
Transcriptomic Profiling Reveals That the Differentially Expressed PtNAC9 Transcription Factor Stimulates the Salicylic Acid Pathway to Enhance the Defense Response against Bursaphelenchus xylophilus in Pines


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
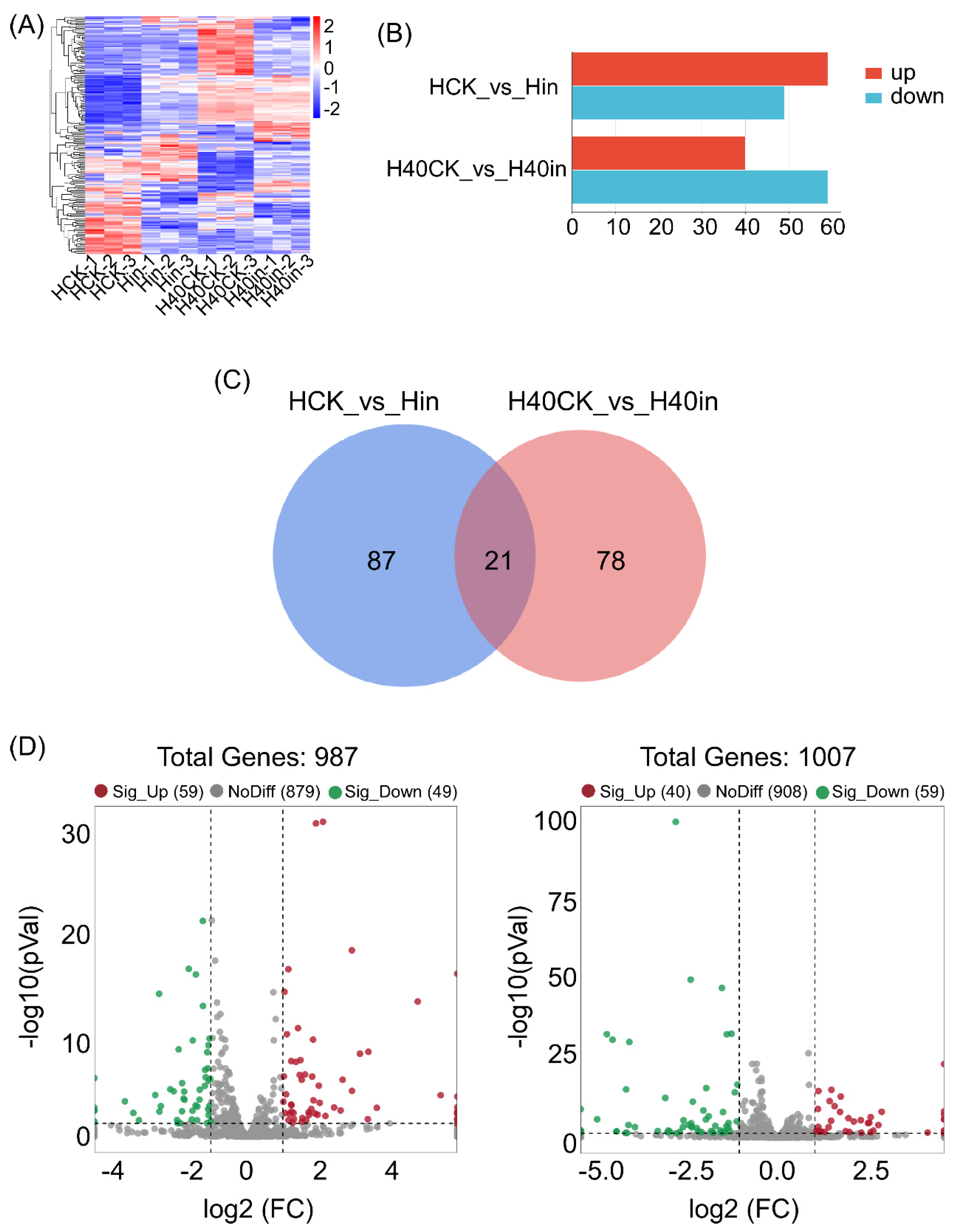
2.1. Identification of Differentially Expressed Transcription Factors (TFs)
2.2. Functional Enrichment and Annotation of TFs in Response to B. xylophilus
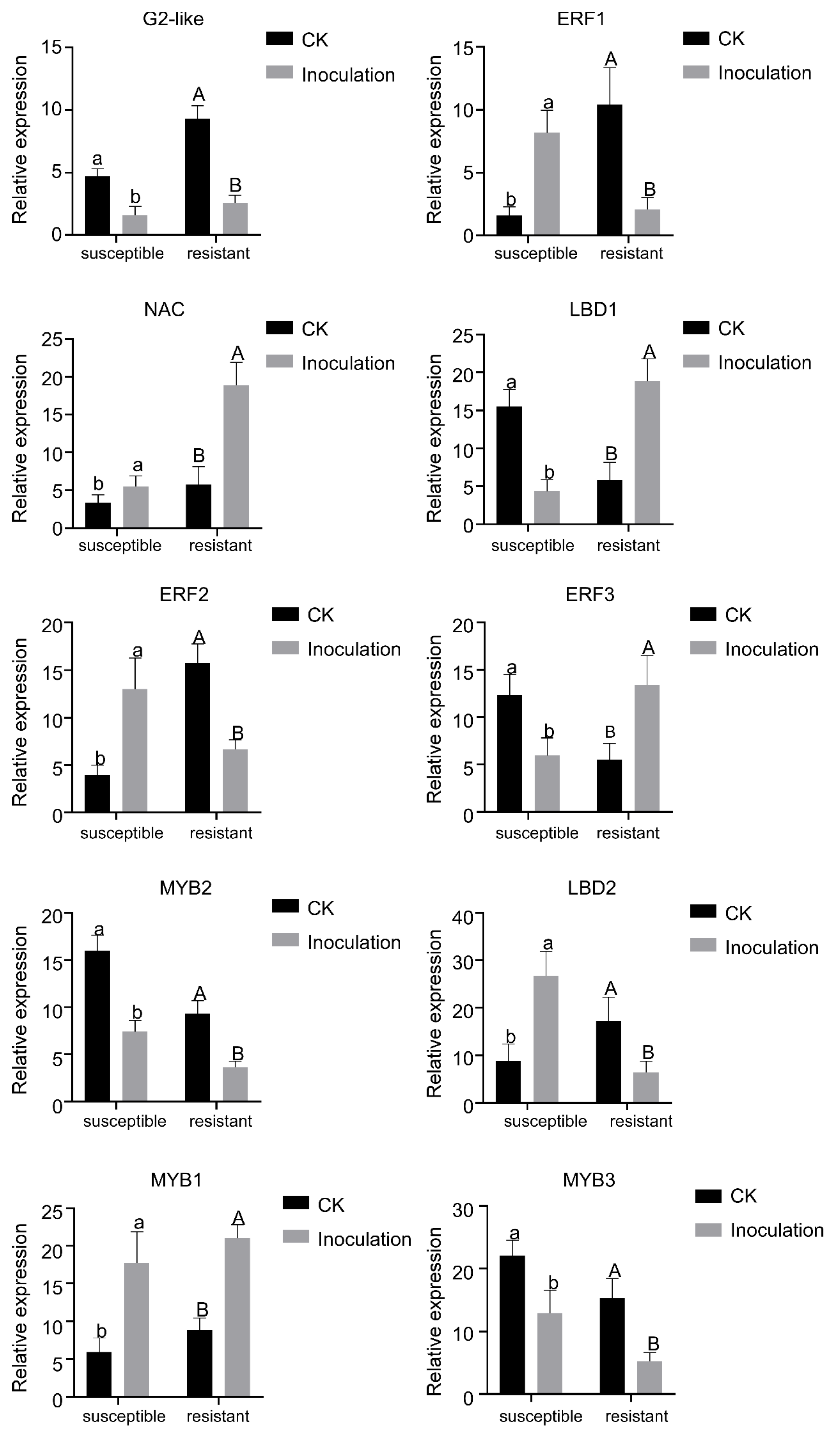
2.3. Validation of TFs by Real Time-Quantitative PCR
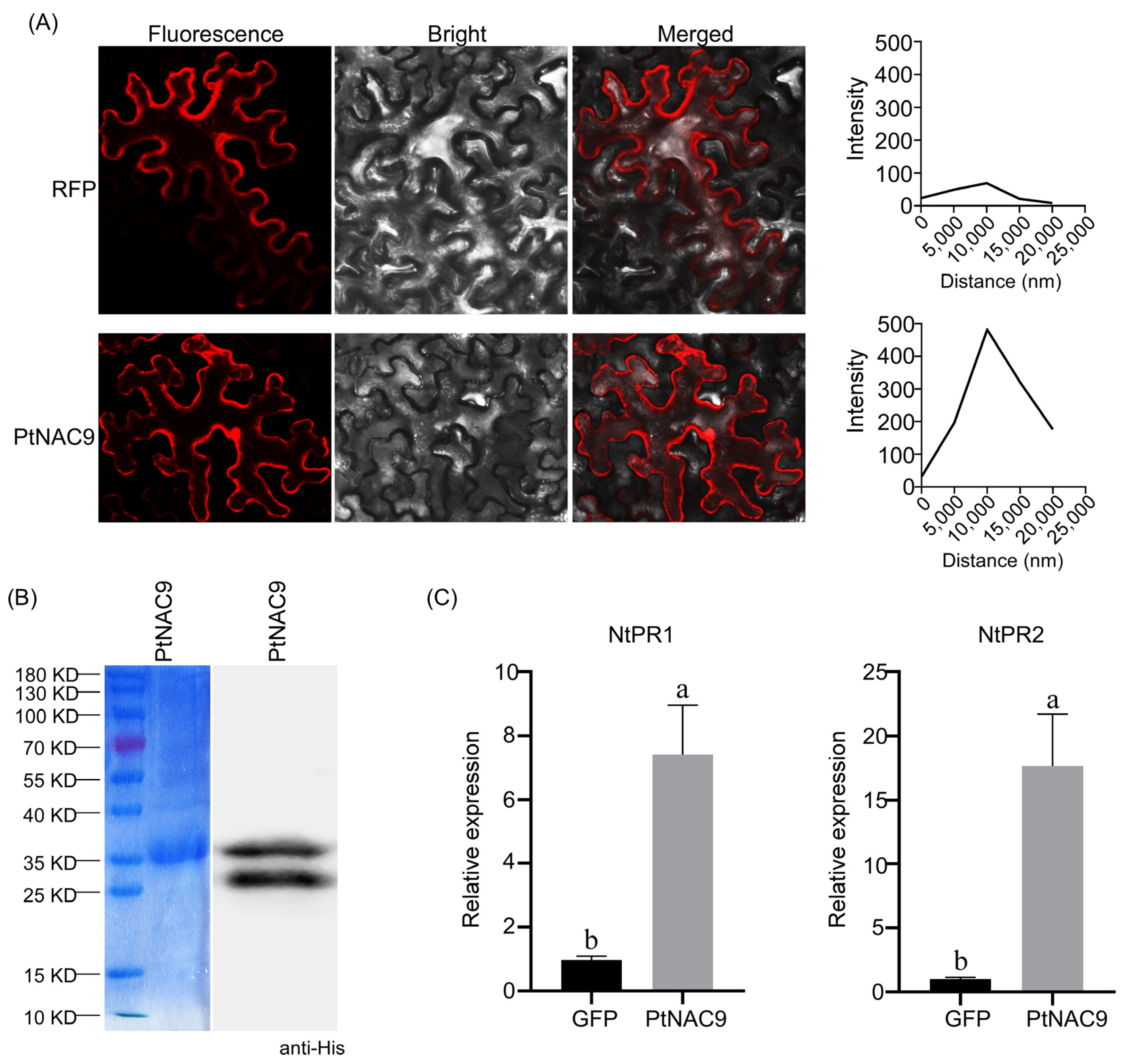
2.4. Subcellular Localization and Functional Verification of PtNAC9
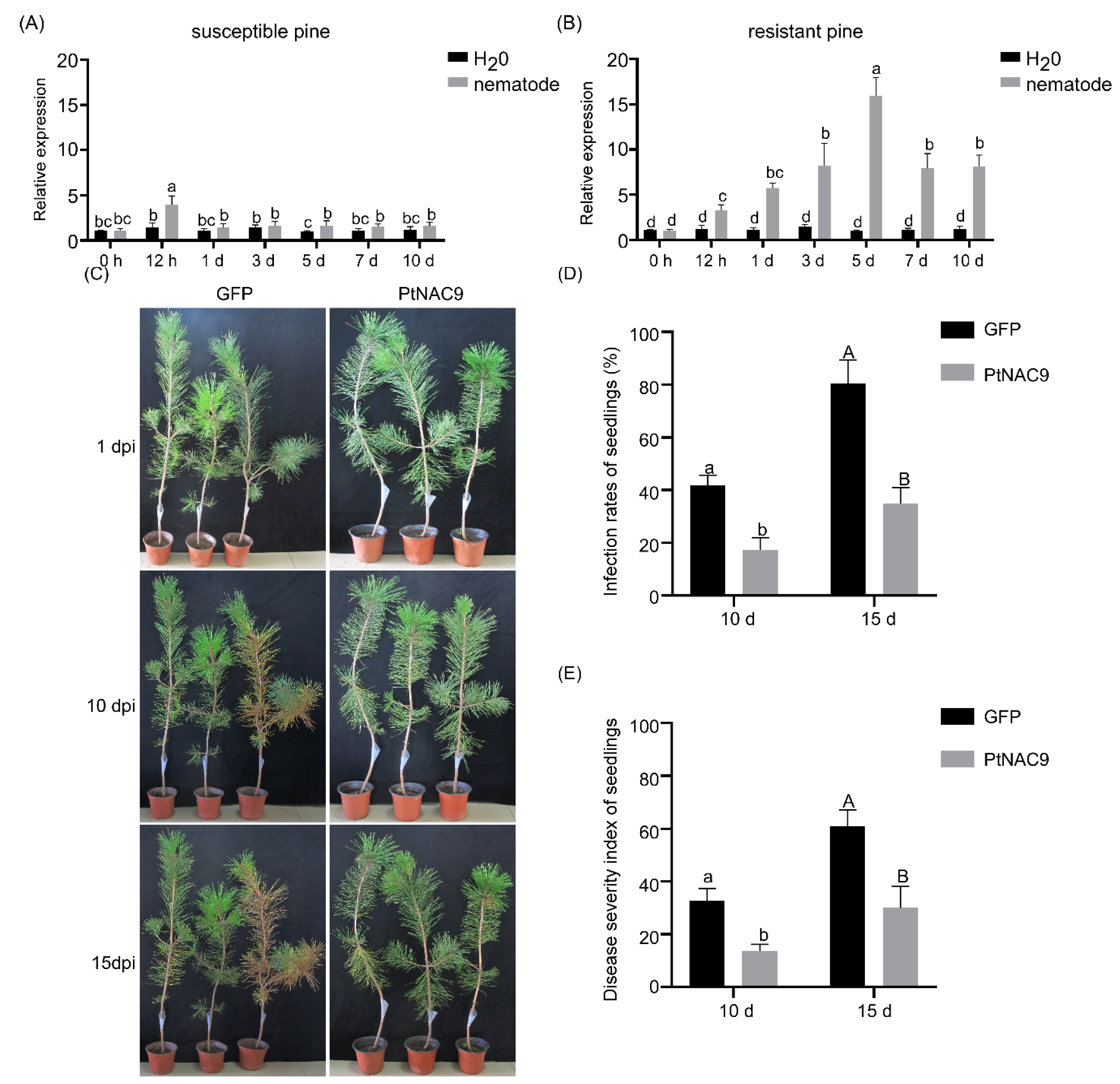
2.5. PtNAC9 Positively Regulates the Resistance of Pines to B. xylophilus
2.6. PtNAC9 Influences the Accumulation of Defense-Related Plant Hormones and Genes
3. Discussion
4. Materials and Methods
4.1. Identification of Differentially Expressed Transcription Factors (DETFs)
4.2. Functional Annotation and Pathway Analysis
4.3. Real-Time Quantitative PCR
4.4. Agrobacterium Tumefaciens Infiltration Assays
4.5. Subcellular Localization
4.6. Eukaryotic Expression of the Recombinant PtNAC9 Protein
4.7. Inoculation Assay
4.8. Determination of Salicylic Acid and Enzyme Activity in Pines
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Back, M.A.; Bonifácio, L.; Inácio, M.L.; Mota, M.; Boa, E.J.P.P. Pine wilt disease: A global threat to forestry. Plant Pathol. 2024, 73, 1026–1041. [Google Scholar] [CrossRef]
- Yang, R.; Deng, C.; Ouyang, B.; Ye, Z. Molecular analysis of two salt-responsive NAC-family genes and their expression analysis in tomato. Mol. Biol. Rep. 2011, 38, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, B.; Yu, D.; Tang, X.; Sun, J.; Zhang, B. A near-complete genome assembly of Monochamus alternatus a major vector beetle of pinewood nematode. Sci. Data 2024, 11, 312. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Khan, M.R. Pine Wood Nematode in Coniferous Forests and Their Management by Novel Biological and Biotechnological Interventions. In Novel Biological and Biotechnological Applications in Plant Nematode Management; Khan, M.R., Ed.; Springer Nature: Singapore, 2023; pp. 489–514. [Google Scholar]
- Proença, D.N.; Francisco, R.; Kublik, S.; Schöler, A.; Vestergaard, G.; Schloter, M.; Morais, P.V. The Microbiome of Endophytic, Wood Colonizing Bacteria from Pine Trees as Affected by Pine Wilt Disease. Sci. Rep. 2017, 7, 4205. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Espada, M.; Mota, M.; Nóbrega, F.; Ramos, A.P.; Inácio, M.L. Fungal communities of the pine wilt disease complex: Studying the interaction of Ophiostomatales with Bursaphelenchus xylophilus. Front. Plant Sci. 2022, 13, e908308. [Google Scholar] [CrossRef]
- Ikegami, M.; Jenkins, T.A.R. Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model-Pine Wilt disease as a model case. For. Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- Yu, X.Q.; Niu, H.Q.; Liu, C.; Wang, H.L.; Yin, W.; Xia, X. PTI-ETI synergistic signal mechanisms in plant immunity. Plant Biotechnol. J. 2024, 22, 1–16. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou BP, M.; Ding, P.; Xin, X.F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef]
- Lopes Fischer, N.; Naseer, N.; Shin, S.; Brodsky, I.E. Effector-triggered immunity and pathogen sensing in metazoans. Nat. Microbiol. 2020, 5, 14–26. [Google Scholar] [CrossRef]
- Biswas, D.; Gain, H.; Mandal, A. MYB transcription factor: A new weapon for biotic stress tolerance in plants. Plant Stress 2023, 10, 100252. [Google Scholar] [CrossRef]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors—Being in the right place with the right company. Curr. Opin. Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Chen, D. Exploring Plant Transcription Factor Regulatory Networks. In Integrative Bioinformatics: History and Future; Springer: Singapore, 2022; pp. 171–195. [Google Scholar]
- Aerts, N.; Chhillar, H.; Ding, P.; Van Wees, S.C.M. Transcriptional regulation of plant innate immunity. Essays Biochem. 2022, 66, 607–620. [Google Scholar]
- Cheng, C.; Xu, L.; Xu, D.; Lou, Q.; Lu, M.; Sun, J. Does cryptic microbiota mitigate pine resistance to an invasive beetle-fungus complex? Implications for invasion potential. Sci. Rep. 2016, 6, 33110. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wang, R.; Shi, Y.; Shen, H.; Yang, L. Reactive oxygen metabolism in the proliferation of Korean pine embryogenic callus cells promoted by exogenous GSH. Sci. Rep. 2023, 13, 2218. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Sirén, H. Bioactive phenolic compounds, metabolism and properties: A review on valuable chemical compounds in Scots pine and Norway spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Wen, T.; Feng, Y.; Zhang, Y. Transcriptomic analysis reveals differentially expressed genes associated with pine wood nematode resistance in resistant Pinus thunbergii. Tree Physiol. 2023, 43, 995–1008. [Google Scholar] [CrossRef]
- Chen, N.; Shao, Q.; Lu, Q.; Li, X.; Gao, Y.; Xiao, Q. Research progress on function of NAC transcription factors in tomato (Solanum lycopersicum L.). Euphytica 2023, 219, 22. [Google Scholar] [CrossRef]
- Li, W.; Zeng, Y.; Yin, F.; Wei, R.; Mao, X. Genome-wide identification and comprehensive analysis of the NAC transcription factor family in sunflower during salt and drought stress. Sci. Rep. 2021, 11, 19865. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Kim, S.Y.; Hyeon do, Y.; Kim, D.H.; Dong, T.; Park, Y.; Jin, J.B.; Joo, S.-H.; Kim, S.-K.; Hong, J.C.; et al. The Arabidopsis NAC transcription factor ANAC096 cooperates with bZIP-type transcription factors in dehydration and osmotic stress responses. Plant Cell. 2013, 25, 4708–4724. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Zhang, J.; Ren, Y.; Li, M.; Tian, S.; Yu, Y.; Zuo, Y.; Gong, G.; Zhang, H.; et al. The NAC transcription factor ClNAC68 positively regulates sugar content and seed development in watermelon by repressing ClINV and ClGH3.6. Hortic. Res. 2021, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Das, R.; Mandal, A.; Kundu, P. Functional characterization of tomato membrane-bound NAC transcription factors. Plant Mol. Biol. 2017, 93, 511–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiu, Z.; Zhu, W.; Wang, N.; Bai, M.; Kuang, H.; Cai, C.; Zhong, X.; Kong, F.; Lü, P.; et al. The NAC transcription factors SNAP1/2/3/4 are central regulators mediating high nitrogen responses in mature nodules of soybean. Nat. Commun. 2023, 14, 4711. [Google Scholar] [CrossRef] [PubMed]
- Meisrimler, C.N.; Pelgrom AJ, E.; Oud, B.; Out, S.; Van den Ackerveken, G. Multiple downy mildew effectors target the stress-related NAC transcription factor LsNAC069 in lettuce. Plant J. 2019, 99, 1098–1115. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests 2023, 14, 174. [Google Scholar] [CrossRef]
- Zhou, H.; Yuan, X.; Zhou, H.; Shen, H.; Ma, L.; Sun, L.; Fang, G.; Sun, H. Surveillance of pine wilt disease by high resolution satellite. J. For. Res. 2022, 33, 1401–1408. [Google Scholar] [CrossRef]
- Zuccaro, A.; Langen, G. Breeding for resistance: Can we increase crop resistance to pathogens without compromising the ability to accommodate beneficial microbes? New Phytol. 2020, 227, 279–282. [Google Scholar] [CrossRef]
- Wu, X.Q.; Zhang, Y.; Chen, W.S. Resistance and histopathological observation of 13 resistant families of Pinus thunbergii to Bursaphelenchus xylophilus. Acta Phytopathol. Sin. 2008, 38, 44–50. [Google Scholar]
- Zhang, Y.; Wu, X.Q.; Wang, J. Determination and histopathological observation of resistance to PWD in Pinus densiflora. J. Nanjing Univ. (Nat. Sci. Ed.) 2007, 31, 110–114. [Google Scholar]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Lu, J.; Wu, T.; Zhang, B.; Liu, S.; Song, W.; Qiao, J.; Ruan, H. Types of nuclear localization signals and mechanisms of protein import into the nucleus. Cell Commun. Signal. 2021, 19, 60. [Google Scholar] [CrossRef]
- Liu, B.; Ouyang, Z.; Zhang, Y.; Li, X.; Hong, Y.; Huang, L.; Liu, S.; Zhang, H.; Li, D.; Song, F. Tomato NAC Transcription Factor SlSRN1 Positively Regulates Defense Response against Biotic Stress but Negatively Regulates Abiotic Stress Response. PLoS ONE 2014, 9, e102067. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated NAC transcription factor (SlNAC35) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant. 2016, 158, 45–64. [Google Scholar] [CrossRef]
- Anisimova, O.K.; Shchennikova, A.V.; Kochieva, E.Z.; Filyushin, M.A. Pathogenesis-Related Genes of PR1, PR2, PR4, and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021, 22, 6688. [Google Scholar] [CrossRef]
- Molinari, S.; Fanelli, E.; Leonetti, P. Expression of tomato salicylic acid (SA)-responsive pathogenesis-related genes in Mi-1-mediated and SA-induced resistance to root-knot nematodes. Mol. Plant Pathol. 2014, 15, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Bagal, U.R.; Leebens-Mack, J.H.; Lorenz, W.W.; Dean, J.F.D. The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage. BMC Genom. 2012, 13, S1. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.-Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef]
- Wang, Q.; Han, C.; Ferreira, A.O.; Yu, X.; Ye, W.; Tripathy, S.; Kale, S.D.; Gu, B.; Sheng, Y.; Sui, Y.; et al. Transcriptional Programming and Functional Interactions within the Phytophthora sojae RXLR Effector Repertoire. Plant Cell. 2011, 23, 2064–2086. [Google Scholar] [CrossRef]
- Hu, B.; Sakakibara, H.; Kojima, M.; Takebayashi, Y.; Bußkamp, J.; Langer, G.J.; Peters, F.S.; Schumacher, J.; Eiblmeier, M.; Kreuzwieser, J.; et al. Consequences of Sphaeropsis tip blight disease for the phytohormone profile and antioxidative metabolism of its pine host. Plant Cell. Environ. 2018, 41, 737–754. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, T.-Y.; Wang, X.-Y.; Wu, X.-Q.; Ye, J.-R. Transcriptomic Profiling Reveals That the Differentially Expressed PtNAC9 Transcription Factor Stimulates the Salicylic Acid Pathway to Enhance the Defense Response against Bursaphelenchus xylophilus in Pines. Forests 2024, 15, 1538. https://doi.org/10.3390/f15091538
Wen T-Y, Wang X-Y, Wu X-Q, Ye J-R. Transcriptomic Profiling Reveals That the Differentially Expressed PtNAC9 Transcription Factor Stimulates the Salicylic Acid Pathway to Enhance the Defense Response against Bursaphelenchus xylophilus in Pines. Forests. 2024; 15(9):1538. https://doi.org/10.3390/f15091538
Chicago/Turabian StyleWen, Tong-Yue, Xin-Yu Wang, Xiao-Qin Wu, and Jian-Ren Ye. 2024. "Transcriptomic Profiling Reveals That the Differentially Expressed PtNAC9 Transcription Factor Stimulates the Salicylic Acid Pathway to Enhance the Defense Response against Bursaphelenchus xylophilus in Pines" Forests 15, no. 9: 1538. https://doi.org/10.3390/f15091538
APA StyleWen, T.-Y., Wang, X.-Y., Wu, X.-Q., & Ye, J.-R. (2024). Transcriptomic Profiling Reveals That the Differentially Expressed PtNAC9 Transcription Factor Stimulates the Salicylic Acid Pathway to Enhance the Defense Response against Bursaphelenchus xylophilus in Pines. Forests, 15(9), 1538. https://doi.org/10.3390/f15091538