Spatial Pattern of Drought-Induced Mortality Risk and Influencing Factors for Robinia pseudoacacia L. Plantations on the Chinese Loess Plateau

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
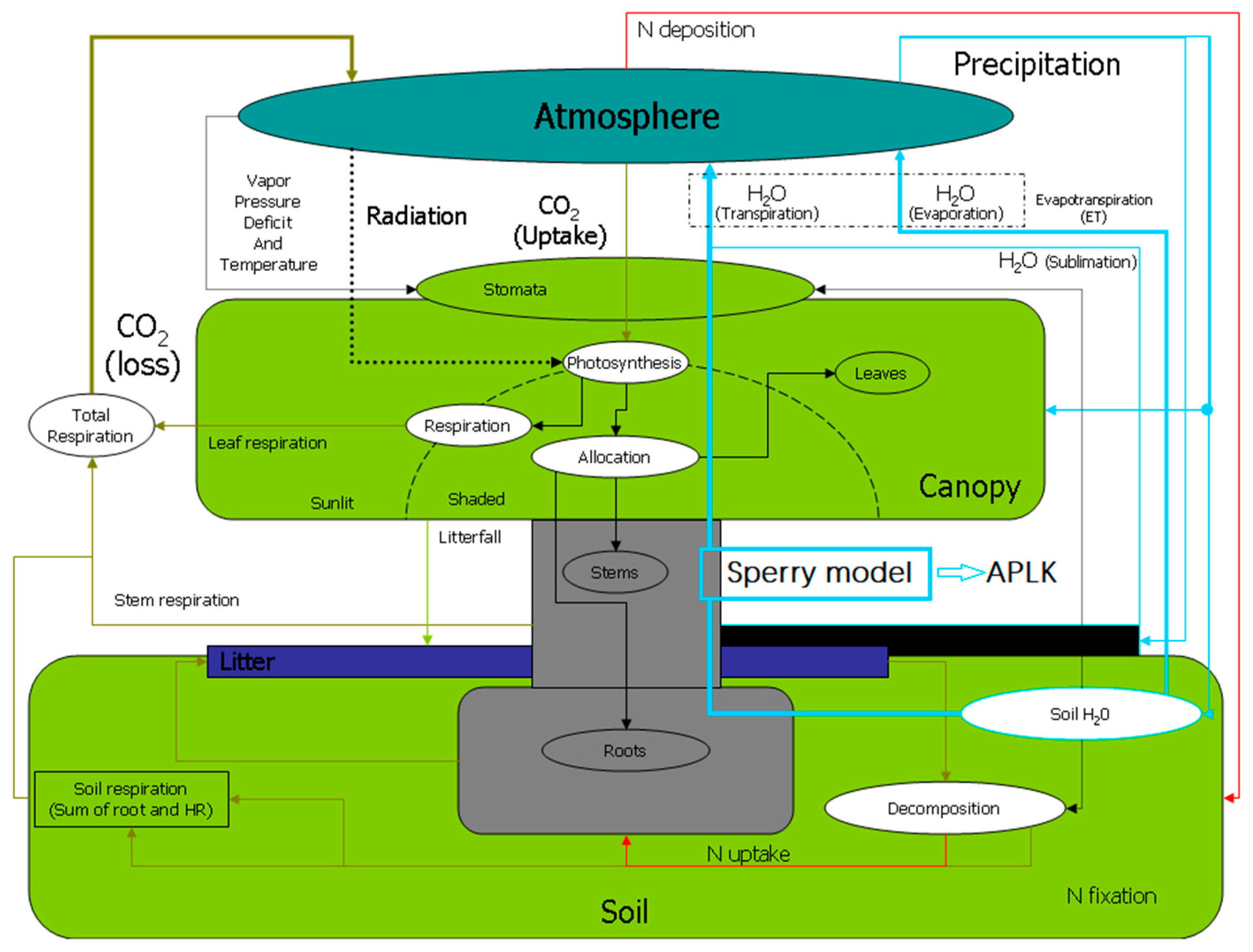
2.2. BBGC-SPERRY Model Description

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| (a) Biome-BGC model | |
| Transfer growth period fraction of growing season (%) | 0.2 |
| Litterfall fraction of growing season (%) | 0.2 |
| Annual leaf and fine root turnover fraction (yr−1) | 1.00 |
| Annual live wood turnover fraction (yr−1) | 0.7 |
| Allocation new fine root C:new leaf C | 1.0 |
| Allocation new stem C:new leaf C | 2.20 |
| Allocation new live wood C:new total wood C | 0.209 [40] |
| Allocation new root C:new stem C | 0.22 |
| Allocation current growth proportion (%) | 0.5 |
| C:N of leaves | 18.8 [45] |
| C:N of leaf litter | 32.2 [46] |
| C:N of fine roots | 26.7 [45] |
| C:N of live wood | 50 |
| C:N of dead wood | 550 |
| Leaf litter labile proportion | 0.35 |
| Leaf litter cellulose proportion | 0.40 |
| Leaf litter lignin proportion | 0.26 [45] |
| Fine root labile proportion | 0.34 |
| Fine root cellulose proportion | 0.44 |
| Fine root lignin proportion | 0.22 |
| Dead wood cellulose proportion | 0.68 |
| Dead wood lignin proportion | 0.32 |
| Canopy water interception coefficient (LAI−1 d−1) | 0.045 |
| Canopy light extinction coefficient | 0.54 |
| All-sided to projected leaf area ratio | 2.0 |
| Specific leaf area (m2 kg−1 C) | 27.9 [45] |
| Ratio of shaded SLA:sunlit SLA | 1.3 |
| Fraction of leaf N in Rubisco | 0.14 |
| Cuticular conductance (m s−1) | 0.00006 |
| Boundary layer conductance (m s−1) | 0.01 |
| (b) Sperry model | |
| Weibull function b and c for root, stem, leaf | b = 2.6, c = 4.1 [33] |
| Maximum whole-plant hydraulic conductance per leaf area (mmol s−1 m−2 MPa−1) | 4.2 |
| Maximum diffusive conductance (mol s−1 m−2) | 0.1 |
| Average % resistance in rhizosphere (%) | 20.5 |
| Root depth coefficient | 0.99 [47] |
| Number of root and soil layers | 5 |
2.3. Model Evaluation
2.4. Assessment of Drought-Induced Mortality Risk
2.5. Statistical Analyses
3. Results
3.1. Model Evaluation and Uncertainty Analysis
3.2. Spatial Pattern of Plant Hydraulic Safety
3.3. Spatial Pattern of Drought-Induced Mortality Risk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heinrich, V.H.A.; Vancutsem, C.; Dalagnol, R.; Rosan, T.M.; Fawcett, D.; Silva-Junior, C.H.L.; Cassol, H.L.G.; Achard, F.; Jucker, T.; Silva, C.A.; et al. The carbon sink of secondary and degraded humid tropical forests. Nature 2023, 615, 436–442. [Google Scholar] [CrossRef]
- Piao, S.L.; Wang, X.H.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.; Ciais, P.; Mmervik, H.T.; et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 2020, 1, 14–27. [Google Scholar] [CrossRef]
- Peng, J.; Hu, T.; Qiu, S.J.; Hu, Y.N.; Dong, J.Q.; Lin, Y.F. Balancing the effects of forest conservation and restoration on south China Karst greening. Earth’s Future 2023, 11, e2023EF003487. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Konings, A.G.; Trugman, A.T.; Yu, K.; Bowling, D.R.; Gabbitas, R.; Karp, D.S.; Pacala, S.; Sperry, J.S.; Sulman, B.N.; et al. Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature 2018, 561, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Tavares, J.V.; Oliveira, R.S.; Mencuccini, M.; Signori-Müller, C.; Pereira, L.; Diniz, F.C.; Gilpin, M.; Zevallos, M.M.J.; Yupayccana, C.S.; Acosta, M.; et al. Basin-wide variation in tree hydraulic safety margins predicts the carbon balance of Amazon forests. Nature 2023, 617, 111–117. [Google Scholar] [CrossRef]
- Bauman, D.; Fortunel, C.; Delhaye, G.; Malhi, Y.; Cernusak, L.A.; Bentley, L.P.; Rifai, S.W.; Aguirre-Gutiérrez, J.; Menor, I.O.; Phillips, O.L.; et al. Tropical tree mortality has increased with rising atmospheric water stress. Nature 2022, 608, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; Lopez, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Gao, Y.Q.; Chen, Z.T.; Chen, J.X.; Yang, L.; Li, J.Y.; He, Q.; Qiu, Q.; Su, Y. A bibliometric analysis of the mechanisms underlying drought-induced tree mortality. Forests 2024, 15, 1037. [Google Scholar] [CrossRef]
- Zhao, A.Z.; Liu, X.F.; Zhu, X.F.; Pan, Y.Z.; Li, Y.Z. Spatiotemporal patterns of droughts based on SWAT model for the Weihe River Basin. Prog. Geogr. 2015, 34, 1156–1166. [Google Scholar]
- Wang, L.N.; Zhu, Q.K.; Yan, D.H.; Meng, L.I. The assessment on drought risk of Pinus tabuliformis plantations in the Loess Plateau. South-North Water Transf. Water Sci. Technol. 2017, 15, 179–195. [Google Scholar]
- Tai, X.N.; Mackay, D.S.; Anderegg, W.R.L.; Sperry, J.S.; Brooks, P.D. Plant hydraulics improves and topography mediates prediction of aspen mortality in southwestern USA. New Phytol. 2017, 213, 113–127. [Google Scholar] [CrossRef]
- Li, L.C.; Yang, Z.L.; Matheny, A.M.; Zheng, H.; Swenson, S.C.; Lawrence, D.M.; Barlage, M.; Yan, B.Y.; McDowell, N.G.; Leung, L.R. Representation of plant hydraulics in the Noah-MP land surface model: Model development and multiscale evaluation. J. Adv. Model. Earth Syst. 2021, 13, e2020MS002214. [Google Scholar] [CrossRef]
- Kennedy, D.; Swenson, S.; Oleson, K.W.; Lawrence, D.M.; Fisher, R.; da Costa, A.C.L.; Gentine, P. Implementing plant hydraulics in the community land model, version 5. J. Adv. Model. Earth Syst. 2019, 11, 485–513. [Google Scholar] [CrossRef]
- McDowell, N.; Allen, C.D.; Anderson-Teixeira, K.; Brando, P.; Brienen, R.; Chambers, J.; Christoffersen, B.; Davies, S.; Doughty, C.; Duque, A.; et al. Drivers and mechanisms of tree mortality in moist tropical forests. New Phytol. 2018, 219, 851–869. [Google Scholar] [CrossRef]
- Trugman, A.T. Integrating plant physiology and community ecology across scales through trait-based models to predict drought mortality. New Phytol. 2022, 234, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Simeone, C.; Maneta, M.P.; Holden, Z.A.; Sapes, G.; Sala, A.; Dobrowski, S.Z. Coupled ecohydrology and plant hydraulics modeling predicts ponderosa pine seedling mortality and lower treeline in the US Northern Rocky Mountains. New Phytol. 2019, 221, 1814–1830. [Google Scholar] [CrossRef]
- O’Grady, A.P.; Mitchell, P.J.M.; Pinkard, E.A.; Tissue, D.T. Thirsty roots and hungry leaves: Unravelling the roles of carbon and water dynamics in tree mortality. New Phytol. 2013, 200, 294–297. [Google Scholar] [CrossRef]
- Doughty, C.E.; Metcalfe, D.B.; Girardin, C.A.J.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, M.C.; Rocha, W.; et al. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Delzon, S.; Cochard, H. Recent advances in tree hydraulics highlight the ecological significance of the hydraulic safety margin. New Phytol. 2014, 203, 355–358. [Google Scholar] [CrossRef]
- Brodribb, T.; Brodersen, C.R.; Carriqui, M.; Tonet, V.; Dominguez, C.R.; McAdam, S. Linking xylem network failure with leaf tissue death. New Phytol. 2021, 232, 68–79. [Google Scholar] [CrossRef]
- Yan, X.Y.; Zhang, Z.D.; Zhao, X.F.; Huang, M.B.; Wu, X.F.; Guo, T.Q. Differentiated responses of plant water use regulation to drought in Robinia pseudoacacia plantations on the Chinese Loess Plateau. Agric. Water Manag. 2024, 291, 108659108659. [Google Scholar] [CrossRef]
- Hu, Y.W.; Zhao, J.C.; Li, Y.; Tang, P.; Yang, Z.; Zhang, J.J.; Sun, R.X. Biomass and carbon stock capacity of Robinia pseudoacacia plantations at different densities on the Loess Plateau. Forests 2024, 15, 1242. [Google Scholar] [CrossRef]
- Ma, C.K.; Luo, Y.; Shao, M.A. Comparative modeling of the effect of thinning on canopy interception loss in a semiarid black locust (Robinia pseudoacacia) plantation in Northwest China. J. Hydrol. 2020, 590, 125234. [Google Scholar] [CrossRef]
- Xu, M.X.; Liu, G.B. The characteristics and evolution of soil nutrient in artifcial black locust (Robinia pseudoacacia) forest-land in the hilly Loess Plateau. Plant Nutr. Fertil. Sci. 2004, 10, 40–46. [Google Scholar]
- Jia, X.X.; Shao, M.A.; Zhu, Y.J.; Luo, Y. Soil moisture decline due to afforestation across the Loess Plateau, China. J. Hydrol. 2017, 546, 113–122. [Google Scholar] [CrossRef]
- Shao, M.A.; Wang, Y.Q.; Xia, Y.Q.; Jia, X.X. Soil drought and water carrying capacity for vegetation in the critical zone of the Loess Plateau: A review. Vadose Zone J. 2018, 17, 170077. [Google Scholar] [CrossRef]
- Xiang, W.; Si, B.C.; Li, H.J.; Li, M.; Song, J.X.; Tian, Y.L. Impacts of Deep-Rooted Apple Tree on Soil Water Balance in the Semi-Arid Loess Plateau, China. Forests 2024, 15, 930. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Huang, M.B. Effect of root-zone vertical soil moisture heterogeneity on water transport safety in soil-plant-atmosphere continuum in Robinia pseudoacacia. Agric. Water Manag. 2021, 246, 106702. [Google Scholar] [CrossRef]
- Wei, J.S.; Li, Z.S.; Feng, X.Y.; Zhang, Y.; Chen, W.L.; Xing, W.U.; Jiao, L.; Wang, X.C. Ecological and physiological mechanisms of growth decline of Robinia pseudoacacia plantations in the Loess Plateau of China: A review. Chin. J. Appl. Ecol. 2018, 29, 2433–2444. [Google Scholar]
- Liu, J.H. Study on Growth Traits of Robinia pseudoacacia Plantation and Its Naturalization Process on Loess Plateau. Ph.D. Thesis, Graduate University of Chinese Academy of Sciences, Beijing, China, 2008. [Google Scholar]
- Jia, H.S.; Guan, C.F.; Zhang, J.S.; He, C.X.; Yin, C.J.; Meng, P. Drought effects on tree growth, water use efficiency, vulnerability and canopy health of Quercus variabilis-Robinia pseudoacacia mixed plantation. Front. Plant Sci. 2022, 13, 1018405. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Huang, M.B.; Yang, Y.N.; Zhao, X.F. Evaluating drought-induced mortality risk for Robinia pseudoacacia plantations along the precipitation gradient on the Chinese Loess Plateau. Agric. For. Meteorol. 2020, 284, 107897. [Google Scholar] [CrossRef]
- Sperry, J.S.; Wang, Y.J.; Wolfe, B.T.; Mackay, D.S.; Anderegg, W.R.L.; McDowell, N.G.; Pockman, W.T. Pragmatic hydraulic theory predicts stomatal responses to climatic water deficits. New Phytol. 2016, 212, 577–589. [Google Scholar] [CrossRef]
- Yan, X.Y.; Zhang, Z.D.; Huang, M.B.; Zhao, X.F.; Yang, F.; Wu, X.F. The impact of climate change on growth and drought-induced mortality risk of Robinia pseudoacacia plantations along a precipitation gradient on the Chinese Loess Plateau. Agric. For. Meteorol. 2022, 325, 109160. [Google Scholar] [CrossRef]
- Yang, W.Z.; Ma, Y.X.; Han, S.F.; Yang, X.M. Water ecological regionalization of afforestation in Loess Plateau. J. Soil. Water Conserv. 1994, 8, 1–9. [Google Scholar]
- Wang, Y.Q.; Hu, W.; Sun, H.; Zhao, Y.; Zhang, P.P.; Li, Z.M.; Zhou, Z.X.; Tong, Y.P.; Liu, S.Z.; Zhou, J.X.; et al. Soil moisture decline in China’s monsoon loess critical zone: More a result of land-use conversion than climate change. Proc. Natl. Acad. Sci. USA 2024, 121, e2322127121. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Jia, X.X.; Shao, M.A. Loess thickness variations across the Loess Plateau of China. Surv. Geophys. 2018, 39, 715–727. [Google Scholar] [CrossRef]
- Jia, X.X.; Shao, M.A.; Yu, D.X.; Zhang, Y.; Binley, A. Spatial variations in soil-water carrying capacity of three typical revegetation species on the Loess Plateau, China. Agric. Ecosyst. Environ. 2019, 273, 25–35. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and sensitivity analysis of the Biome–BGC terrestrial ecosystem model: Net primary production controls. Earth Interact. 2000, 4, 1–85. [Google Scholar] [CrossRef]
- Thornton, P.E.; Law, B.E.; Gholz, H.L.; Clark, K.L.; Falge, E.; Ellsworth, D.S.; Goldstein, A.H.; Monson, R.K.; Hollinger, D.; Falk, M.; et al. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agric. For. Meteorol. 2002, 113, 185–222. [Google Scholar] [CrossRef]
- Thornton, P.E.; Rosenbloom, N.A. Ecosystem model spin-up: Estimating steady state conditions in a coupled terrestrial carbon and nitrogen cycle model. Ecol. Model. 2005, 189, 25–48. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Monteith, J.L. Evaporation and environment. Symp. Soc. Exp. Biol. 1965, 19, 205–234. [Google Scholar]
- Ma, R.T.; An, S.S.; Huang, Y.M. C, N and P stoichiometry characteristics of different-aged Robinia pseudoacacia plantations on the Loess Plateau, China. Chin. J. Appl. Ecol. 2017, 28, 2787–2793. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M. Nitrogen immobilization in decaying hardwood leaf litter as a function of initial nitrogen and lignin content. Can. J. Bot. 1982, 60, 2263–2269. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Huang, M.B.; Zhang, Y.K. Vertical distribution of fine-root area in relation to stand age and environmental factors in black locust (Robinia pseudoacacia) forests of the Chinese Loess Plateau. Can. J. For. Res. 2018, 48, 1148–1158. [Google Scholar] [CrossRef]
- Huang, M.B.; Zettl, J.D.; Barbour, S.L.; Elshorbagy, A.; Si, B.C. The impact of soil moisture availability on forest growth indices for variably layered coarse-textured soils. Ecohydrology 2013, 6, 214–227. [Google Scholar] [CrossRef]
- Wang, Y.L. Characteristics of Photosynthesis and Water Use of Dominant Tree Species in Two Typical Forests and Related Factors in the Semiarid Region of Loess Plateau, China. Ph.D. Thesis, Graduate University of Chinese Academy of Sciences, Beijing, China, 2010. [Google Scholar]
- Arya, L.M.; Leij, F.J.; van Genuchten, M.T.; Shouse, P.J. Scaling parameter to predict the soil water characteristic from particle-size distribution data. Soil Sci. Soc. Am. J. 1999, 63, 510–519. [Google Scholar] [CrossRef]
- van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- Niu, L.T.; Jia, X.X.; Li, X.D.; Zhao, C.L.; Ren, L.D.; Hu, W.; Zhu, P.; Li, D.F.; Zhang, B.Q.; Shao, M.A. Developing novel ensemble models for predicting soil hydraulic properties in China’s arid region. J. Hydrol. 2024, 636, 131354. [Google Scholar] [CrossRef]
- McDowell, N.G.; Fisher, R.A.; Xu, C.G.; Domec, J.C.; Hölttä, T.; Mackay, D.S.; Sperry, J.S.; Boutz, A.; Dickman, L.; Gehres, N.; et al. Evaluating theories of drought-induced vegetation mortality using a multimodel–experiment framework. New Phytol. 2013, 200, 304–321. [Google Scholar] [CrossRef]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Wang, R.Q.; Zhang, L.L.; Zhang, S.X.; Cai, J.; Tyree, M.T. Water relations of Robinia pseudoacacia L.: Do vessels cavitate and refill diurnally or are R-shaped curves invalid in Robinia? Plant Cell Environ. 2014, 37, 2667–2678. [Google Scholar] [CrossRef]
- Sperry, J.S.; Love, D.M. What plant hydraulics can tell us about responses to climate-change droughts. New Phytol. 2015, 207, 14–27. [Google Scholar] [CrossRef]
- Tai, X.N.; Mackay, D.S.; Sperry, J.S.; Brooks, P.; Anderegg, W.R.L.; Flanagan, L.B.; Rood, S.B.; Hopkinson, C. Distributed plant hydraulic and hydrological modeling to understand the susceptibility of riparian woodland trees to drought-induced mortality. Water Resour. Res. 2018, 54, 4901–4915. [Google Scholar] [CrossRef]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models part I—A discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Poyatos, R.; Granda, V.; Molowny-Horas, R.; Mencuccini, M.; Steppe, K.; Martínez-Vilalta, J. SAPFLUXNET: Towards a global database of sap flow measurements. Tree Physiol. 2016, 36, 1449–1455. [Google Scholar] [CrossRef]
- Beven, K. On doing better hydrological science. Hydrol. Process. 2008, 22, 3549–3553. [Google Scholar] [CrossRef]
- Bai, X.; Jia, X.X.; Jia, Y.H.; Shao, M.A.; Hu, W. Modeling long-term soil water dynamics in response to land-use change in a semi-arid area. J. Hydrol. 2020, 585, 124824. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Jia, X.X.; Zhu, P.; Huang, M.B.; Ren, L.D.; Shao, M.A. Estimating the optimal vegetation coverage for the dominant tree and shrub species over China’s northwest drylands. Sci. China Earth Sci. 2024, 67, 1500–1517. [Google Scholar] [CrossRef]
- Li, Y.; Shao, M.A.; Wang, W.Y.; Wang, Q.J. Sensitive analyses of parameters in unsaturated soil hydraulic conductivity model. Adv. Water Sci. 2003, 14, 593–597. [Google Scholar]
- Li, J.K.; Zhao, R.S.; Li, Y.J.; Chen, L. Modeling the effects of parameter optimization on three bioretention tanks using the HYDRUS-1D model. J. Environ. Manag. 2018, 217, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.B.; Fredlund, D.G.; Fredlund, M.D. Comparison of Measured and PTF Predictions of SWCCs for Loess Soils in China. Geotech. Geol. Eng. 2010, 28, 105–117. [Google Scholar] [CrossRef]
- da Silva, A.C.; Armindo, R.A.; dos Santos Brito, A.; Schaap, M.G. SPLINTEX: A physically-based pedotransfer function for modeling soil hydraulic functions. Soil Tillage Res. 2017, 174, 261–272. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, M.B.; Lian, J.J. Spatial distributions of optimal plant coverage for the dominant tree and shrub species along a precipitation gradient on the central Loess Plateau. Agric. For. Meteorol. 2015, 206, 69–84. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Huang, M.B.; Zhao, X.F.; Wu, L.H. Adjustments of leaf traits and whole plant leaf area for balancing water supply and demand in Robinia pseudoacacia under different precipitation conditions on the Loess Plateau. Agric. For. Meteorol. 2019, 279, 107733. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Piao, S.L.; Myneni, R.B.; Huang, M.T.; Zeng, Z.Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, W.; Li, S.D.; Xie, B.N.; Shi, F.Z.; Zhao, J.X. Spatial distribution of optimal plant cover and its influencing factors for Populus simonii carr. on the Bashang Plateau, China. Land 2022, 11, 890. [Google Scholar] [CrossRef]
- Feki, M.; Ravazzani, G.; Pellicone, G.; Caloiero, T. Impact of forest management on wood production under climate change in the Bonis catchment. Forests 2024, 15, 539. [Google Scholar] [CrossRef]
- Hidy, D.; Barcza, Z.; Hollós, R.; Dobor, L.; Acs, T.; Zacháry, D.; Filep, T.; Pásztor, L.; Incze, D.; Dencso, M.; et al. Soil-related developments of the Biome-BGCMuSo v6.2 terrestrial ecosystem model. Geosci. Model. Dev. 2022, 15, 2157–2181. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.E.N.; Peckham, S.D.; Gower, S.T.; Ewers, B.E. Effects of fire on regional evapotranspiration in the central Canadian boreal forest. Glob. Chang. Biol. 2009, 15, 1242–1254. [Google Scholar] [CrossRef]
- Gharsallah, O.; Facchi, A.; Gandolfi, C. Comparison of six evapotranspiration models for a surface irrigated maize agro-ecosystem in Northern Italy. Agric. Water Manag. 2013, 130, 119–130. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Cochard, H.; Delzon, S.; Boivin, T.; Burlett, R.; Cailleret, M.; Corso, D.; Delmas, C.E.L.; De Caceres, M.; Diaz-Espejo, A.; et al. Plant hydraulics at the heart of plant, crops and ecosystem functions in the face of climate change. New Phytol. 2024, 241, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Lu, X.J.; Wei, Z.W.; Zhu, S.G.; Wei, N.; Zhang, S.P.; Yuan, H.; Shang Guan, W.; Liu, S.F.; Zhang, S.L.; et al. New Representation of Plant Hydraulics Improves the Estimates of Transpiration in Land Surface Model. Forests 2021, 12, 722. [Google Scholar] [CrossRef]
- Joshi, J.; Stocker, B.D.; Hofhansl, F.; Zhou, S.; Dieckmann, U.; Prentice, I.C. Towards a unified theory of plant photosynthesis and hydraulics. Nat. Plants 2022, 8, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Sperry, J.S.; Venturas, M.D.; Todd, H.N.; Trugman, A.T.; Anderegg, W.R.L.; Wang, Y.; Tai, X. The impact of rising CO2 and acclimation on the response of US forests to global warming. Proc. Natl. Acad. Sci. USA 2019, 116, 25734–25744. [Google Scholar] [CrossRef]
- Potkay, A.; Trugman, A.T.; Wang, Y.; Venturas, M.D.; Anderegg, W.R.L.; Mattos, C.R.C.; Fan, Y. Coupled whole-tree optimality and xylem hydraulics explain dynamic biomass partitioning. New Phytol. 2021, 230, 2226–2245. [Google Scholar] [CrossRef]
- Mackay, D.S.; Savoy, P.R.; Grossiord, C.; Tai, X.; Pleban, J.R.; Wang, D.R.; McDowell, N.G.; Adams, H.D.; Sperry, J.S. Conifers depend on established roots during drought: Results from a coupled model of carbon allocation and hydraulics. New Phytol. 2020, 225, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Torres-Ruiz, J.M.; Albers, J.J. Conifer desiccation in the 2021 NW heatwave confirms the role of hydraulic damage. Tree Physiol. 2022, 42, 722–726. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhausser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Elshorbagy, A.; Barbour, S.L. Probabilistic approach for design and hydrologic performance assessment of reconstructed watersheds. J. Geotech. Geoenviron. Eng. 2007, 133, 1110–1118. [Google Scholar] [CrossRef]
- Alam, M.S.; Barbour, S.L.; Elshorbagy, A.; Huang, M.B. The impact of climate change on the water balance of oil sands reclamation covers and natural soil profiles. J. Hydrometeorol. 2018, 19, 1731–1752. [Google Scholar] [CrossRef]
- Hou, Q.C.; Huang, X.; Han, S.F. Study on the forming of “small olded-tree” and the transforming way in the Loess Plateau. J. Soil Water Conserv. 1991, 5, 64–72. [Google Scholar]
- Wang, L.; Li, Y.Y. Study on relationship between growth of artificial Robinia pseudoacacia plantation and soil desiccation in the Loess Plateau of northern Shannxi Province. Sci. Silvae Sin. 2004, 40, 84–91. [Google Scholar]
- Chen, H.S.; Shao, M.G.; Li, Y.Y. Soil desiccation in the Loess Plateau of China. Geoderma 2008, 143, 91–100. [Google Scholar] [CrossRef]
- Sperry, J.S.; Adler, F.R.; Campbell, G.S.; Comstock, J.P. Limitation of plant water use by rhizosphere and xylem conductance: Results from a model. Plant Cell Environ. 1998, 21, 347–359. [Google Scholar] [CrossRef]
- Wu, Y.Z.; Huang, M.B.; Warrington, D.N. Black locust transpiration responses to soil water availability as affected by meteorological factors and soil texture. Pedosphere 2015, 25, 57–71. [Google Scholar] [CrossRef]
- Feng, X.M.; Fu, B.J.; Piao, S.L.; Wang, S.; Ciais, P.; Zeng, Z.Z.; Lü, Y.H.; Zeng, Y.; Li, Y.; Jiang, X.H.; et al. Revegetation in China’s Loess Plateau is approaching sustainable water resource limits. Nat. Clim. Chang. 2016, 6, 1019–1022. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Zhao, T.N.; Shi, C.Q.; Ma, Q. Simulation of vegetation cover based on the theory of ecohydrological optimality in the Yongding River watershed, china. Forests 2021, 12, 1377. [Google Scholar] [CrossRef]
- Wang, S.F.; Yang, M.; Gao, X.D.; Li, B.; Cai, Y.H.; Li, C.J.; He, H.L.; Zhao, X.N. Ecohydrological response to deep soil desiccation in a semiarid apple orchard. Agric. For. Meteorol. 2024, 354, 110089. [Google Scholar] [CrossRef]
- Love, D.M.; Venturas, M.D.; Sperry, J.S.; Brooks, P.D.; Pettit, J.L.; Wang, Y.; Anderegg, W.R.L.; Tai, X.; Mackay, D.S. Dependence of aspen stands on a subsurface water subsidy: Implications for climate change impacts. Water Resour. Res. 2019, 55, 1833–1848. [Google Scholar] [CrossRef]
- Walthert, L.; Ganthaler, A.; Mayr, S.; Saurer, M.; Waldner, P.; Walser, M.; Zweifel, R.; von Arx, G. From the comfort zone to crown dieback: Sequence of physiological stress thresholds in mature European beech trees across progressive drought. Sci. Total Environ. 2021, 753, 141792. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, K.; Duan, C.H.; Li, G.L.; Zhen, Q.; Zheng, J.Y. Evaporation effect of infiltration hole and its comparison with mulching. Agric. Water Manag. 2023, 275, 108049. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.Y.; Li, G.L.; Ma, J.B.; Zhang, S.Q.; Zheng, J.Y. Effect of using an infiltration hole and mulching in fish-scale pits on soil water, nitrogen, and organic matter contents: Evidence from a 4-year field experiment. Land Degrad. Dev. 2021, 32, 4203–4211. [Google Scholar] [CrossRef]
- Song, X.L.; Wu, P.; Gao, X.D.; Yao, J.; Zou, Y.F.; Zhao, X.N.; Siddique, K.H.M.; Hu, W. Rainwater collection and infiltration (RWCI) systems promote deep soil water and organic carbon restoration in water-limited sloping orchards. Agric. Water Manag. 2020, 242, 106400. [Google Scholar] [CrossRef]
- Ma, C.K.; Yang, W.Y.; Zhou, B.B.; Wang, Q.J.; Shao, M.A. In situ rainwater harvesting system slows forest decline through increasing soil water content, fine-root traits, and plant hydraulic conductivity. Forests 2024, 15, 571. [Google Scholar] [CrossRef]
- Ma, L.H.; Wang, X.; Gao, Z.Y.; Wang, Y.K.; Nie, Z.Y.; Liu, X.L. Canopy pruning as a strategy for saving water in a dry land jujube plantation in a loess hilly region of China. Agric. Water Manag. 2019, 216, 436–443. [Google Scholar] [CrossRef]
- Wang, G.H.; Chen, Z.X.; Shen, Y.Y.; Yang, X.L. Thinning promoted the rejuvenation and highly efficient use of soil water for degraded Caragana korshinskii plantation in semiarid loessal regions. Land Degrad. Dev. 2023, 34, 992–1003. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, Y.J.; Hu, X.C.; Sun, S.J.; Zhang, J.S.; Cai, J.F.; Meng, P. Productivity and intrinsic water use efficiency of Robinia pseudoacacia plantations based on Biome-BGC model. For. Res. 2023, 23, 1–10. [Google Scholar]
- Li, P.; Li, H.J.; Si, B.C. Estimating deep soil water depletion and availability under planted forest on the Loess Plateau, China. Sci. Total Environ. 2024, 929, 172228. [Google Scholar] [CrossRef]







| Variable | Regression Equation | R2 | p |
|---|---|---|---|
| Minimum APLK | 0.885 − 0.494ELV − 0.543MAP + 0.151SA | 0.50 | 0.0001 |
| Maximum APLK | 1.314 − 0.648MAP − 0.177MAT − 0.399CC | 0.66 | 0.0001 |
| Mean APLK | 0.696 − 0.794MAP + 0.209SA | 0.77 | 0.0001 |
| DMRP | 0.687 − 0.687MAP − 0.300MAT + 0.243SA | 0.68 | 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-D.; Liu, T.-H.; Huang, M.-B.; Yan, X.-Y.; Liu, M.-H.; Yan, J.-H.; Chen, F.-Y.; Yan, W.; Niu, J.-Q. Spatial Pattern of Drought-Induced Mortality Risk and Influencing Factors for Robinia pseudoacacia L. Plantations on the Chinese Loess Plateau. Forests 2024, 15, 1477. https://doi.org/10.3390/f15081477
Zhang Z-D, Liu T-H, Huang M-B, Yan X-Y, Liu M-H, Yan J-H, Chen F-Y, Yan W, Niu J-Q. Spatial Pattern of Drought-Induced Mortality Risk and Influencing Factors for Robinia pseudoacacia L. Plantations on the Chinese Loess Plateau. Forests. 2024; 15(8):1477. https://doi.org/10.3390/f15081477
Chicago/Turabian StyleZhang, Zhong-Dian, Tong-Hui Liu, Ming-Bin Huang, Xiao-Ying Yan, Ming-Hua Liu, Jun-Hui Yan, Fei-Yan Chen, Wei Yan, and Ji-Qiang Niu. 2024. "Spatial Pattern of Drought-Induced Mortality Risk and Influencing Factors for Robinia pseudoacacia L. Plantations on the Chinese Loess Plateau" Forests 15, no. 8: 1477. https://doi.org/10.3390/f15081477