
Is It Possible to Predict a Forest Insect Outbreak? Backtesting Using Remote Sensing Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
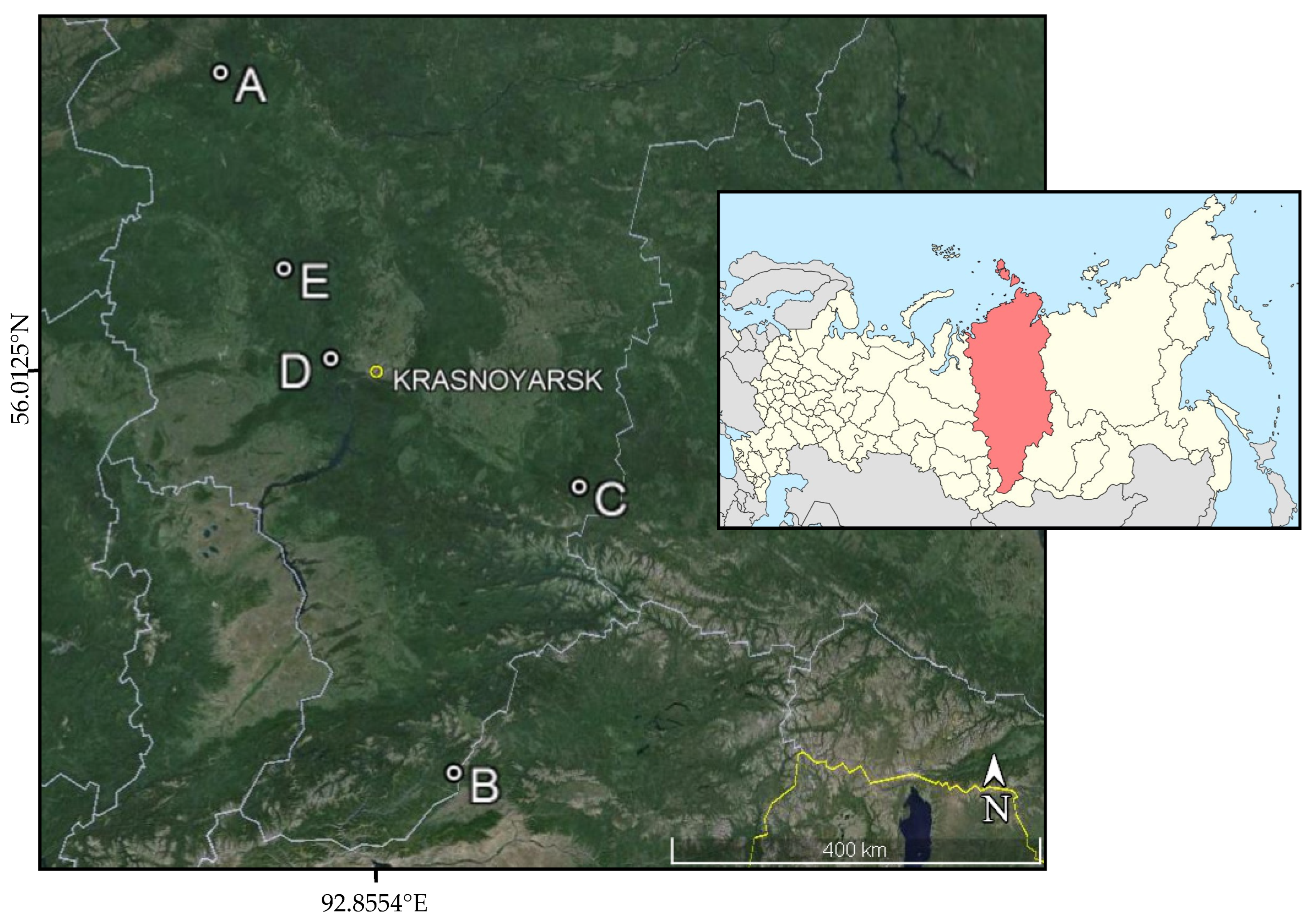
2.1. Field Study Locations and Characterization of the Study Areas
- (1)
- The Siberian silk moth D. sibiricus is a large butterfly with a wingspan of 60–80 mm in the female and 40–60 mm in the male. Its color varies from light yellowish brown or light gray to almost black. Females lay eggs on needles of coniferous trees, mainly in the lower part of the crown. In one clutch, there are usually several dozen eggs (up to 200), and in total, a female can lay up to 800 eggs; however, most often, the fertility does not exceed 200–300 eggs. The caterpillars feed on needles of almost all coniferous species but prefer fir Abies sibirica Ledeb, spruce Picea obovata Ledeb., and larch Larix sibirica Ldb. [35,36,37].
- (2)
- The white mottled sawyer M. urussovi is a large beetle with a black body and appendages and with brighter yellowish apices of the elytra. It lives within the geographic range of dark coniferous forests and prefers A. sibirica and P. obovata. It is one of the most formidable pests that cause considerable damage to coniferous forests of Siberia. Its larvae develop under the bark and in the wood of coniferous trees for 2, or less often, 3 years, although under favorable conditions, they can go through the developmental cycle within 1 year. Within their geographic range, these beetles inhabit coniferous, less often mixed, forests dominated by fir A. sibirica and/or spruce P. obovata as well as in the cutting areas and coniferous stands that are damaged by D. sibiricus and Lymantria dispar L. M. urussovi also occurs in mountain coniferous forests, usually at an altitude of up to 2000 m above sea level. There are known outbreaks of M. urussovi covering territories of tens and hundreds of thousands of hectares that have led to massive suppression of fir forests [38,39].
- (3)
- The four-eyed fir bark beetle (P. proximus Blandford) is a species of bark beetle. It is a dangerous invasive dendrophagous pest of fir A. sibirica. The penetration of P. proximus into taiga ecosystems of Siberia and the formation of P. proximus outbreak foci in them represent the only known case of large-scale insect invasion in this territory today. It damages A. sibirica, and less often, P. obovata and P. sibirica. P. proximus is one of the main causes of the recent large-scale drying out of Siberian fir forests. In zones of outbreak of these beetles, there is a decrease in the productivity of dark coniferous forests [40,41,42].
2.2. Methods
- -
- Products MOD11A1 and MYD11A1, Land surface temperature LST. This parameter correlates well with meteorological observations of air temperature. The observations of the parameter are daily, and spatial resolution is 1 × 1 km.
- -
- Initial red and near infrared spectral channels for calculating the NDVI vegetation index contained in products MOD09Q1 and MYD09Q1. These are an 8-day composite (cleaned and sampled data for a period of 8 days) and the spatial resolution is 250 × 250 m.
2.3. Data Collection
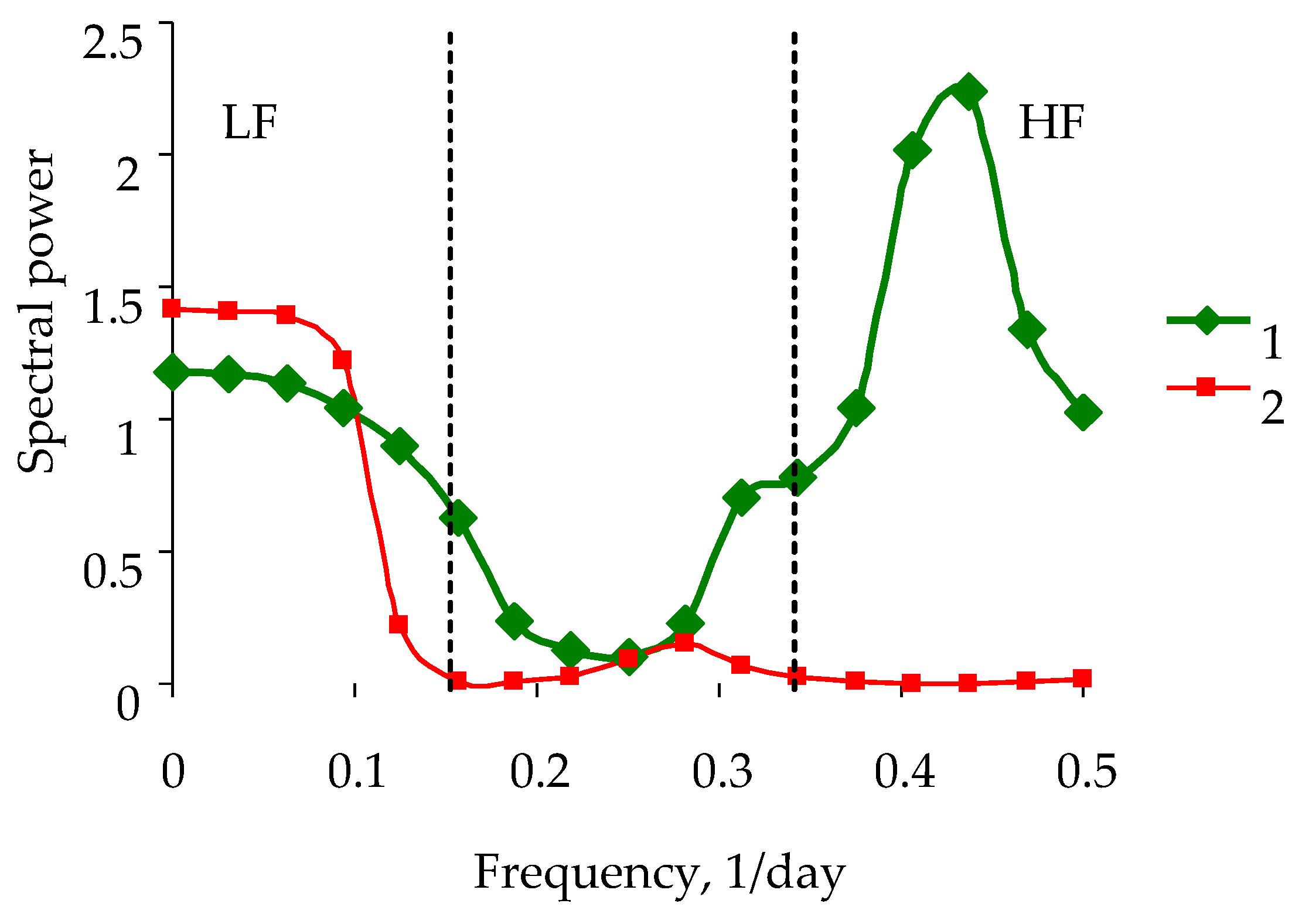
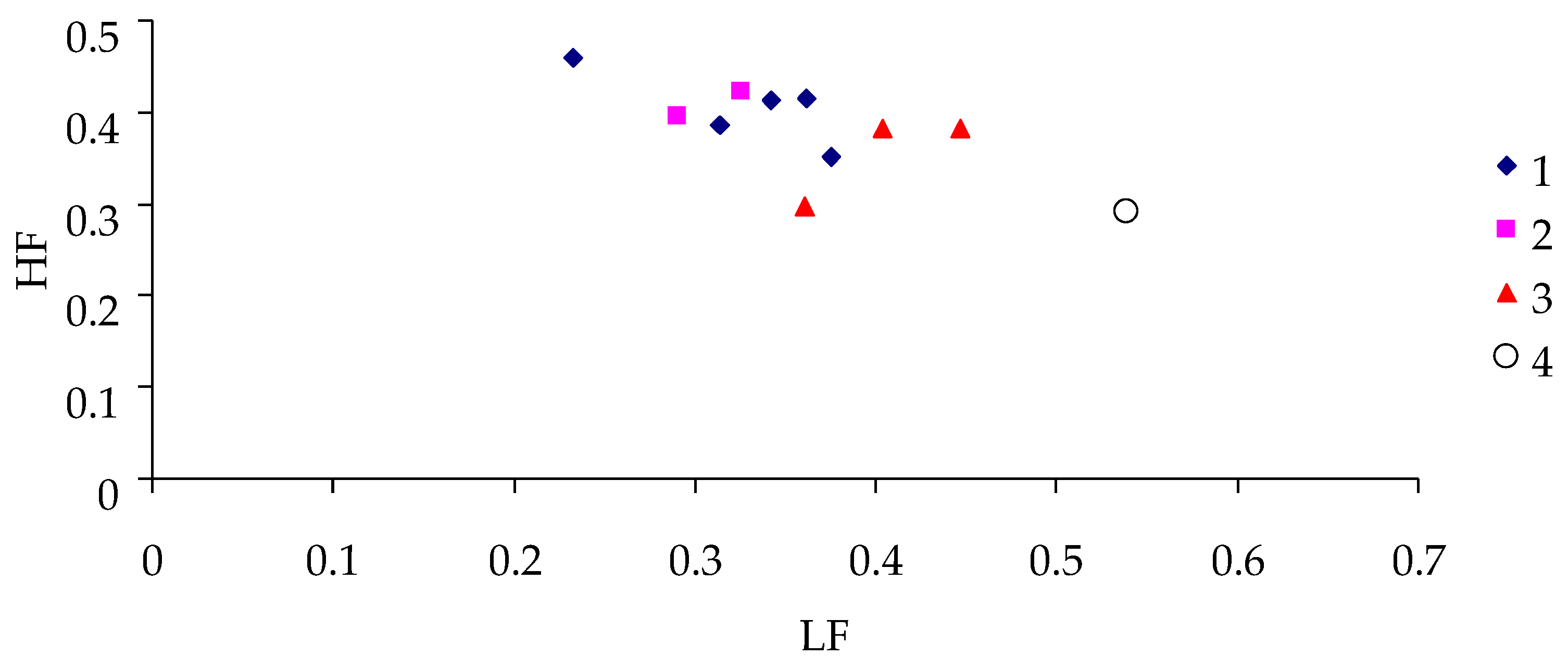
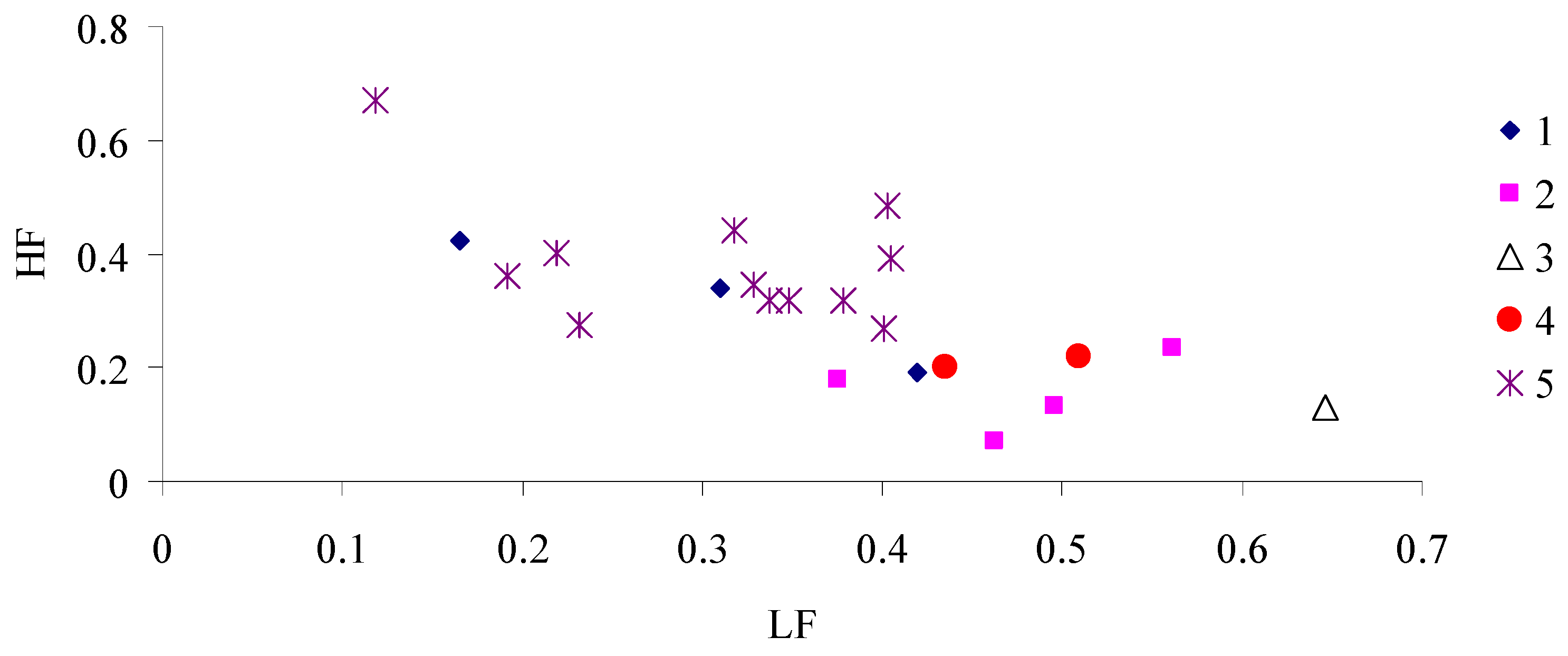
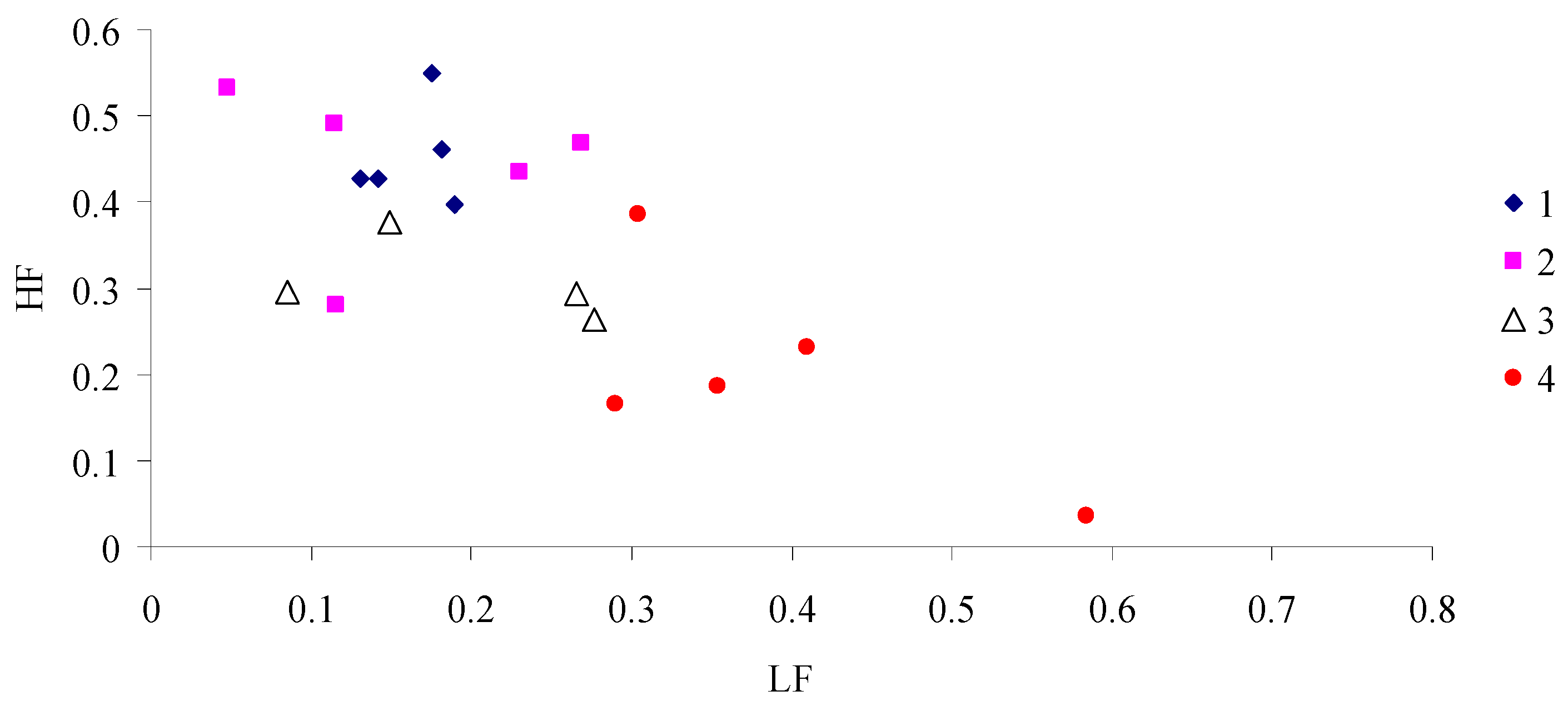
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dyer, L.A.; Carson, W.P.; Leigh, E.G. Insect outbreaks in tropical forests: Patterns, mechanisms, and consequences. In Insect Outbreaks Revisit; Wiley: Hoboken, NJ, USA, 2012; pp. 219–245. [Google Scholar] [CrossRef]
- Kneeshaw, D.; Sturtevant, B.R.; Cooke, B.; Work, T.; Pureswaran, D.; De Grandpre, L.; MacLean, D.A. Insect disturbances in forest ecosystems. In Routledge Handbook of Forest Ecology; Routledge: London, UK, 2015; pp. 93–113. [Google Scholar]
- Silins, I.; Karklina, A.; Miezite, O.; Jansons, A. Trends in Outbreaks of Defoliating Insects Highlight Growing Threats for Central European Forests, and Implications for Eastern Baltic Region. Forests 2021, 12, 799. [Google Scholar] [CrossRef]
- Berner, L.T.; Law, B.E.; Meddens, A.J.H.; Hicke, J.A. Tree mortality from fires, bark beetles, and timber harvest during a hot and dry decade in the western United States (2003–2012). Environ. Res Lett. 2017, 12, 065005. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Brockerhoff, E.G.; Kalisz, S.; Nuñez, M.A.; Wardle, D.A.; Wingfield, M.J. Biological invasions in forest ecosystems. Biol. Invasions 2017, 19, 3437–3458. [Google Scholar] [CrossRef]
- Mezei, P.; Jakuš, R.; Pennerstorfer, J.; Havašová, M.; Škvarenina, J.; Ferenčík, J.; Slivinský, J.; Bičárová, S.; Bilcik, D.; Blazenec, M.; et al. Storms, temperature maxima and the Eurasian spruce bark beetle Ips typographus—An infernal trio in Norway spruce forests of the Central European High Tatra Mountains. Agric. For. Meteorol. 2017, 242, 85–95. [Google Scholar] [CrossRef]
- van Lierop, P.; Lindquist, E.; Sathyapala, S.; Franceschini, G. Global forest area disturbance from fire, insect pests, diseases and severe weather events. For. Ecol. Manag. 2015, 352, 78–88. [Google Scholar] [CrossRef]
- Bentz, B.; Bonello, P.; Delb, H.; Fettig, C.; Poland, T.; Pureswaran, D.; Seybold, S. Advances in understanding and managing insect pests of forest trees. In Achieving Sustainable Management of Boreal and Temperate Forests; Stanturf, J., Ed.; Burleigh Dodds Science Publishing Ltd.: Cambridge, UK, 2019; pp. 515–585. [Google Scholar]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negron, J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest insects and climate change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef]
- Candau, J.N.; Fleming, R.A. Forecasting the response of spruce budworm defoliation to climate change in Ontario. Can. J. For. Res. 2011, 41, 1948–1960. [Google Scholar] [CrossRef]
- Godefroid, M.; Meurisse, N.; Groenen, F.; Kerdelhué, C.; Rossi, J.P. Current and future distribution of the invasive oak processionary moth. Biol. Invasions 2020, 22, 523–534. [Google Scholar] [CrossRef]
- Björkman, C.; Bylund, H.; Nilsson, U.; Nordlander, G.; Schroeder, M. Forest management to mitigate insect damage in a changing climate: Possibilities and uncertainties. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CABI: Wallingford, CT, USA, 2015; pp. 248–266. [Google Scholar]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef]
- Ren, P.; Neron, V.; Rossi, S.; Liang, E.; Bouchard, M.; Deslauriers, A. Warming counteracts defoliation-induced mismatch by increasing herbivore-plant phenological synchrony. Glob. Chang. Biol. 2020, 26, 2072–2080. [Google Scholar] [CrossRef]
- Liu, E.Y.; Lantz, V.A.; MacLean, D.A.; Hennigar, C. Economics of early intervention to suppress a potential spruce budworm outbreak on crown land in New Brunswick, Canada. Forests 2019, 10, 481. [Google Scholar] [CrossRef]
- Jactel, H.; Brockerhoff, E.; Duelli, P. A test of the biodiversity-stability theory: Meta-analysis of tree species diversity effects on insect pest infestations, and re-examination of responsible factors. In Forest Diversity and Function: Temperate and Boreal Systems; Scherer-Lorenzen, M., Korner, C., Schulze, E.D., Eds.; Springer: Berlin, Germany, 2005; pp. 235–262. [Google Scholar]
- Jactel, H.; Brockerhoff, E.G. Tree diversity reduces herbivory by forest insects. Ecol. Lett. 2007, 10, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Klapwijk, M.J.; Bylund, H.; Schroeder, M.; Björkman, C. Forest management and natural biocontrol of insect pests. Forestry 2016, 89, 253–262. [Google Scholar] [CrossRef]
- MacLean, D.A.; Erdle, T.A.; MacKinnon, W.E.; Porter, K.B.; Beaton, K.P.; Cormier, G.; Morehouse, S.; Budd, M. The spruce budworm decision support system: Forest protection planning to sustain long-term wood supply. Can. J. For. Res. 2001, 31, 1742–1757. [Google Scholar] [CrossRef]
- Dymond, C.C.; Wulder, M.A.; Shore, T.L.; Nelson, T.; Boots, B.; Riel, B.G. Evaluation of risk assessment of mountain pine beetle infestations. West. J. Appl. For. 2006, 21, 5–13. [Google Scholar] [CrossRef]
- Tucker, C.J.; Sellers, P.J. Satellite remote sensing of primary production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar] [CrossRef]
- Liu, Y.; Hill, M.J.; Zhang, X.; Wang, Z.; Richardson, A.D.; Hufkens, K.; Filippa, G.; Baldocchi, D.D.; Ma, S.; Verfaillie, J.; et al. Using data from Landsat, MODIS, VIIRS and Pheno Cams to monitor the phenology of California oak/grass savanna and open grassland across spatial scales. Agric. For. Meteorol. 2017, 237–238, 311–325. [Google Scholar] [CrossRef]
- Rechid, D.; Raddatz, T.J.; Jacob, D. Parameterization of snow-free land surfacealbedo as a function of vegetation phenology based on MODIS data and appliedin climate modelling. Theor. Appl. Climatol. 2009, 95, 245–255. [Google Scholar] [CrossRef]
- Bayarjargal, Y.; Karnieli, A.; Bayasgalan, M.; Khudulmur, S.; Gandush, C.; Tucker, C.J. A comparative study of NOAA-AVHRR derived drought indices using change vector analysis. Int. J. Remote Sens. 2006, 105, 9–22. [Google Scholar] [CrossRef]
- Cunha, M.; Richter, C. A time-frequency analysis on the impact of climate variability with focus on semi-natural montane grassland meadows. IEEE Trans. Geosci. Remote Sens. 2014, 52, 6156–6164. [Google Scholar] [CrossRef]
- Jacquin, A.; Sheeren, D.; Lacombe, J.P. Vegetation cover degradation assessment in Madagascar savanna based on trend analysis of MODIS NDVI time series. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, S3–S10. [Google Scholar] [CrossRef]
- Liang, L.; Chen, Y.; Hawbaker, T.; Zhu, Z.; Gong, P. Mapping mountain pine beetle mortality through growth trend analysis of time-series landsat data. Remote Sens. 2014, 6, 5696–5716. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef]
- Verbesselt, J.; Zeileis, A.; Herold, M. Near real-time disturbance detection using satellite image time series. Remote Sens. Environ. 2012, 123, 98–108. [Google Scholar] [CrossRef]
- Spruce, J.P.; Sader, S.; Ryan, R.E.; Smoot, J.; Kuper, P.; Ross, K.; Prados, D.; Russell, J.; Gasser, G.; McKellip, R. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks. Remote Sens. Environ. 2011, 115, 427–437. [Google Scholar] [CrossRef]
- Thayn, J.B. Using a remotely sensed optimized Disturbance Index to detect insect defoliation in the Apostle Islands, Wisconsin, USA. Remote Sens. Environ. 2013, 136, 210–217. [Google Scholar] [CrossRef]
- Olsson, P.O.; Lindstrom, J.; Eldundh, L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI. Remote Sens. Environ. 2016, 181, 42–53. [Google Scholar] [CrossRef]
- Rozhkov, A.S. Siberian Silk Moth; Nauka: Moscow, Russia, 1963; 175p. (In Russian) [Google Scholar]
- Kondakov, Y.P. Patterns of the Siberian silkmoth outbreaks. In Ecology of Populations of Forest Animals of Siberia; Nauka Publ.: Novosibirsk, Russia, 1974; pp. 206–265. (In Russian) [Google Scholar]
- Isaev, A.S.; Khlebopros, R.G.; Kiselev, V.V.; Kondakov, Y.P.; Nedorezov, L.V.; Soukhovolsky, V.G. Forest Insects Population Dynamics; Publishing House of the Eurasian Entomological Journal: Novosibirsk, Russia, 2009; 115p. [Google Scholar]
- Isaev, A.S.; Girs, G.I. Interaction between Tree and Xylophagous Insects; Nauka Publ.: Novosibirsk, Russia, 1975; 344p. (In Russian) [Google Scholar]
- Isaev, A.S.; Rozhkov, A.S.; Kiselev, V.V. Black Fir Longhorned Beetle Monochamus Urussovi (Fisch.); Nauka Publ.: Novosibirsk, Russia, 1988; 271p, ISBN 5-02-029406-3. (In Russian) [Google Scholar]
- Kerchev, I.A. Ecology of the Ussuri polygraph Polygraphus proximus Blandf. (Coleoptera: Curculionidae, Scolytinae) in the West Siberian region of invasion. Russ. J. Biol. Invasions 2014, 2, 80–94. (In Russian) [Google Scholar]
- Krivets, S.A.; Bisirova, E.M.; Kerchev, I.A.; Pats, E.N.; Chernova, N.A. Population characteristics and influence of the Ussuri polygraph on the state of fir forests of North-Eastern Altai. Proc. News St. Petersburg For. Acad. 2014, 207, 37–48. (In Russian) [Google Scholar]
- Krivets, S.A.; Kerchev, I.A.; Bisirova, E.M.; Debkov, N.M. Modern distribution and forecast of expansion of the invasive range of the Ussuri Polygraphus proximus Blandford, 1894 in the Tomsk region (Western Siberia). Eurasian Entomol. J. 2018, 17, 53–60. (In Russian) [Google Scholar] [CrossRef]
- Public Database of MODIS Satellite Systems. Available online: http://modis.gsfc.nasa.gov (accessed on 10 May 2024).
- Cochran, J.A. The Analysis of Linear Integral Equations; McGraw-Hill Book Co.: New York, NY, USA, 1972; 370p. [Google Scholar]
- Jerry, A.J. Introduction to Integral Equations with Applications; Marcel Dekker: New York, NY, USA; Basel, Switzerland, 1985; 254p. [Google Scholar]
- Hackbusch, W. Integral Equations: Theory and Numerical Treatment; Birkhäuser Verlag: Boston, MA, USA, 1995; 362p. [Google Scholar]
- Polyanin, A.D.; Manzhirov, A.V. Handbook of Integral Equations; Chapman and Hall/CRC: Boca Raton, FL, USA, 2008; 1081p. [Google Scholar]
- Wei, W.W.S. Time Series Analysis; Addison Wesley: Boston, MA, USA, 2008; 614p. [Google Scholar]
- Application for Extracting and Exploring Analysis Ready Samples (AppEEARS). Available online: https://lpdaacsvc.cr.usgs.gov/appeears/ (accessed on 9 May 2024).
- Meddens, A.J.H.; Hicke, J.A.; Vierling, L.A.; Hudak, A.T. Evaluating methods to detect bark beetle-caused tree mortality using single-date and multi-date Landsat imagery. Remote Sens. Environ. 2013, 132, 49–58. [Google Scholar] [CrossRef]
- Bright, B.C.; Hudak, A.T.; Meddens, A.J.H.; Egan, J.M.; Jorgensen, C.L. Mapping Multiple Insect Outbreaks across Large Regions Annually Using Landsat Time Series Data. Remote Sens. 2020, 12, 1655. [Google Scholar] [CrossRef]
- Ganz, S.; Adler, P.; Kändler, G. Forest cover mapping based on a combination of aerial images and Sentinel-2 satellite data compared to National Forest Inventory data. Forests 2020, 11, 1322. [Google Scholar] [CrossRef]
- Abdollahnejad, A.; Panagiotidis, D.; Surový, P.; Modlinger, R. Investigating the Correlation between Multisource Remote Sensing Data for Predicting Potential Spread of Ips typographus L. Spots in Healthy Trees. Remote Sens. 2021, 13, 4953. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovalev, A.; Tarasova, O.; Soukhovolsky, V.; Ivanova, Y. Is It Possible to Predict a Forest Insect Outbreak? Backtesting Using Remote Sensing Data. Forests 2024, 15, 1458. https://doi.org/10.3390/f15081458
Kovalev A, Tarasova O, Soukhovolsky V, Ivanova Y. Is It Possible to Predict a Forest Insect Outbreak? Backtesting Using Remote Sensing Data. Forests. 2024; 15(8):1458. https://doi.org/10.3390/f15081458
Chicago/Turabian StyleKovalev, Anton, Olga Tarasova, Vladislav Soukhovolsky, and Yulia Ivanova. 2024. "Is It Possible to Predict a Forest Insect Outbreak? Backtesting Using Remote Sensing Data" Forests 15, no. 8: 1458. https://doi.org/10.3390/f15081458