Abstract
Pinus massoniana Lamb. is an evergreen conifer; however, some current-year seedlings exhibit “winter reddening” at the onset of winter. The biological significance of this reddening is unclear. We examine the physiological responses of needles during the reddening process and explore the relationships between physiological traits and seeding cold resistance. Based on needle color, we recognize non-reddened, partially reddened, and fully reddened needle stages. As reddening progresses, chlorophyll fluorescence parameters (maximum light energy conversion efficiency of photosystem II (PSII) photochemistry, PSII potential activity, effective photosynthetic quantum yield, non-photochemical and photochemical quenching coefficients, and actual quantum yield of PSII photochemistry) decrease, reducing photosynthetic efficiency. Concurrently, the proportion of regulated energy dissipation in quantum yield of PSII decreases, and that of PSII non-regulated energy dissipation increases. Antioxidant enzyme activities (catalase and peroxidase) and osmoregulatory substances (soluble sugars and proteins and proline) increase, and malondialdehyde levels and relative cell damage at 4 °C and −10 °C gradually increase. Although P. massoniana seedlings adapt to low-temperature environments as their needles redden by increasing antioxidant enzyme activities and osmoregulatory substances and by adjusting photosynthetic efficiency and light energy distribution, cell membrane damage persists. Cold resistance in P. massoniana seedlings is not fully established during winter reddening.
1. Introduction
Abiotic stresses, such as low temperature, high light intensity, ultraviolet radiation, and drought, can trigger complex oxidative stress responses in plants that involve physiological, molecular biological, metabolic, and leaf color changes [1,2]. Leaf color, an essential phenotypic trait of plants, is influenced by multiple regulatory factors [3]. Under low-temperature stress, thylakoid membranes undergo lipid peroxidation, leading to the accumulation of malondialdehyde (MDA), which damages the cell-membrane system. This results in physiological metabolic disorders, abnormal growth and development, and changes in leaf color and simultaneously activates the plant′s antioxidant defense mechanism [4]. This defense mechanism, comprising enzymatic and non-enzymatic components, balances the synthesis and scavenging of reactive oxygen species (ROS) to prevent cell damage [5,6] and plays an important role in enhancing the plant′s cold tolerance [7,8,9]. Enzymatic antioxidants, which are part of the antioxidant enzyme system, and non-enzymatic antioxidants, including osmolytes (soluble sugars (Ss), soluble proteins (Sp), and proline (Pro)), carotenoids, and anthocyanins, comprise this defense system [10].
The photosynthetic system of plants subjected to low-temperature stress is affected by changes in stomatal aperture, biosynthesis of photosynthetic pigments, and photosystem activity, leading to alterations in overall photosynthetic capacity and efficiency [11]. Chlorophyll fluorescence analysis has been widely used to evaluate resistance traits and mechanisms in response to environmental stress, to identify the sites and extent of damage to the photosynthetic apparatus, and to indicate the degree of injury and level of plant cold tolerance [12,13]. During the leaf coloring process of Liquidambar formosana, the maximum photochemical efficiency, actual photochemical efficiency, and electron transfer rate decline rapidly, indicating that changes in leaf color can affect photosynthetic efficiency [14]. However, most research on photosynthetic characteristics related to leaf color change has examined broad-leaved trees as opposed to evergreen conifers.
Leaf reddening is a common phenomenon that occurs in various plant developmental stages throughout the year [15]. More research has examined leaf reddening during spring and summer than during autumn and winter (“winter reddening”) [16,17,18]. In many evergreen trees, leaf reddening is a temporary winter phenomenon, and leaves regain their green color in spring [19]. The applicability of winter reddening remains controversial. Reddening may represent an antioxidant stress response, enabling plants to cope with environmental factors. During winter, cold, dry, and bright sunlight conditions can lead to excessive energy capture, photoinhibition, ROS formation, and greater photooxidative damage. Under these conditions, plant leaves often change color, turning brown, red, or purple, suggesting that winter leaf reddening is a mechanism to mitigate these stressors [16,20]. In the Japanese cedar Cryptomeria fortunei, the yellow-brown coloration of needles under low-temperature stress is caused by the accumulation of flavonoids, which may enhance cold tolerance [21]. However, for the mountain tallow Triadica cochinchinensis, MDA content increases as leaves redden, suggesting that it does not improve the species′ ability to withstand winter cold environments [22].
Pinus massoniana Lamb., an evergreen conifer native to China, is an important economic and forestry tree. During cultivation, at the onset of winter, some current-year seedlings exhibit winter reddening of needles. The physiological changes and their interrelationships during needle reddening in winter are not well understood, limiting the understanding of the biological significance of this phenomenon. Therefore, using current-year P. massoniana seedlings, we investigate dynamic physiological changes during needle reddening stages under low-temperature winter conditions. Changes in osmotic regulatory substances, antioxidant enzyme activities, chlorophyll fluorescence parameters, and the cell membrane system during development are analyzed. In addition, physiological responses during needle reddening in winter are elucidated, and the relationship between these physiological traits and cold resistance is explored. These results can be applied to provide theoretical support for further research on the biological significance of winter reddening in pine seedlings
2. Materials and Methods
2.1. Study Site and Plant Materials
P. massoniana seeds were sourced from the Duyun Ma′anshan state-owned forest farm (26°90′ N, 107°180′ E, 920 m elevation) in China. At the end of April 2022, container seedlings were cultivated in a greenhouse nursery at Guizhou University (26°4′ N, 106°7′ E). Container measurements were 8.8 cm (outer diameter), 5.8 cm (bottom diameter), and 7.6 cm (height). The substrate mixture comprised peat:coir:pumice:perlite at a 5:2:2:1 ratio, following the cultivation and management methods of Wang et al [23]. On 23 December 2022, 100 eight-month-old seedlings were randomly selected and exposed to outdoor cold treatment. Needle samples were collected after the chlorophyll fluorescence parameter measurements were completed at three developmental stages based on color change: NR (non-reddened), when all plants exhibited normal green needles (23 December 2022); PR (partially reddened), with approximately 50% of needles showing red coloration (28 December 2022); and FR (fully reddened), when all needles appeared red (7 January 2023) (Figure 1). Four seedlings, each one used as a biological replicate, were randomly harvested at each stage and rinsed with distilled water. Then, their phenotype was documented using photographs. Needle samples were then promptly frozen in liquid nitrogen and stored at −80 °C for further analysis.

Figure 1.
The winter reddening process in P. massoniana seedling needles. NR: non-reddening; PR: partial reddening; FR: full reddening.
2.2. Chlorophyll Fluorescence Parameters
Following Wang et al. [24], chlorophyll fluorescence parameters of P. massoniana seedling needles were measured using MONITORING-PAM (Heinz Walz GmbH, Effeltrich, Germany). Prior to measurements, a dark adaptation period of 35–40 min was allowed. Slow induction kinetics curves were determined using light (<0.5 μmol·m−2s−1) to measure initial fluorescence (Fo), followed by a 0.6 s saturating pulse (approximately 10,000 μmol·m−2s−1) to measure maximum fluorescence (Fm). From Fo and Fm, variable fluorescence (Fv), maximum light energy conversion efficiency of PSII photochemistry (Fv/Fm), PSII potential activity (Fv/Fo), effective photosynthetic quantum yield (Fv′/Fm′), actual quantum yield of PSII photochemistry (Y(II)), photochemical quenching coefficient (qP), non-photochemical quenching coefficient (NPQ), quantum yield of PSII regulated energy dissipation (Y(NPQ)), and quantum yield of PSII non-regulated energy dissipation (Y(NO)) were computed (Table 1). Four biological replicates were examined for each treatment.

Table 1.
Chlorophyll fluorescence yield equations.
2.3. Physiological Characteristics
Peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), Ss, Pro, Sp, and MDA were measured using reagent test kits provided by Suzhou Michy Biomedical Technology Co., Ltd. (Suzhou, China). The experiment included four biological replicates for each treatment.
2.4. Cell Membrane Damage
Following methods of Climent et al. [25], the relative ion leakage from plant tissues was determined to assess membrane damage. Ten needles from seedlings at NR, PR, and FR stages were collected. The apical and basal ends of each needle were removed, and the needles were cut into approximately 15 mm segments. Each sample was replicated four times and placed in 15 mL centrifuge tubes. Control samples were kept at 4 °C. For samples frozen at −10 °C, approximately 0.5 g of crushed ice (made using ultra-pure water) was added to tubes, and samples were left for 4 h. After thawing at 4 °C h−1, 10 mL of ultra-pure water was added to each centrifuge tube containing needle samples, and electrolyte conductivity was measured at 25 °C (C1) using a conductivity meter 24 h later. Samples were then boiled in a high-temperature (120 °C) and high-pressure sterilizer at for 30 min and kept at 25 °C for 12 h. Then, total electrolyte conductivity (C2) was measured. Each treatment was determined with four biological replicates. The formula was as follows:
The relative cellular damage of the unfrozen (4 °C) control samples was evaluated as follows:
R0 = (C1/C2) × 100%,
The relative cellular damage of the frozen samples at −10 °C was assessed as follows:
Rt = (C1/C2) × 100%,
2.5. Statistical Analysis
Data were organized and calculated using Excel, and statistical analyses were performed using SPSS software (version 23.0). One-way analysis of variance (ANOVA) was applied to evaluate the significance of findings, with thresholds established at p < 0.05 (significant) and p < 0.01 (highly significant). Various physiological and biochemical markers throughout the winter reddening process were analyzed with correlation and principal component analysis (PCA) using Origin 2021 software, from which relevant figures were also generated. Results for each parameter are presented as means ± standard errors (SE). Methodologies for conducting stepwise regression and pathway analyses follow protocols outlined by Wang et al. [23], and the determination coefficient was calculated following Xie et al. [26].
3. Results
3.1. Needle Chlorophyll Fluorescence
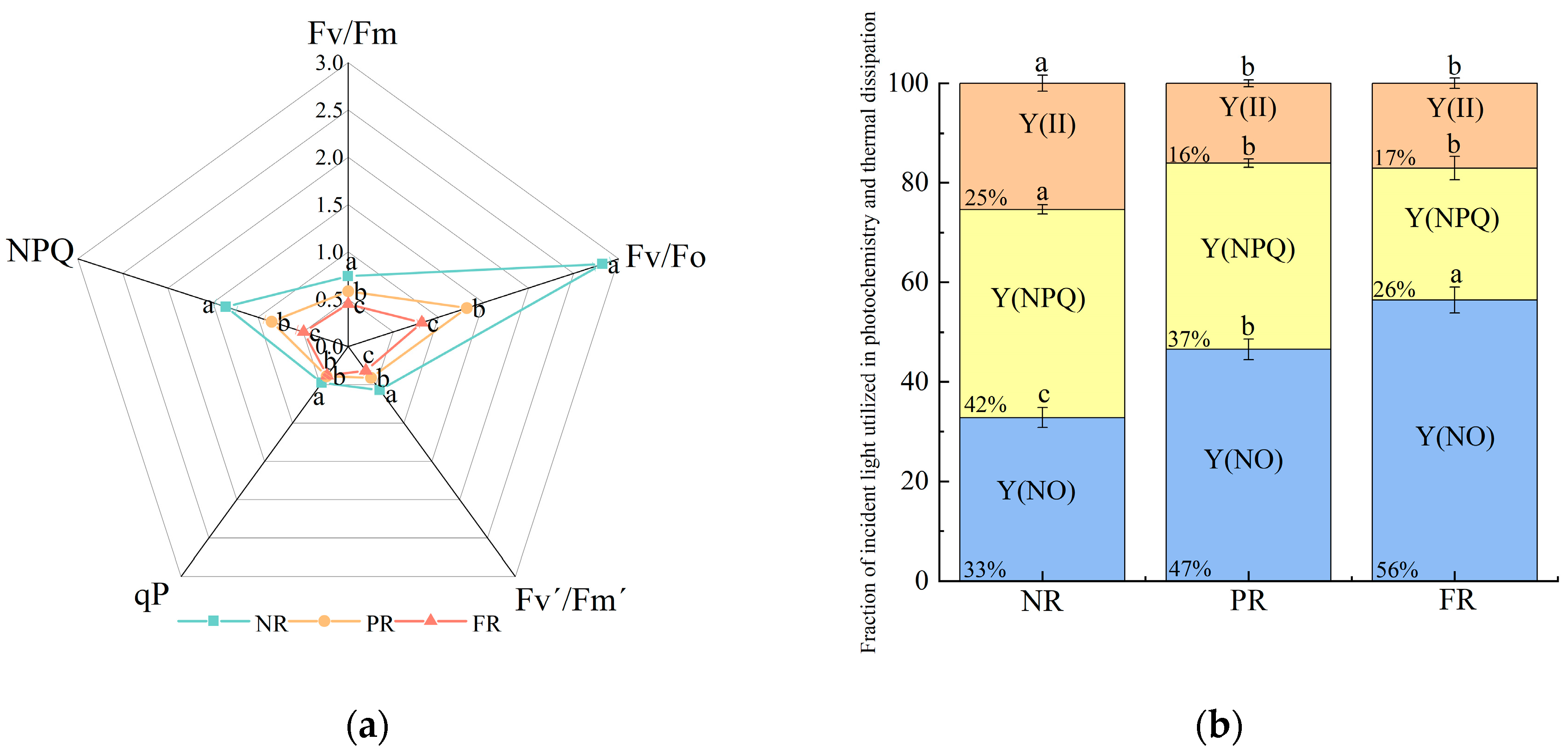
Fv/Fm and Fv/Fo decreased significantly (Figure 2a), indicating a reduced potential of seedlings to convert light into chemical energy as their needles red. This suggests that low winter temperatures increasingly impact seeding photosynthetic efficiency. NPQ and qP values during the PR and FR stages were significantly lower than those during the NR stage. No significant differences in qP values were noted between PR and FR stages. Additionally, Fv′/Fm′ also significantly decreased as needle reddening progressed (Figure 2a). These results indicate the gradual reductions in the efficiency of light energy conversion and heat dissipation capacity during the needle reddening process. Simultaneously, the proportion of Y(II) decreased and stabilized during the PR and FR stages. In addition, the proportion of Y(NPQ) gradually decreased, whereas that of Y(NO) gradually increased (Figure 2b). This suggests that the light energy distribution patterns of needles changed as they reddened, and the energy was reallocated to Y(NO).
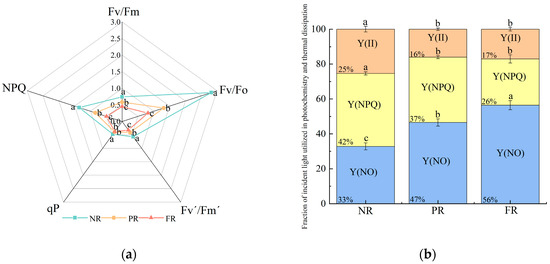
Figure 2.
P. massoniana seedling (a) chlorophyll fluorescence induction parameters and (b) fraction of incident light used in photochemistry and thermal dissipation during winter reddening of needles. Fv/Fm: maximum light energy conversion efficiency of PSII photochemistry; Fv/Fo: PSII potential activity; Y(II): actual quantum yield of PSII photochemistry; Fv′/Fm′: effective photosynthetic quantum yield; qP: photochemical quenching coefficient; NPQ: non-photochemical quenching coefficient; Y(NPQ): quantum yield of PSII-regulated energy dissipation; Y(NO): quantum yield of PSII non-regulated energy dissipation. Vertical bars represent means ± standard error (SE). Different letters signify statistical differences at p < 0.05, as determined by Duncan′s multiple range test (n = 4). Abbreviations: NR, non-reddening; PR, partial reddening; FR, full reddening.
3.2. Osmoregulatory Substances in the Needles
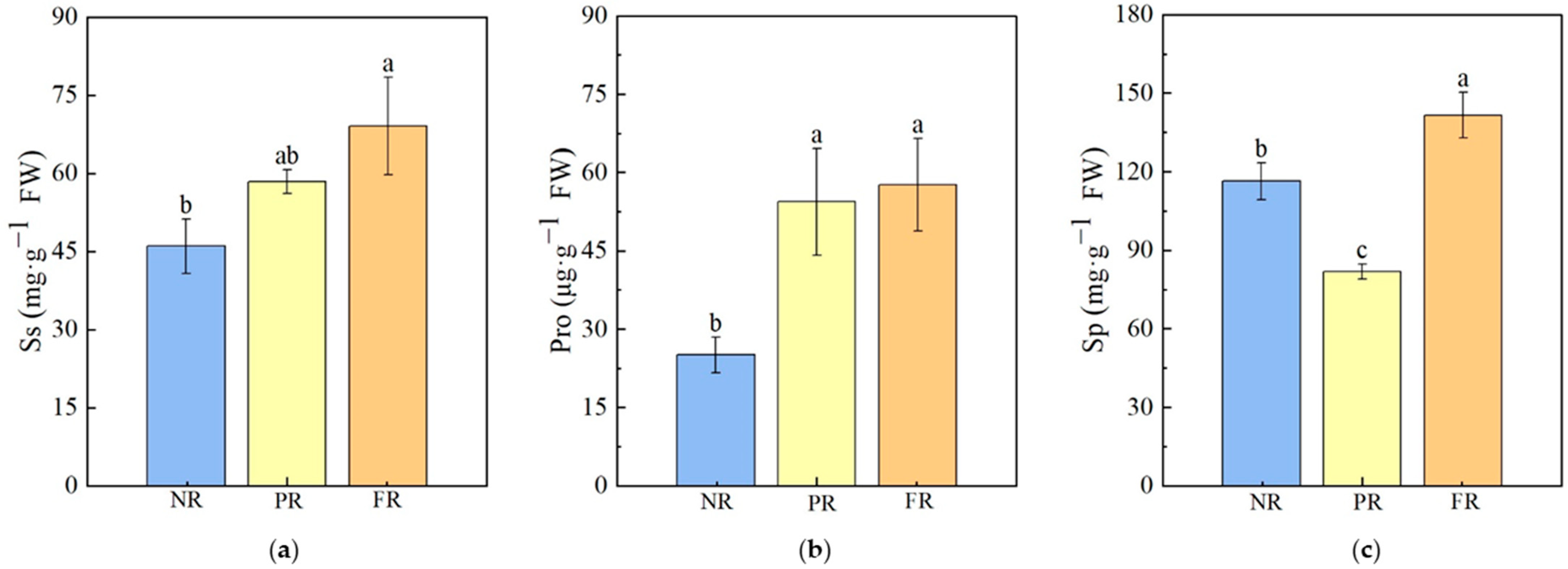
During the green-to-red needle color transition, changes in the contents of osmotic regulatory substances occurred. The Ss content gradually increased with needle reddening, with significantly higher levels observed in the FR stage compared with the NR stage. However, no significant differences were found between PR and either the NR or FR stages. Pro levels during the PR and FR stages were significantly higher than those during the NR stage; no significant differences in Pro levels were noted between the PR and FR stages. Sp contents during the FR stage were significantly higher than those during either NR or PR stages. The following trend was observed: FR > NR > PR (Figure 3).
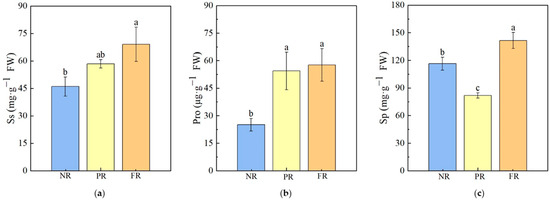
Figure 3.
P. massoniana seedling osmoregulatory substances in needles. (a) Ss: soluble sugar. (b) Pro: Proline. (c) Sp: soluble protein. Vertical bars represent means ± standard error (SE). Different letters signify statistical differences at p < 0.05, as determined by Duncan′s multiple range test (n = 4). Abbreviations: NR, non-reddening; PR, partial reddening; FR, full reddening.
3.3. Needle Antioxidant Enzyme Activities
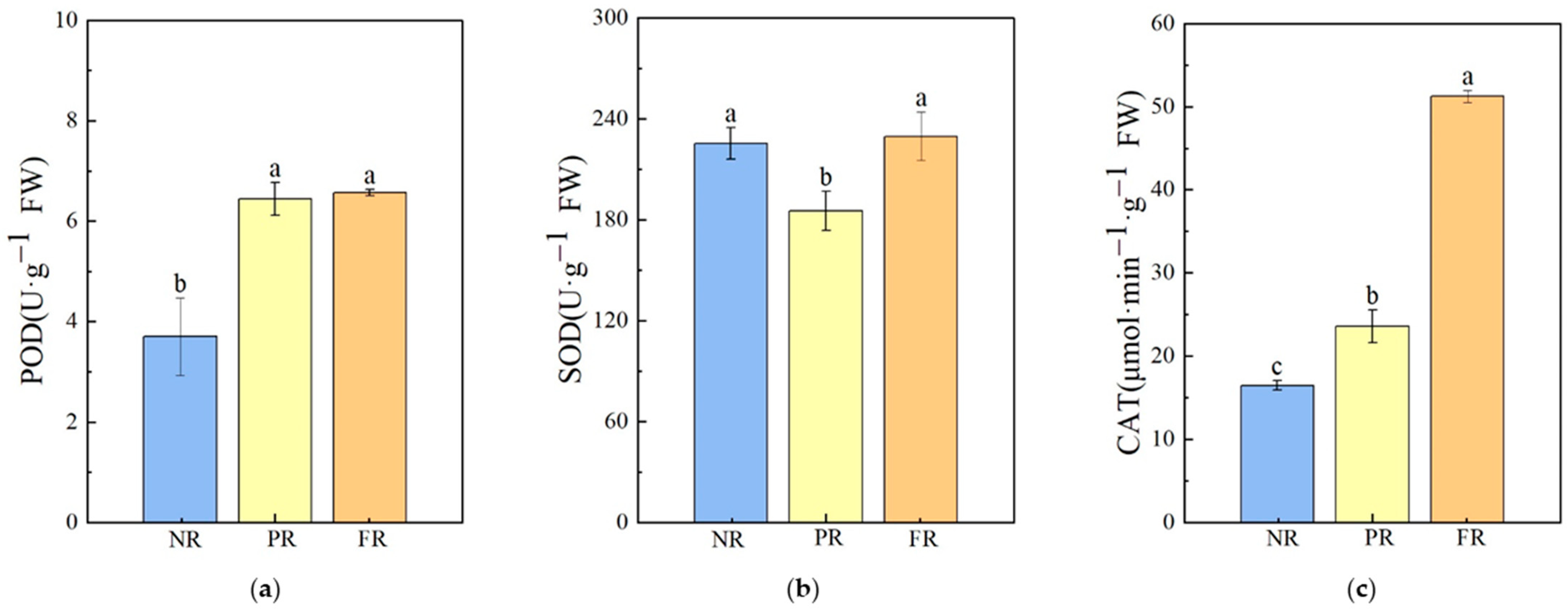
Compared with the NR stage, POD enzyme activities during the PR and FR stages significantly increased; there was no significant difference in POD enzyme activities between the PR and FR stages. SOD enzyme activity significantly decreased during the PR stage and recovered to a level similar to that of the NR stage during the FR stages. CAT enzyme activity increased significantly. Compared with the NR stage, there was a 43.07% increase in CAT activity during the PR stage, and a 117.49% increase was noted during the FR stage when compared with the PR stage (Figure 4). These results indicate that during the winter needle reddening process, the antioxidant response primarily involved an increase in CAT and POD enzyme activities.
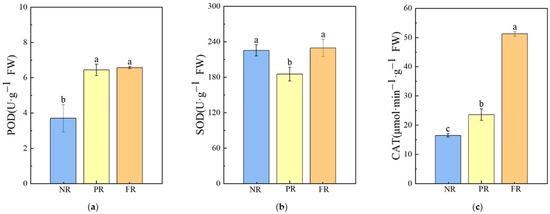
Figure 4.
Antioxidant enzyme activities in P. massoniana seedling needles during winter reddening. (a) POD: peroxidase. (b) SOD: superoxide dismutase. (c) CAT: catalase. Vertical bars represent means ± standard error (SE). Different letters signify statistical differences at p < 0.05, as determined using Duncan′s multiple range test (n = 4). Abbreviations: NR, non-reddening; PR, partial reddening; FR, full reddening.
3.4. Cell Membrane Damage in Needles
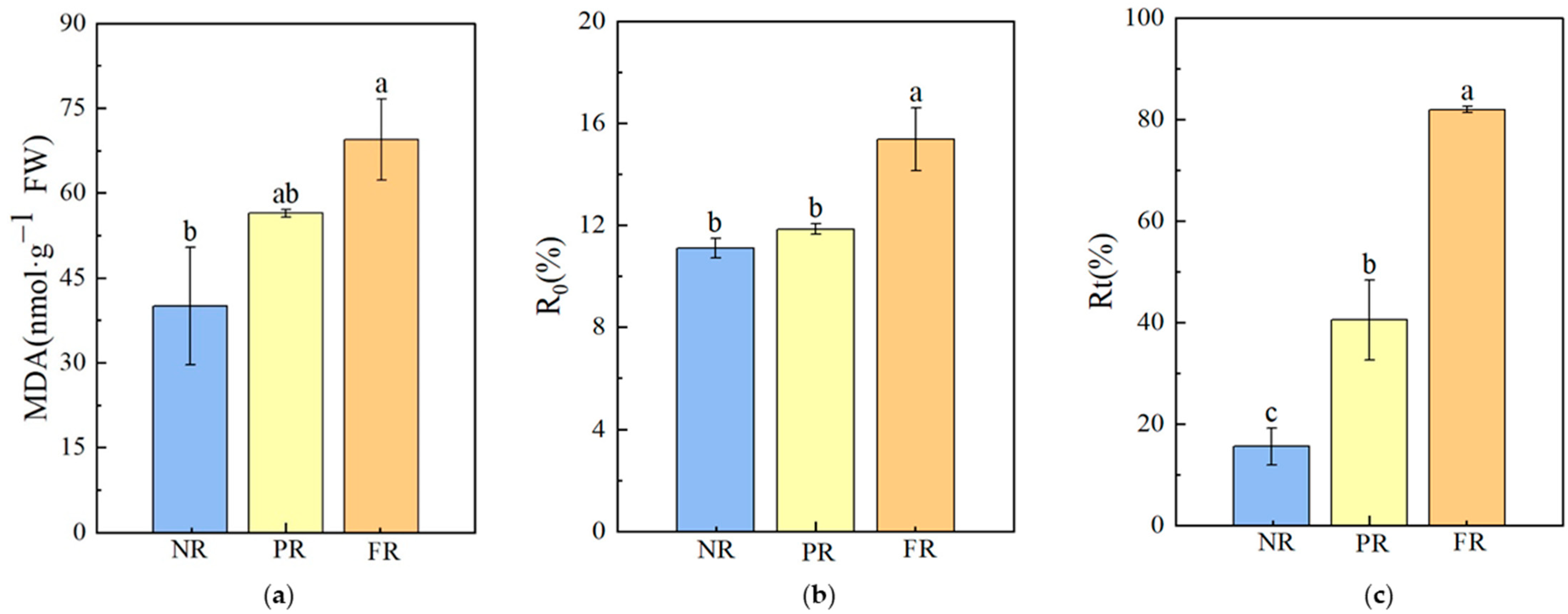
During the FR stage, MDA and the R0 value at 4 °C were significantly higher compared with NR stage values, with increases of 73.74% and 38.47%, respectively. No significant differences in MDA and R0 values were noted between NR and PR stages. As needles reddened, the Rt value at −10 °C significantly trended upward (Figure 5), increasing by 5.28× that of the NR to FR stages. This indicates a higher degree of cell membrane damage during the FR stage than during the NR stage. Additionally, Rt values at −10 °C for the NR, PR, and FR stages all exceeded the R0 value obtained at 4 °C during the FR stage, suggesting that the degree of cell membrane damage in freezing conditions (−10 °C) was more severe than under 4 °C during the needle reddening process.
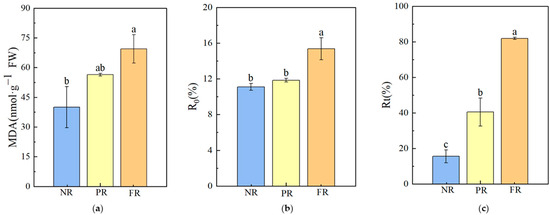
Figure 5.
P. massoniana seedling cell membrane damage characteristics during winter reddening. (a) MDA: malondialdehyde. (b) R0: relative cell damage at 4 °C. (c) Rt: relative cell damage at −10 °C. Vertical bars represent means ± standard error (SE). Different letters signify statistical differences at p < 0.05, as determined using Duncan′s multiple range test (n = 4). Abbreviations: NR, non-reddening; PR, partial reddening; FR, full reddening.
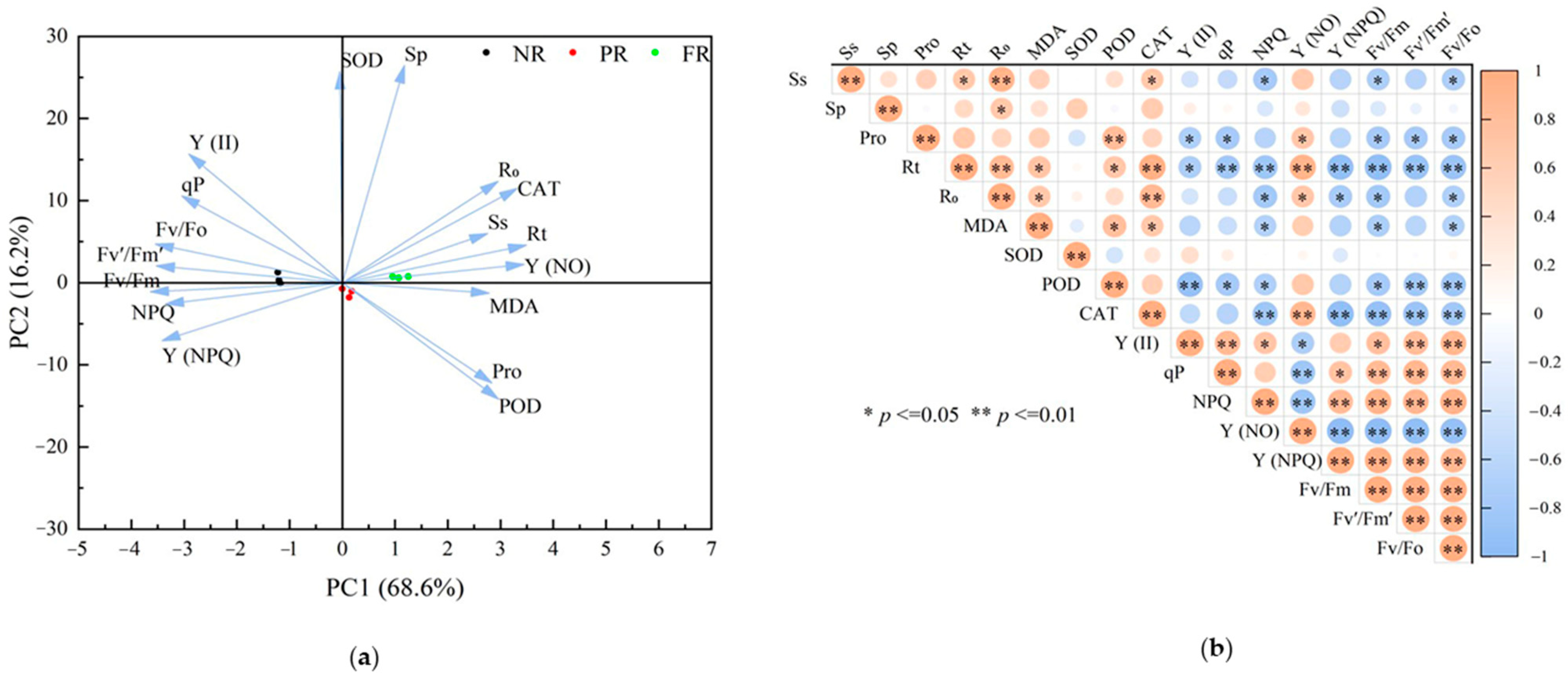
3.5. Relationships between Physiological Traits
To examine relationships between cell membrane damage and physiological traits (chlorophyll fluorescence parameters, osmotic regulators, and antioxidant enzyme activities) during the NR, PR, and FR stages, we used PCA. The first two PCA axes effectively differentiated samples (PC1, 68.6%; PC2,16.2%) (Figure 6a). Correlation analysis revealed degrees of correlations between R0 and various physiological traits during the winter reddening process. Specifically, R0 correlated significantly or highly significantly and positively with Rt, MDA, CAT, Y(NO), Sp, and Ss. Conversely, R0 correlated significantly negatively with Fv/Fm, Fv/Fo, Y(NPQ), and NPQ (Figure 6b).
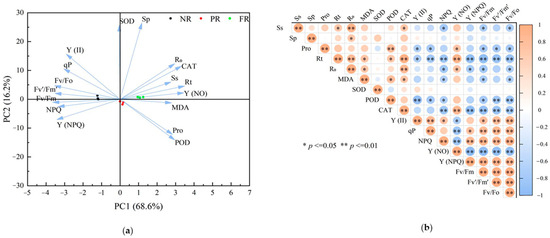
Figure 6.
Principal component and correlation of physiological characteristics in winter reddening of P. massoniana seedlings needles. (a) Principal component analysis (PCA) with the color of dots representing different stages of reddening, arrows indicating the relationships between each physiological index and the principal components, and the angle between the arrow and axis reflecting the degree of correlation. (b) Correlation analysis showcasing significant relationships.
3.6. Effect of Relative Factors on the Degree of Cell Membrane Damage
To identify relationships between multiple independent and dependent variables, our linear regression analysis yielded the following equation:
y = 7.125 + 0.094 x1 + 0.008 x2 + 0.082 x3 − 6.582 x4 − 0.009 x5 + 0.02 x6,
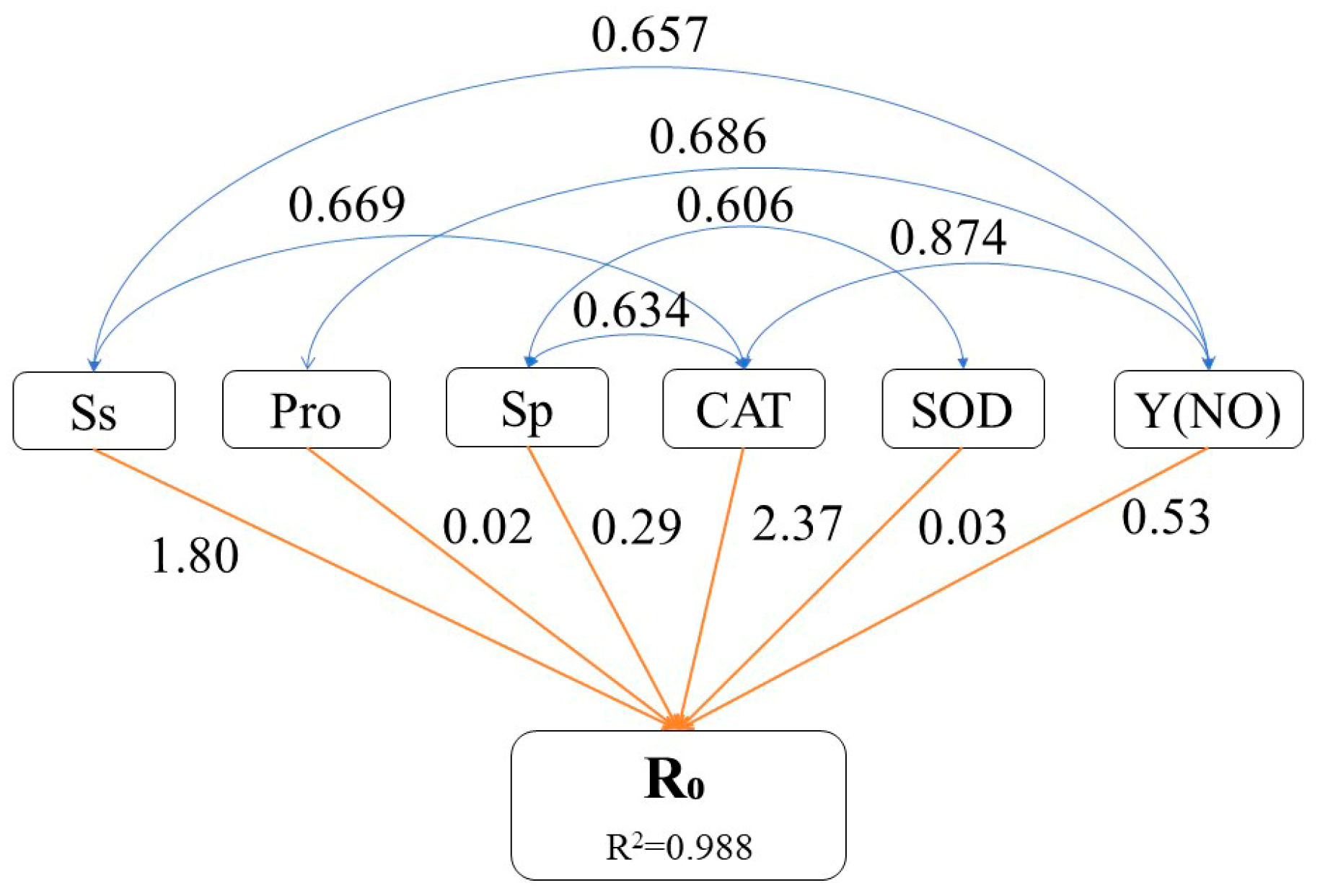
Path analysis was performed to identify physiological traits that exerted a significant influence on the extent of cell membrane damage. The decision coefficient for the increase in R0 was the highest for elevated CAT enzyme activity (2.37), indicating that increased CAT enzyme activity most significantly impacted the R0 value (Figure 7). Additionally, elevated levels of Ss and Y(NO) had relatively substantial decision coefficients for the increase in R0 value (1.80 and 0.53, respectively), signifying their importance in augmenting the R0 value. Changes in Sp and Pro contents as well as SOD enzyme activity had relatively minor decision coefficients for increasing the R0 value (0.29, 0.02, and 0.03, respectively), suggesting that these factors had little impact on the increased R0 value.
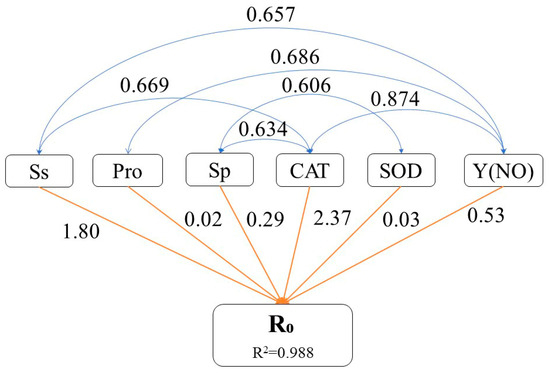
Figure 7.
Pathway analysis of physiological characteristics of P. massoniana seedling needles during winter reddening. Pathway analysis using the R0 value as the dependent variable, illustrating how various factors contribute to the degree of cell membrane damage. Pathways in blue indicate that the correlation coefficient is above 0.6, whereas those in orange indicate the determination coefficient. For clarity, only the most relevant pathways are shown, omitting non-significant interactions. Ss: soluble sugar; Pro: proline; Sp: soluble protein; R0: relative cell damage at 4 °C; SOD: superoxide dismutase; CAT: catalase; Y(NO): quantum yield of PSII non-regulated energy dissipation.
4. Discussion
4.1. Chlorophyll Fluorescence Parameters in Winter-Reddened Needles
Changes in the color of leaves, the primary organs for photosynthesis in plants, can reduce absorption of light energy, thereby affecting photosynthesis [14]. When this occurs, one of the first parameters to change is the Fv/Fm value. This change is an indication of the extent of environmental stress on plant photosynthesis which [27], under optimal conditions, typically ranges from 0.75 to 0.85 [28]. Fv/Fm values for the spruce Picea [29] and purple coneflower Echinacea purpurea [30] decreased under low-temperature stress. We report that the needles of P. massoniana seedlings continuously turn red during winter, and the Fv/Fm values during the PR and FR stages are significantly lower than those observed during the NR stage (0.75). This trend is consistent with the decrease in the PSII potential activity Fv/Fo (Figure 2), indicating that seedlings were significantly affected by low temperatures, leading to a gradual decline in Fv/Fo and Fv′/Fm′ [31,32]. Additionally, both qP and Y(II) trended downwards, indicating a decline in overall photosynthetic efficiency with continued reddening.
When environmental factors affect plant photosynthesis, plants often protect themselves by reducing the photosynthetic activity of the leaves and chloroplasts and dissipating absorbed energy [33]. NPQ in red leaves is usually higher than in green leaves because red leaves often dissipate excess light energy as heat to protect the photosystem [14]. However, we report that the NPQ of P. massoniana needles decreases as the needles turn red, indicating that excess light energy was not being dissipated as heat during needle reddening. Furthermore, green needles of P. massoniana seedlings can reduce photosystem damage caused by excess light energy by increasing Y(NPQ). However, the light energy absorbed by the PSII reaction center is distributed to Y(II), Y(NPQ), and Y(NO); thus, Y(II) + Y(NPQ) + Y(NO) = 1 [34]. During winter reddening of P. massoniana seedling needles, the proportions of Y(II) and Y(NPQ) decreased, and that of Y(NO) gradually increased. These results indicate that during winter reddening of P. massoniana seedling needles, the overall needle photosynthetic efficiency decreases, and that energy is reallocated to Y(NO) to regulate the appropriate photosynthetic efficiency for seedling survival.
4.2. Changes in Antioxidant Enzymes and Osmotic Regulators during Needle Reddening
When plants are damaged by cold, a series of defense mechanisms (activation of antioxidant enzyme systems and osmotic regulating substances) are triggered [4,35,36]. The antioxidant enzyme system comprises POD, SOD, and CAT, which eliminate oxidative damage caused by excessive ROS. SOD converts excess oxygen radicals in plants into H2O2, and CAT and POD then convert H2O2 into H2O and O2, alleviating oxidative damage to membrane systems and enhancing plant cold resistance [37]. Different tree species use different enzymes to enhance antioxidant capabilities during winter leaf reddening. For example, the savin juniper Sabina vulgaris uses POD and CAT to exert antioxidant functions during leaf color changes in winter [38], whereas winter red leaves of the Chinese buxus Buxus microphylla primarily use SOD and CAT [39]. For P. massoniana, we report a continuous increase in POD and CAT, but no significant difference in SOD activity between the FR and NR stages. Therefore, during the red needle stage, CAT and POD primarily work together to promote antioxidation rather than SOD. Because CAT activity increased significantly and POD activity did not differ between the PR and FR stages, CAT is the main antioxidant enzyme used during the winter red needle process in P. massoniana.
Osmotic substances (Ss, Sp, and Pro) are important components of non-enzymatic antioxidants, and their increased contents play important roles in enhancing plant tolerance to low temperatures [35,36]. Ss protects frost-resistant tissues by stabilizing membrane structures and proteins and represents an essential substance to improve overwinter cold resistance [40,41]. The accumulation of Ss stabilizes cytoplasmic concentrations, helps to preserve water, maintains cell membrane stability, and prevents ice crystal formation between cells at low temperatures, regulating plant adaptability under cold stress [42,43]. For P. massoniana, we report that Ss contents continuously trend upwards with needle reddening, indicating that their increased contents can regulate seeding adaptability during winter. This finding is similar to results reported by Wang et al. [44]. Sp is strongly hydrophilic and can enhance cell water-retention capacity, regulate cell turgor pressure, and lower the cytoplasmic freezing point. The accumulation of Sp in low-temperature stressed environments helps improve plant cold resistance [45]. We report that Sp contents peak in the FR stage, suggesting that Sp is involved in enhancing seedling cold resistance during winter needle reddening. Pro occurs widely in plants and serves as an important physiological indicator for evaluating cold resistance; its functions include scavenging ROS, balancing intracellular homeostasis during stress processes, regulating plant growth and development, and acting as a signaling molecule that influences other metabolic pathways [8,46]. We report that Pro contents increase with needle reddening, but there is no significant difference in Pro contents between the PR and FR stages. This phenomenon may be because increased Pro contents within cells when seedlings lose water at low temperatures leads to higher liquid concentrations within cells, thereby protecting the cell against cell freezing and maintaining a stable cell membrane [47].
4.3. Cold Resistance of Seedlings during Winter Reddening
MDA levels increase in plants at low winter temperatures. This leads to extracellular leakage and increased electrolyte permeability, which damage the cell membrane system [7]. MDA is a byproduct of lipid peroxidation and is released when cell membranes are damaged. Additionally, MDA [48] and relative cell membrane damage levels [25] are often used to indicate plant cold resistance. Rt and R0 values indicate the relative degree of cell membrane damage. Therefore, high MDA, Rt, and R0 values indicate severe cell membrane damage and weak cold resistance, while low MDA, Rt, and R0 values indicate little or no cell membrane damage and strong cold resistance [25,46]. For P. massoniana, we examined the relationships between physiological traits and MDA, Rt, and R0 values. We report a significant positive correlation between R0 values and CAT, Y(NO), and Ss content (Figure 6). Pathway analysis reveals that the decision coefficients of CAT enzyme activity, Ss content, and Y(NO) proportion on R0 values were relatively high (2.37, 1.80, and 0.53, respectively). Furthermore, with changes in osmotic regulators, antioxidant enzyme activity, and chlorophyll fluorescence parameters, no significant changes in R0 values were noted during the NR and PR stages. This indicates that increased Ss, Sp, and Pro contents; CAT and POD enzyme activity levels; and allocation of energy to Y(NO) facilitated the adaptation of seedlings to cold environments. Among these factors, CAT enzyme activity, Ss content, and Y(NO) proportion played important roles in the cold resistance of seedlings. Although these physiological traits can regulate the ability of seedlings to cope with low-temperature environments during the reddening process, the accumulation of MDA continues to increase and both R0 and Rt values trend upward as needles redden. This phenomenon suggests that during the winter reddening of seedling needles, the ability of seedlings to withstand low temperatures is not enhanced.
5. Conclusions
During the winter reddening process of needles of P. massoniana seedlings, the primary mechanisms involved include the downregulation of overall photosynthetic efficiency and alterations in seedling energy allocation patterns. Energy was directed toward Y(NO) to regulate photosynthetic efficiency for seeding survival. CAT and POD antioxidant enzyme activities and the osmotic regulatory substances Ss, Sp, and Pro, increased, enabling seedlings to adapt to low-temperature environments. However, seedlings exhibited no significant improvement in cold or frost resistance as a consequence.
Author Contributions
H.H.: methodology, investigation, formal analysis, writing—original draft, and writing—review & editing. Y.X.: investigation and formal analysis. S.X.: investigation. X.L.: investigation. H.W.: writing—review & editing. Y.Z.: writing—review & editing. F.W.: conceptualization, methodology, writing—review & editing, project administration, funding acquisition, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (grant numbers 32260406 and 31660201).
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Natural variation in flavonol and anthocyanin metabolism during cold acclimation in Arabidopsis thaliana accessions. Plant Cell Environ. 2015, 38, 1658–1672. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Shinohara, K.; Kakubari, Y.; Mukai, Y. Photoprotective role of rhodoxanthin during cold acclimation in Cryptomeria japonica. Plant Cell Environ. 2003, 26, 715–723. [Google Scholar] [CrossRef]
- Zhao, M.H.; Li, X.; Zhang, X.X.; Zhang, H.; Zhao, X.Y. Mutation mechanism of leaf color in plants: A review. Forests 2020, 11, 851. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, A.; Zhang, J.; Jiang, M. Abscisic acid is a key inducer of hydrogen peroxide production in leaves of maize plants exposed to water stress. Plant Cell Physiol. 2006, 47, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Jezek, M.; Allan, A.C.; Jones, J.J.; Geilfus, C.M. Why do plants blush when they are hungry? New Phytologist 2023, 239, 494–505. [Google Scholar] [CrossRef]
- Wang, S.M.; Wang, Y.S.; Su, B.Y.; Zhou, Y.Y.; Chang, L.F.; Ma, X.Y.; Li, X.M. Ecophysiological responses of five mangrove species (Bruguiera gymnorrhiza, Rhizophora stylosa, Aegiceras corniculatum, Avicennia marina, and Kandelia obovata) to chilling stress. Front. Mar. Sci. 2022, 9, 846566. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Liu, L.J.; Chen, W.F.; Wang, K.; Zhang, C.X. Effects of different irrigation minima on green period and cold-resistance physiological indi-ces of Festuca arundinacea. Chin. J. Appl. Ecol. 2012, 23, 38–44. [Google Scholar]
- Aazami, M.A.; Asghari-Aruq, M.; Hassanpouraghdam, M.B.; Ercisli, S.; Baron, M.; Sochor, J. Low temperature stress mediates the antioxidants pool and chlorophyll fluorescence in Vitis vinifera L. cultivars. Plants 2021, 10, 1877. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Wang, Y.; Xiao, Y.; Yang, J.; Wang, R.; Jiang, Y.; Huang, R.; Liu, X.; Jiang, Y. Relationships between leaf color changes, pigment levels, enzyme activity, photosynthetic fluorescence characteristics and chloroplast ultrastructure of Liquidambar formosana Hance. J. For. Res. 2022, 33, 1559–1572. [Google Scholar] [CrossRef]
- Archetti, M.; Döring, T.F.; Hagen, S.B.; Hughes, N.M.; Leather, S.R.; Lee, D.W.; Lev-Yadun, S.; Manetas, Y.; Ougham, H.J.; Schaberg, P.G.; et al. Unravelling the evolution of autumn colours: An interdisciplinary approach. Trends Ecol. Evol. 2009, 24, 166–173. [Google Scholar] [CrossRef]
- Hughes, N.M.; Morley, C.B.; Smith, W.K. Coordination of anthocyanin decline and photosynthetic maturation in juvenile leaves of three deciduous tree species. New Phytologist 2007, 175, 675–685. [Google Scholar] [CrossRef]
- Karageorgou, P.; Manetas, Y. The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 2006, 26, 613–621. [Google Scholar] [CrossRef]
- Chen, M.; Li, H.; Zhang, W.; Huang, L.; Zhu, J. Transcriptomic analysis of the differences in leaf color formation during stage transitions in Populus× euramericana ‘Zhonghuahongye’. Agronomy 2022, 12, 2396. [Google Scholar] [CrossRef]
- Nikiforou, C.; Zeliou, K.; Kytridis, V.P.; Kyzeridou, A.; Manetas, Y. Are red leaf phenotypes more or less fit? The case of winter leaf reddening in Cistus creticus. Environ. Exp. Bot. 2010, 67, 509–514. [Google Scholar] [CrossRef]
- Hughes, N.M.; Neufeld, H.S.; Burkey, K.O. Functional role of anthocyanins in high-light winter leaves of the evergreen herb Galax urceolata. New Phytologist 2010, 168, 575–587. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, L.; Hu, H.; Yang, J.; Cui, J.; Wei, G.; Xu, J. Transcriptome and metabolome changes in Chinese cedar during cold acclimation reveal the roles of flavonoids in needle discoloration and cold resistance. Tree Physiol. 2022, 42, 1858–1875. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.P. Screening of Superior Triadica cochinchinensis Loureiro Plants and Studyon the Mechanism of Leaf Color Changeunder Low Temperature Stress. Master’s Thesis, College of Horticulture Fujian Agriculture and Forestry University, Fuzhou, China, 2022. [Google Scholar]
- Wang, H.; Wu, F.; Li, M.; Liang, D.; Ding, G. Heteroblastic foliage affects the accumulation of non-structural carbohydrates and biomass in Pinus massoniana (Lamb.) Seedlings. Forests 2021, 12, 1686. [Google Scholar] [CrossRef]
- Wang, H.; Wu, F.; Li, M.; Zhu, X.; Shi, C.; Ding, G. Morphological and physiological responses of Pinus massoniana seedlings to different light gradients. Forests 2021, 12, 523. [Google Scholar] [CrossRef]
- Climent, J.; e Silva, F.C.; Chambel, M.R.; Pardos, M.; Almeida, M.H. Freezing injury in primary and secondary needles of Mediterranean pine species of contrasting ecological niches. Ann. For. Sci. 2009, 66, 407. [Google Scholar] [CrossRef]
- Xie, Y.; Pei, N.; Hao, Z.; Shi, Z.; Chen, L.; Mai, B.; Liu, Q.; Luo, J.; Luo, M.; Sun, B. Juvenile leaf color changes and physiological characteristics of Acer tutcheri (Aceraceae) during the spring season. Forests 2023, 14, 328. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Granda, E.; Scoffoni, C.; Rubio-Casal, A.E.; Sack, L.; Valladares, F. Leaf and stem physiological responses to summer and winter extremes of woody species across temperate ecosystems. Oikos 2014, 123, 1281–1290. [Google Scholar] [CrossRef]
- Adams, G.T.; Perkins, T.D. Assessing cold tolerance in Picea using chlorophyll fluorescence. Environ. Exp. Bot. 1993, 33, 377–382. [Google Scholar] [CrossRef]
- Asadi-Sanam, S.; Pirdashti, H.; Hashempour, A.; Zavareh, M.; Nematzadeh, G.A.; Yaghoubian, Y. The physiological and biochemical responses of eastern purple coneflower to freezing stress. Russ. J. Plant Physiol. 2015, 62, 515–523. [Google Scholar] [CrossRef]
- Zhang, S.R. A Discussion on chlorophyll fluorescence kinetics parameters and their significance. Chin. Bull. Bot. 1999, 16, 444. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Moustaka, J.; Tanou, G.; Giannakoula, A.; Adamakis, I.D.; Panteris, E.; Eleftheriou, E.P.; Moustakas, M. Anthocyanin accumulation in poinsettia leaves and its functional role in photo-oxidative stress. Environ. Exp. Bot. 2020, 175, 104065. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wu, F.; Zhu, X.K.; Xie, W.B. Effects of leaf types on growth and chlorophyll fluorescence characteristics in Pinus massoniana seedlings. Sci. Silvae Sin. 2019, 55, 183–192. [Google Scholar]
- Xu, Z.; Mahmood, K.; Rothstein, S.J. ROS induces anthocyanin production via late biosynthetic genes and anthocyanin deficiency confers the hypersensitivity to ROS-generating stresses in Arabidopsis. Plant Cell Physiol. 2017, 58, 1364–1377. [Google Scholar] [CrossRef]
- Wen, L.; Chen, T. Responses of pigmentation and δ 13 C in Qilian juniper to cold and drought stresses under natural conditions in the Qilian Mountains, China. Acta Physiol. Plant. 2014, 36, 2211–2218. [Google Scholar] [CrossRef]
- Guo, Z.; Cai, L.; Liu, C.; Chen, Z.; Guan, S.; Ma, W.; Pan, G. Low-temperature stress affects reactive oxygen species, osmotic adjustment substances, and antioxidants in rice (Oryza sativa L.) at the reproductive stage. Sci. Rep. 2022, 12, 6224. [Google Scholar] [CrossRef]
- Cao, P.P.; Jia, G.X. Changes of leaf color and relationship with antioxidant system in two types of Sabina vulgaris in winter. Plant Physiol. J. 2015, 51, 763–770. [Google Scholar]
- Jiang, X.R.; Peng, J.G.; Guo, L.; Gao, R.F.; Liu, Y. Relationship between winterleaf reddening and carotenoids, reactive oxygen species in Buxus microphylla L. J. Beijing For. Univ. 2015, 37, 93–99. [Google Scholar]
- Wisniewski, M.; Nassuth, A.; Teulières, C.; Marque, C.; Rowland, J.; Cao, P.B.; Brown, A. Genomics of cold hardiness in woody plants. Crit. Rev. Plant Sci. 2014, 33, 92–124. [Google Scholar] [CrossRef]
- Yu, D.J.; Hwang, J.Y.; Chung, S.W.; Oh, H.D.; Yun, S.K.; Lee, H.J. Changes in cold hardiness and carbohydrate content in peach (Prunus persica) trunk bark and wood tissues during cold acclimation and deacclimation. Sci. Hortic. 2017, 219, 45–52. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, X.T.; Wang, F.; Shao, Y.L.; Zhang, A.M.; Chang, W. The effects of chilling stress on antioxidant enzymes activities and proline, malondialdehyde, soluble sugar contents in three Paphiopedilum Species. Russ. J. Plant Physiol. 2023, 70, 61. [Google Scholar] [CrossRef]
- Yuanyuan, M.; Yali, Z.; Jiang, L.; Hongbo, S. Roles of plant soluble sugars and their responses to plant cold stress. Afr. J. Biotechnol. 2009, 8, 2004–2010. [Google Scholar]
- Wang, H.; Gong, M.; Xin, H.; Tang, L.; Dai, D.; Gao, Y.; Liu, C. Effects of chilling stress on the accumulation of soluble sugars and their key enzymes in Jatropha curcas seedlings. Physiol. Mol. Biol. Plants 2018, 24, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, H.; He, Z. Comparison of cold resistance between aboveground and underground of Sedum aizoon L. seedling with sprout under low temperature. IOP Conf. Ser. Earth Environ. Sci. 2019, 330, 032106. [Google Scholar] [CrossRef]
- Uzal, O. Effects of proline treatments on plant growth, lipid peroxidation and antioxidant enzyme activities of tomato (Solanum lycopersicum L.) seedlings under chilling stress. Gesunde Pflanz. 2022, 74, 729–736. [Google Scholar]
- Fedotova, M.V.; Dmitrieva, O.A. Proline hydration at low temperatures: Its role in the protection of cell from freeze-induced stress. Amino Acids 2016, 48, 1685–1694. [Google Scholar] [CrossRef]
- Liu, W.; Yu, K.; He, T.; Li, F.; Zhang, D.; Liu, J. The low temperature induced physiological responses of Avena nuda L., a cold-tolerant plant species. Sci. World J. 2013, 2013, 7. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).