
| 18 | 0:00; 12:00 | 9:00; 21:00 |
| 20 | 0:00; 12:00 | 10:00; 22:00 |
| 22 | 0:00; 12:00 | 11:00; 23:00 |
| Layer | Haikou | Wenchang | Qionghai | Wanning | ||||
|---|---|---|---|---|---|---|---|---|
| Species | Importance Values (%) | Species | Importance Values (%) | Species | Importance Values (%) | Species | Importance Values (%) | |
| Tree layer | Bruguiera sexangula | 31.57 | Sonneratia caseolaris | 59.19 | Talipariti tiliaceum | 100.00 | Bruguiera sexangula | 33.25 |
| Rhizophora stylosa | 18.68 | Bruguiera sexangula | 20.28 | Laguncularia racemosa | 20.51 | |||
| Sonneratia caseolaris | 11.66 | Talipariti tiliaceum | 17.24 | Talipariti tiliaceum | 18.90 | |||
| Sonneratia apetala | 7.52 | Bruguiera gymnorhiza | 1.64 | Excoecaria agallocha | 13.87 | |||
| Ceriops tagal | 4.94 | Aegiceras corniculatum | 1.64 | Sonneratia apetala | 3.58 | |||
| Shrub layer | Nypa fruticans | 70.70 | Nypa fruticans | 58.82 | Nypa fruticans | 100.00 | Nypa fruticans | 62.70 |
| Aegiceras corniculatum | 29.30 | Acanthus ilicifolius | 41.18 | Bruguiera sexangula | 25.44 | |||
| Laguncularia racemosa | 6.48 | |||||||
| Volkameria inermis | 3.84 | |||||||
| Acanthus ilicifolius | 1.53 | |||||||
| Herb layer | Acrostichum aureum | 100.00 | Acrostichum aureum | 100.00 | Acrostichum aureum | 100.00 | Acrostichum aureum | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhong, C.; Lv, X.; Fang, Z.; Cheng, C.; Hao, J. Germplasm Resource Status and Seed Adaptability of Nypa fruticans Wurmb, an Endangered Species in China. Forests 2024, 15, 1396. https://doi.org/10.3390/f15081396
Zhang M, Zhong C, Lv X, Fang Z, Cheng C, Hao J. Germplasm Resource Status and Seed Adaptability of Nypa fruticans Wurmb, an Endangered Species in China. Forests. 2024; 15(8):1396. https://doi.org/10.3390/f15081396
Chicago/Turabian StyleZhang, Mengwen, Cairong Zhong, Xiaobo Lv, Zanshan Fang, Cheng Cheng, and Jiewei Hao. 2024. "Germplasm Resource Status and Seed Adaptability of Nypa fruticans Wurmb, an Endangered Species in China" Forests 15, no. 8: 1396. https://doi.org/10.3390/f15081396