Abstract
The dry forests of southern Ecuador are characterized by a great floristic variety and high endemism rates in different taxonomic groups; however, these forests are disappearing at an alarming rate due to anthropogenic disturbances. Epiphytic cryptogams (bryophytes and lichens) are important in seasonally dry tropical forests (SDTFs) and, due to their physiology (poikilohydric species), are sensitive to the changes associated with specific human disturbances. We examined the response of epiphytic cryptogams to habitat disturbance in SDTFs (natural and semi-natural forests) by registering the frequency and coverage of epiphytic cryptogams on 182 trees. We recorded a total of 53 epiphytic cryptogams (47 lichens and 6 bryophytes). A greater number of species was found in the natural forest, with 51 species, than the semi-natural forests (DFs), which had 45 species. We found that total richness tended to decrease with increased intensity of disturbance. The impoverishment was particularly related to the reduction in tree density and diversity (e.g., selective logging) in semi-natural compared to natural forests. However, the epiphytic composition showed little relationship with forests disturbance. We conclude that anthropic disturbance, particularly selective logging, are the main factors affecting the diversity of epiphytic cryptogams in the SDTFs from southern Ecuador.
1. Introduction
Tropical dry forests are found from sea level to approximately 2000 m a.s.l.; in southwestern Ecuador and northwestern Peru, they are part of the Tumbesian biogeographic region of endemism [1,2]. These forests are characterized by their richness in plant and animal species, being a refuge for an extraordinary diversity of endemic biota [1]. Tropical dry forests have a smaller stature and lower basal area compared to humid tropical forests [3].
These forests are subject to severe climatic seasonality, marked by a dry period that lasts 5 to 6 months a year [2]. Therefore, ecological processes are strongly seasonal and net primary productivity is lower than in tropical forests since growth only takes place during the wet season [4]. Despite evidence of extreme threats and losses to which they are exposed [5], neotropical dry forests have received less attention than more humid tropical ecosystems [6]. Less than 38% of the remnants of these dry forests currently benefit from some form of protection [6].
Tropical dry forests are disappearing at an alarming rate and, therefore, they have been recognized as one of the most threatened ecosystems in the world, due to anthropogenic disturbance [5,6]. They have experienced significant deforestation; much of their original coverage being converted to pastures and croplands used for timber extraction, firewood collection, and cattle grazing [7,8].
Tropical dry forests in southern Ecuador are known for their high degree of endemism in various taxonomic groups [1,9], which are exposed to multiple threats such as the expansion of agricultural frontiers, selective harvesting, and overgrazing and trampling by cattle and goats [10,11], with negative effects on regeneration and subsequent forest functioning determining a decline in biodiversity and ecosystem services [12]. In this context, several previous studies evaluated the influence of environmental conditions or anthropogenic disturbances on plant [13,14] and animal [15] diversity in this ecosystem. However, little attention has been paid to epiphytic cryptogam communities (i.e., bryophytes and lichens) [16].
Epiphytes are one of the most diverse forms of plant life, whose species richness peaks in the tropics and subtropics, including seasonally dry tropical forests [16,17]. Approximately 10% of all vascular plant species worldwide are epiphytes [18]. Bryophytes and lichens constitute a major fraction of epiphytic organisms [19,20]. They are generally sensitive to forest disturbance [16,21], and their poikilohydric physiology is strongly coupled to moisture and light regimes [22,23]. However, most studies on forest disturbance focus on vascular epiphytes [24,25], and few have focused on epiphytic lichens and bryophytes [16,17,26]. Forest disturbance affects epiphyte diversity in dry forests [21]. For instance, Benítez et al. [16] found a reduction in lichen epiphytes caused by the removal of host trees that harbor high lichen species richness. Werner and Gradstein [21] found that forest disturbance caused severe loss of epiphytic bryophyte diversity. As a general pattern, forest logging produces changes microclimate, for instance, humidity and light availability, which adversely affect diversity [25,27,28]. Additionally, other factors such as the diversity of trees [16,29,30], tree size, the texture and chemical properties of the bark, and the species of the host tree [31,32] play important roles in cryptogam communities in tropical dry forests.
This study aimed to compare the richness and composition of epiphytic lichens and bryophytes in natural versus semi-natural seasonally dry tropical forests. We hypothesized that differences in diversity and composition would be related with the degree of disturbance. Another objective was to determine the effects of tree size, aspect and slope on the diversity of epiphytic lichens and bryophytes. Specifically, we examined the following question: do epiphytic cryptogams (lichens and bryophytes) suffer a reduction in richness as forest disturbance increases in tropical dry forests? Which factors contribute most to differences in epiphytic cryptogams among the two forests disturbance levels?
2. Methods
2.1. Study Area
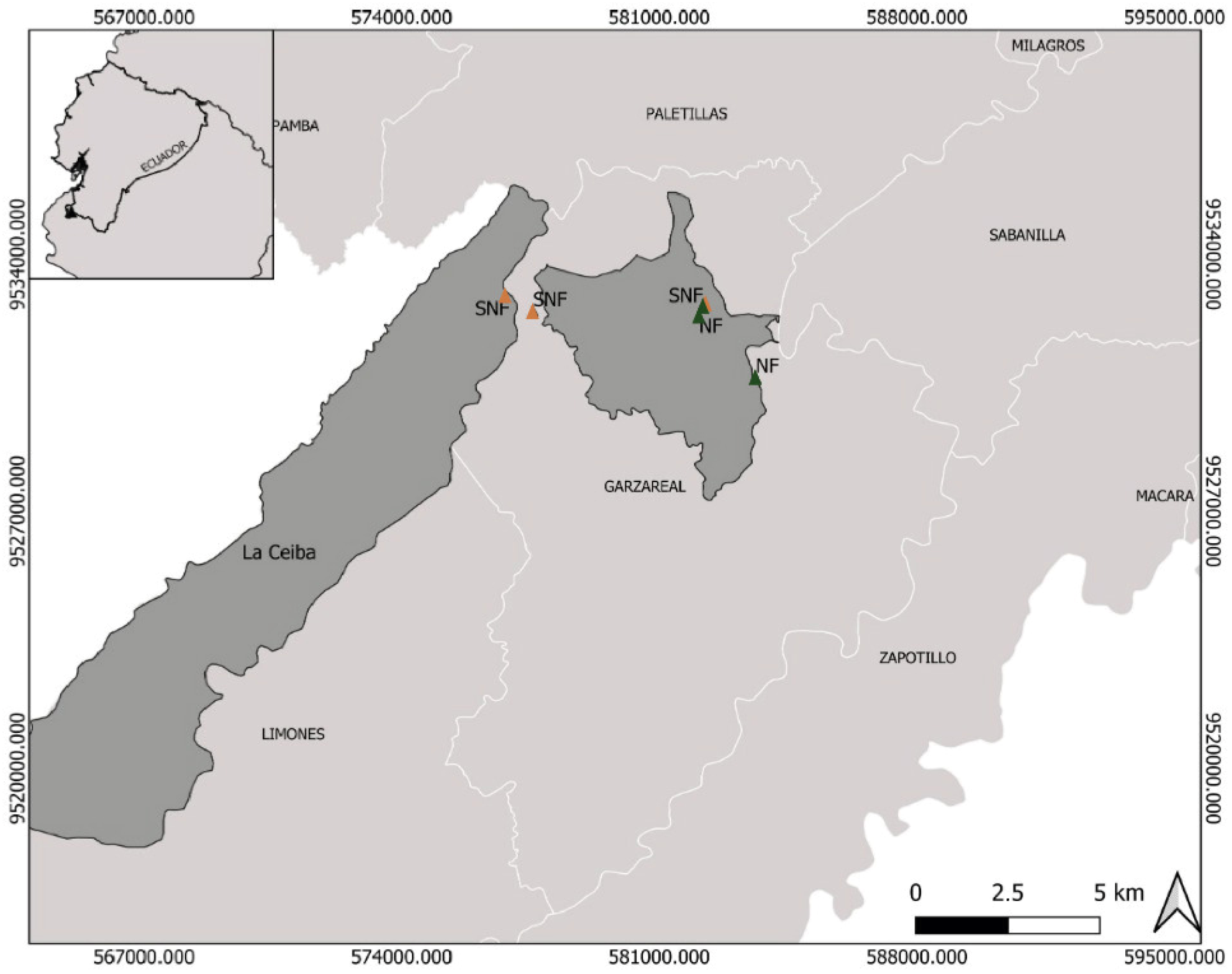
The study area included tropical dry forests located in southern Ecuador (Loja Province; 200–500 m a.s.l) (Figure 1). The mean annual temperature varies between 18 and 26 °C and annual rainfall is ca. 630–1300 mm [33]. We selected six sampling sites, each represented by one of two levels of forest disturbance following a standardized protocol related to vegetation density and reflectance levels [15,34]. The authors identified two types of vegetation based on the level of disturbance:
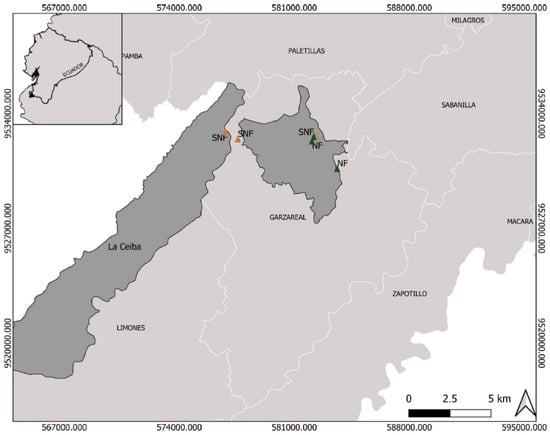
Figure 1.
Study area in Zapotillo, Loja Province, southwestern Ecuador, showing the location of the six tropical dry forest sites. Natural forest (NF) and semi-natural forest (SNF).
Dense or natural (low anthropogenic pressure): These forests are characterized by a dense canopy layer (ca. 50%–70% coverage), with large trees. The upper canopy is composed of Handroanthus chrysanthus (Jacq.) S.O. Grose, Eriotheca ruizii (K. Schum.) A. Robyns., Bursera graveolens (Kunth) Triana & Planch., Cordia macrantha Chodat., Simira ecuadorensis (Standl.) Steyerm, and Piscidia carthagenensis Jacq.
Semi-natural (moderate anthropogenic pressure): These forests have discontinuous fragments as a result of disturbance, with the presence of dominant isolated trees [15,34]. The canopy layer covers about 40%–50% of the area and is primarily composed of Handroanthus chrysanthus and Cordia macrantha species.
2.2. Sampling Design and Data Collection
Three forests (Table 1) were analyzed for each disturbance category (natural and semi-natural). Six plots (10 × 10 m) were selected within each forest, and five trees were sampled within each of the six plots. We sampled the presence/absence and cover of epiphytic cryptogams using 10 × 60 cm and 20 × 30 cm quadrants for shrubs and trees, respectively, for a total of 180 tree trunks. Samples were taken on each tree at 50–100 cm of height on the northern and southern exposure following general protocols for sampling bryophytes and lichens in dry forests [16]. In addition, we measured the elevation (m a.s.l.), slope (°), aspect (cosine transformed), and tree diameter (cm) for all trees and shrubs having a diameter above 7 cm as a proxy for the stand forest structure. The total species richness was defined as the number of species identified within the two grids on each tree. The identification of lichens and bryophytes specimens involved the use of various published keys. To identify lichens, we used conventional microscopy methods and performed spot tests, such as ultraviolet (UV) light and reactions with potassium hydroxide (K), commercial bleach (C), and Lugol’s solution (I). For lichen species nomenclature, we consulted MycoBank (www.mycobank.org/, accessed on 3 September 2024) and LIAS 1995–2016 (A Global Information System for Lichenized and Non-Lichenized Ascomycetes: www.lias.net, accessed on 3 September 2024). For bryophytes, we consulted the Liverworts and Hornworts of Colombia and Ecuador and the Catalogue of the Plants and Lichens of Colombia. The lichen not being resolved to the species level (e.g., Lecanora, Phyllopsora, Rinodina) represents a single taxonomically unresolved species observed multiple times. All specimens were deposited in the Bryophytes and Lichens Collection in the herbarium HUTPL.

Table 1.
Averages of the environmental variables in the natural and semi-natural dry forests studied. MTD = mean tree diameter.
2.3. Data Analyses
We assessed richness (S), Shannon’s diversity (H’), and Simpson’s diversity (D) for cryptogams at tree level. The effects of disturbance, trunk diameter (cm), slope (°), and aspect (cosine transformed) on species richness of bryophytes and lichens were analyzed using generalized linear models (GLMs) [35]. Generalized linear models were fitted assuming a Poisson error distribution for count data (richness) and gamma error distribution for continuous data (diversity indices). Parameters for the GLMs were estimated using Laplace approximations [36].
Non-metric multidimensional scaling (NMDS) was conducted to identify the main factors affecting epiphytic composition. We calculated the resemblance matrix between trees using Bray–Curtis dissimilarity distance. The findings were then visualized in an NMDS ordination diagram. To test whether the two levels of disturbance had significantly different epiphytic species compositions and to examine the effects of forest type, trunk diameter (cm), slope (°), and aspect (cosine-transformed), we conducted a two-factor permutational multivariate analysis of variance (PERMANOVA) [37]. The experimental design in this analysis involved two factors: disturbance (2 levels, fixed factor) and forests (6 levels, random nested within disturbance), with trees serving as replicates (n = 182). The cover data, representing the percentage cover of each lichen and bryophyte per tree, were log10(x + 1) transformed to account for the contributions of both rare and abundant taxa. Multivariate analysis was performed with package ‘vegan’ [38] in the R environment [39].
3. Results
A total of 53 epiphytic cryptogams (47 lichens and 6 bryophytes) were collected from 182 trees (Table 2). The natural forest exhibited the highest number of species, with 51 species observed, while the semi-natural forests had 45 species.
Table 2.
Number of trees on which each epiphytic cryptogam species appears, according to the forest type. * = denote exclusive species per forest type.
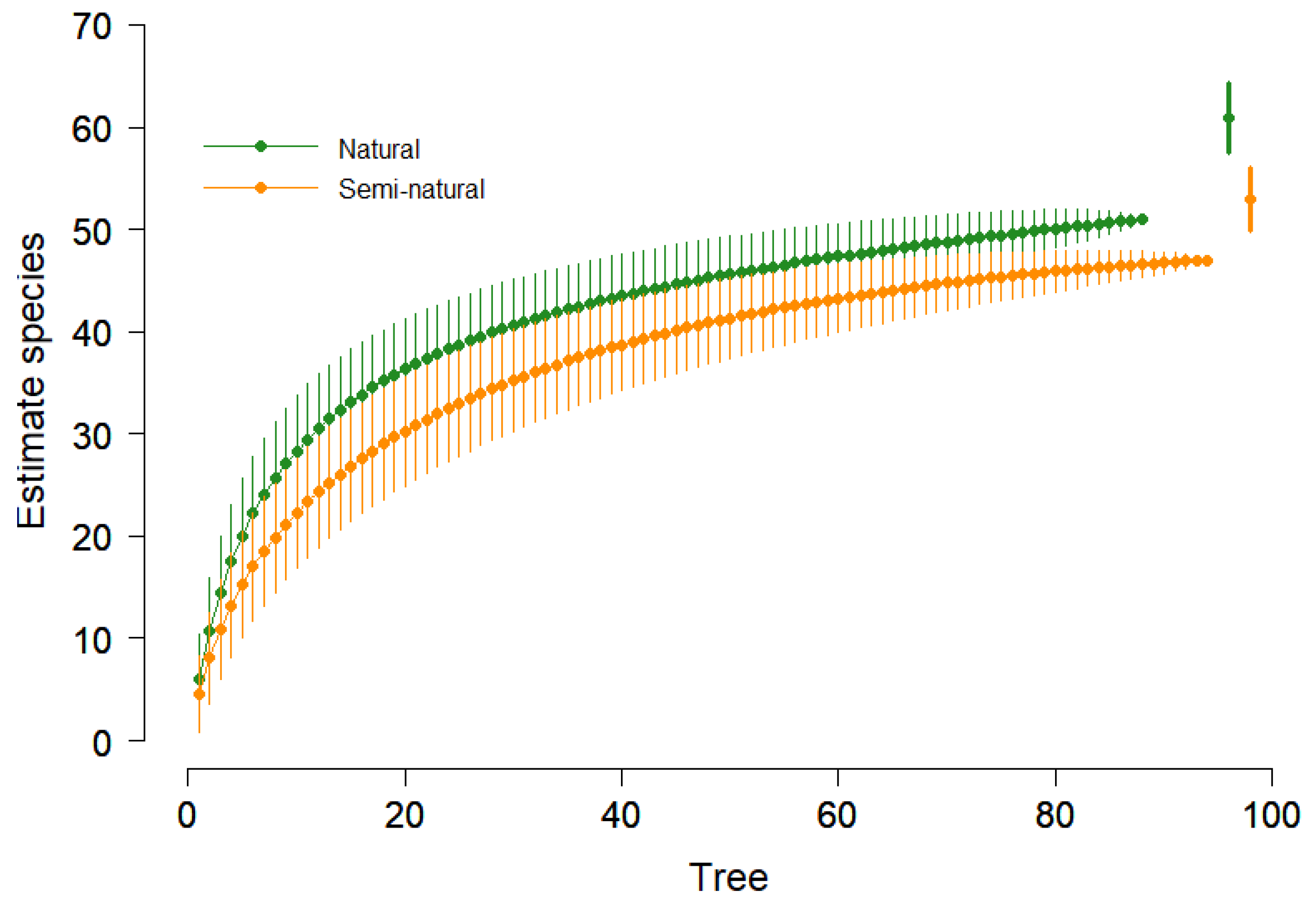
A similar trend was observed using the richness estimator (Chao 2), confirming that natural forests exhibited higher species richness than semi-natural forests (Figure 2).
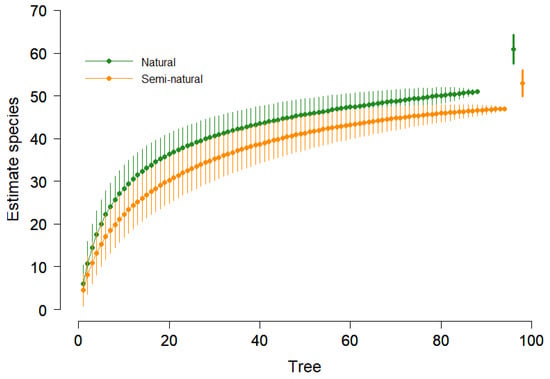
Figure 2.
Species rarefaction curve with 95% confidence interval and Chao 2 estimator (points on right) for two forest types (Natural and Simi-natural).
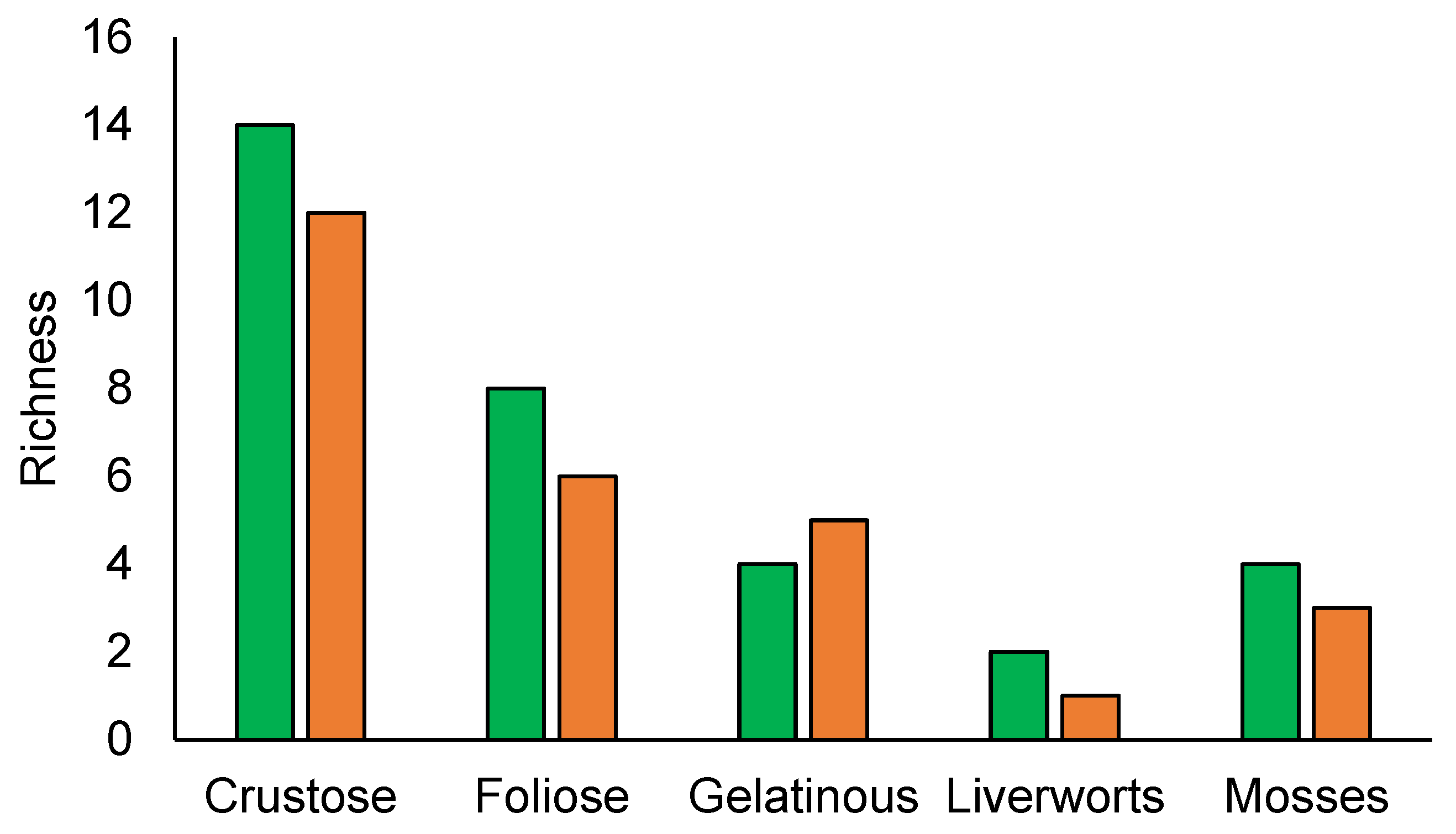
Natural and semi-natural forests lichen composition were dominated by crustose lichens followed by foliose and gelatinose species, while for bryophytes mosses have more species than liverworts (Figure 3).
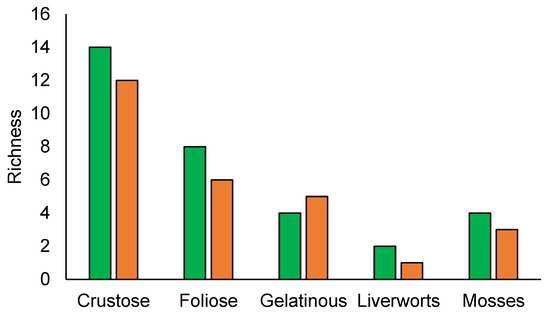
Figure 3.
Species richness by growth forms. Natural forest (green), semi-natural forest (orange).
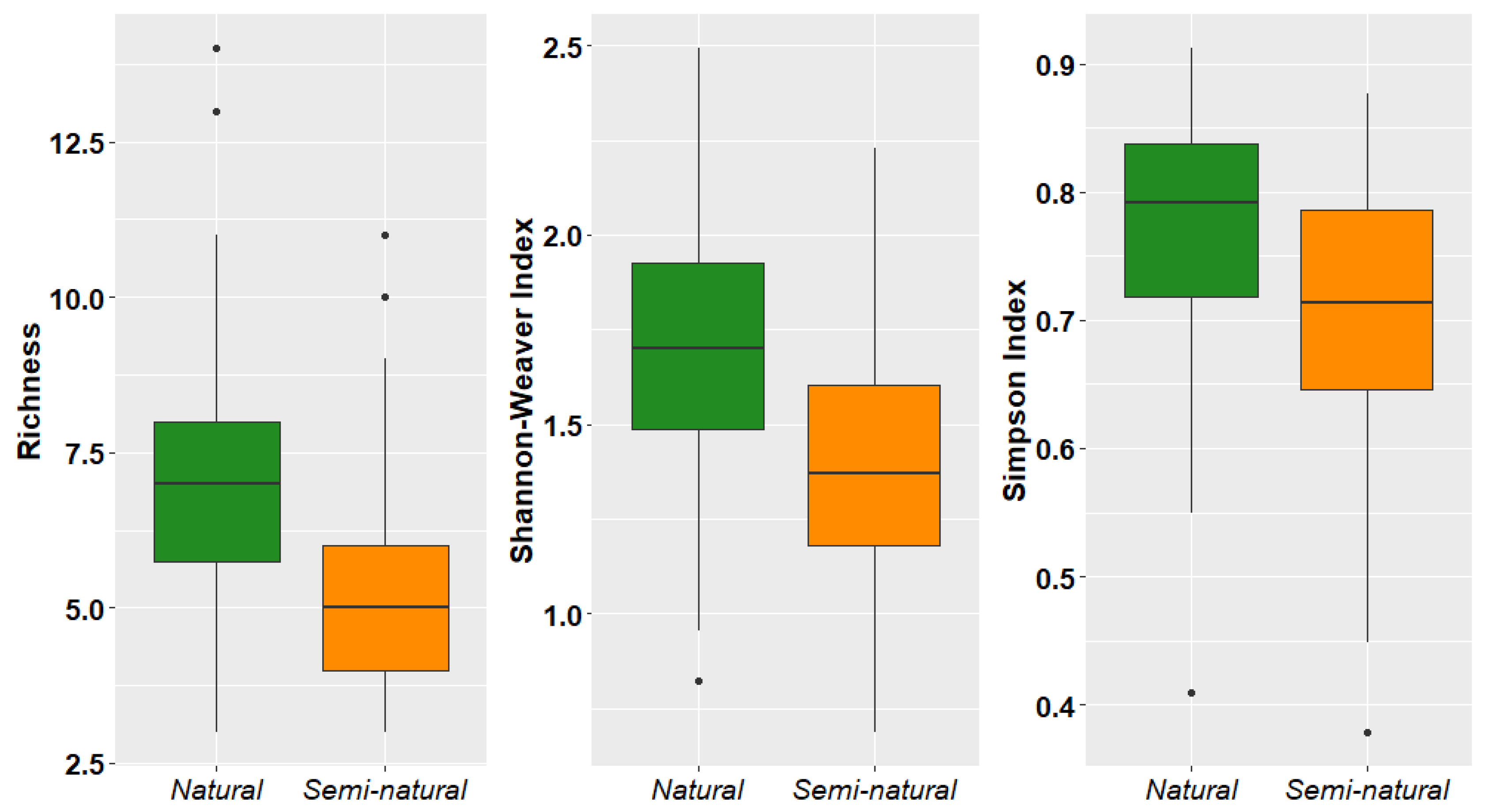
Both richness and diversity of epiphytic lichens and bryophytes (Shannon–Weaver and Simpson indices) were higher in natural forests than semi-natural forests (Figure 4).
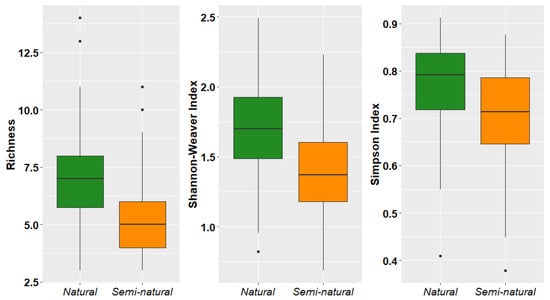
Figure 4.
Species richness and diversity of epiphytic lichens and bryophytes in two dry forest types from southwestern Ecuador.
Analysis of environmental variables showed that disturbance significantly and negatively impacted species richness and diversity at the tree level, while DBH negatively influenced epiphyte diversity (Table 3).
Table 3.
Results of the generalized linear models on community traits at tree level. SE = standard error, Z = Z value. p-values < 0.05 are considered significant.
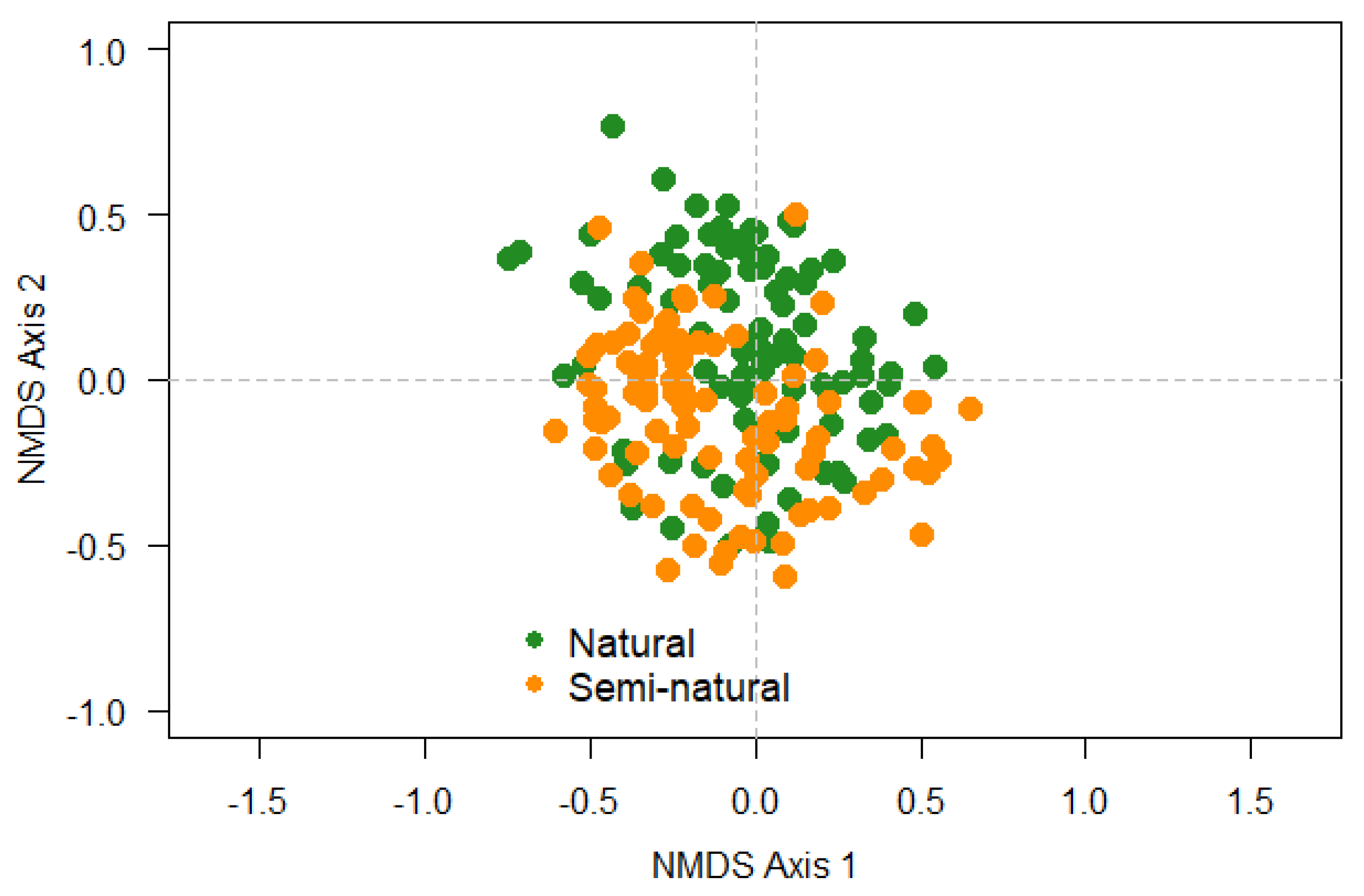
The NMDS analysis revealed that there are no clear differences in community composition of epiphytic cryptogams (lichen and bryophyte) between natural and semi-natural forests (Figure 5).
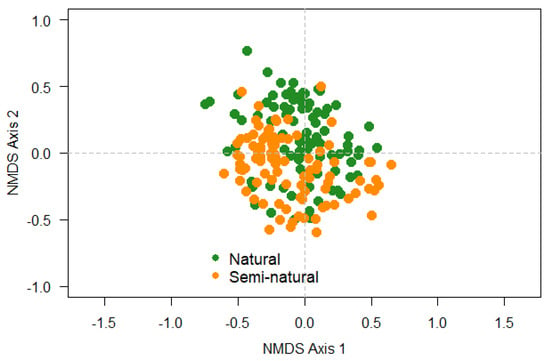
Figure 5.
Non-metric multidimensional scaling analysis of species composition for the samples (host trees) in the studied dry forests (two stands of each forest type) from Ecuador.
The multivariate statistical analyses showed that epiphytic composition of cryptogams was structured according to disturbance, this factor explaining 4% of total variance (Table 4).
Table 4.
Results of two-factor PERMANOVA analysis of species composition. df = degrees of freedom; SS = sum of squares; MS = median squares, F = F-statistic; R2 = coefficient of variation.
4. Discussion
Our results indicate that forest disturbance is an important determinant for cryptogam epiphytic diversity on trunk bases in tropical dry forests of southern Ecuador, having a negative impact on epiphytic lichen diversity (richness and diversity indices), related to changes in forest structure and the removal of host trees. These findings are consistent with studies that used different sampling protocols of lichens and bryophytes, for instance Werner and Gradstein [21], who sampled the diversity and distribution of epiphyte (bryophytes) species across the entire vertical profile, from the base to the treetop and showed that epiphytic cryptogams are sensitive indicators of human disturbance in montane dry forests dominated by Vachellia macracantha (Humb. & Bonpl. ex Willd.) Seigler & Ebinger, as host tree. Similarly, Benítez et al. [16] showed that epiphytic lichens on trunk bases are indicators of human disturbance in the seasonally dry tropical forests (SDTFs) of Ecuador.
The low bryophyte richness (4 mosses and 2 liverworts) in our study is congruent with Werner and Gradstein [21] who reported 13 species of bryophytes, with 9 mosses and 4 liverworts in montane dry forests of Ecuador. Also, only one bryophyte species (Myriocoleopsis minutissima subsp. minutissima) was recorded on 513 trees of seasonally tropical dry forests in Ecuador. Humidity is a key factor for diversity and distribution of bryophyte in tropical montane and dry forests [32,40]. On the other hand, the high lichen richness (47 species) is congruent with Benítez et al. [18] who reported 122 species of lichens in seasonally dry tropical forests of Ecuador. In addition, Lücking et al. [17] reported 61 lichens in seasonally dry tropical forest remnants from Colombia. Following this pattern, García-Martínez and Mercado-Gómez [41] reported 92 species of lichens in a tropical dry forest in the Montes de María, Sucre in Colombia. Thus, tropical dry forests from Colombia and Ecuador showed high values of lichen richness, in particular crustose species.
Cryptogam richness showed greater sensitivity to habitat disturbance. The negative association between richness and disturbance was likely the result of reduced tree density and diversity in semi-natural compared to natural forests after years of selective logging, agriculture, and livestock grazing [15,42]. Benítez et al. [16] and Déleg et al. [43] have shown that tree diversity is a key factor for epiphytic cryptogam richness in tropical lowland rainforests and seasonally dry tropical forests, respectively. This result emphasizes the importance of natural forests, characterized by a relatively high tree density and diversity, for the preservation of cryptogamic biodiversity in tropical dry forests. On the other hand, the diversity of cryptogams (Shannon–Weaver and Simpson indices) at tree level declined with increased tree diameter. Similarly, Benítez et al. [16] shown that large trees in the undisturbed forests had a low diversity of lichens.
Epiphytic cryptogam species composition was not affected by forest disturbance, showing a greater resilience. For lichens and bryophytes, thallus morphology and growth-form types are optimized for the uptake and loss of water [19,44], and therefore the environmental conditions of a specific forests will affect the presence of different morphological types [45]. For instance, Arthonia cinnabarina, Candelaria concolor, Dirinaria picta, Graphys ruiziana, Parmotrema tinctorum and Parmotrema arnoldii were distributed in both natural and semi-natural forests. These species are indicators of drier sites, thriving under higher solar radiation and lower humidity [16,32]. On the other hand, the frequency of occurrence of some species shows the opposite pattern, increasing in semi-natural forests (e.g., Anisomeridium albisedum, Lepraria sp., Leptogium milligranum, Peltula steppae Kalb, and Physcia crispa Nyl). This is likely due to increased light availability and reduced humidity caused by more canopy openness in semi-natural forests. Also, foliose lichens with narrow and broad lobes (e.g., Candelaria; Dirinaria, Parmotrema, Physcia, and Pyxine) that produce acids as secondary compounds were common in natural and semi-natural forests benefiting from the protection against excessive radiation conferred by these compounds [46,47]. Following this pattern, most bryophyte species were represented by sun epiphytes or drought-tolerant species (e.g., Frullania ericoides) in natural and semi-natural forests. Several studies show that species of the Frullania genera and Fabronia ciliaris are restricted to disturbed habitats and they are able to tolerate a high light intensity and low humidity [32,40,48]. However, the liverwort Myriocoleopsis minutissima subsp. minutissima has been reported in undisturbed tropical dry forest in Ecuador [16,21] and the moss Tricherpodium beccarii in tropical dry forest in Argentina [49], so this species can be catalogued as a sensitive/tolerant species.
5. Conclusions
We concluded that tropical forest disturbance significantly and drastically reduces cryptogamic species richness and diversity, primarily due to changes in forest structure and the removal of host trees, which impacts tree diversity and density. The use of epiphytic cryptogams (bryophytes and lichens) will provide an important and promising approach to assess disturbances in tropical dry forests, due to that disturbance of dry forests affects the richness and diversity of lichens and bryophytes (undisturbed forests have greater diversity than disturbed forests), thus can be use as indicators and thus have data to establish conservation measures for dry forests.
Author Contributions
Conceptualization, Á.B., E.G.-M., J.O., D.M.-A. and A.B.; methodology, Á.B., J.O., D.M.-A. and A.B.; formal analysis, Á.B.; investigation, Á.B., J.O., D.M.-A. and A.B.; writing—original draft preparation, Á.B., J.O., D.M.-A., A.B., F.L., E.Y.-S. and E.G.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “Universidad Técnica Particular de Loja”.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank the Ministerio del Ambiente del Ecuador for granting access to the field sites and the necessary collecting permits. Special thanks to Diana Szekely for proofreading the English text.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Best, B.J.; Kessler, M. Biodiversity and Conservation in Tumbesian Ecuador and Peru; BirdLife International: Cambridge, UK, 1995; Volume 218. [Google Scholar]
- Espinosa, C.I.; Cabrera, O.; Luzuriaga, A.L.; Escudero, A. What factors affect diversity and species composition of endangered Tumbesian dry forests in Southern Ecuador? Biotropica 2011, 43, 15–22. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Ecology of Tropical Dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Pennington, R.T.; Prado, D.E.; Pendry, C.A. Neotropical seasonally dry forests and Quaternary vegetation changes. J. Biogeogr. 2000, 27, 261–273. [Google Scholar] [CrossRef]
- Janzen, D.H. Tropical dry forests. Biodiversity 1988, 15, 130–137. [Google Scholar]
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A global overview of the conservation status of tropical dry forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Fajardo, L.; Gonzalez, V.; Nassar, J.M.; Lacabana, P.; Portillo, Q.C.A.; Carrasquel, F.; Rodriguez, J.P. Tropical Dry Forests of Venezuela: Characterization and Current Conservation Status 1. Biotrop. Biol. Conserv. 2005, 37, 531–546. [Google Scholar] [CrossRef]
- Leal-Pinedo, J.M.; Linares-Palomino, R. The dry forests of the Biosphere Reserve of Northwestern (Peru): Tree diversity and conservation status. Caldasia 2005, 27, 195–211. [Google Scholar]
- Aguirre Mendoza, Z.; Buri Sivisaca, D.; Geada López, G.; Betancourt Figueras, Y. Composición florística, estructura y endemismo en una parcela permanente de bosque seco en Zapotillo, provincia de Loja, Ecuador. Arnaldoa 2014, 21, 165–178. [Google Scholar]
- Paladines, R. Propuesta de conservación del bosque seco en el Sur de Ecuador. Lyonia 2003, 4, 183–186. [Google Scholar]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; de la Cruz, M. Deforestation and forest fragmentation in South Ecuador since the 1970s—Losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef]
- Patiño, J.; Ramón, P.; Gusmán-Montalván, E.; Escudero, A.; de la Cruz, M. Fencing promotes fast recovery of demographic processes after grazing-driven collapse in Bursera graveolens forests. For. Ecol. Manag. 2021, 499, 119592. [Google Scholar] [CrossRef]
- Cuevas-Vargas, H.; Cortés-Palacios, H.A. Efectos de la estructura de capital en la innovación. Investig. Adm. 2020, 49, 126. [Google Scholar]
- Graefe, S.; Rodrigo, R.; Cueva, E.; Butz, P.; Werner, F.A.; Homeier, J. Impact of disturbance on forest structure and tree species composition in a tropical dry forest of South Ecuador. Ecotropica 2020, 22, 202002. [Google Scholar]
- Valle, D.; Griffith, D.M.; Jara-Guerrero, A.; Armijos-Ojeda, D.; Espinosa, C.I. A multifaceted approach to understanding bat community response to disturbance in a seasonally dry tropical forest. Sci. Rep. 2021, 11, 5667. [Google Scholar] [CrossRef] [PubMed]
- Benítez, Á.; Aragón, G.; Prieto, M. Lichen diversity on tree trunks in tropical dry forests is highly influenced by host tree traits. Biodivers. Conserv. 2019, 28, 2909–2929. [Google Scholar] [CrossRef]
- Lücking, R.; Moncada, B.; Martínez-Habibe, M.C.; Salgado-Negret, B.E.; Celis, M.; Rojas-Zamora, O.; Borsch, T. Lichen diversity in colombian caribbean dry forest remnants. Caldasia 2019, 41, 194–214. [Google Scholar] [CrossRef]
- Nieder, J.; Prosperí, J.; Michaloud, G. Epiphytes and their contribution to canopy diversity. Plant Ecol. 2001, 153, 51–63. [Google Scholar] [CrossRef]
- Holz, I.; Gradstein, R.S. Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica–species richness, community composition and ecology. Plant Ecol. 2005, 178, 89–109. [Google Scholar] [CrossRef]
- Mandl, N.; Lehnert, M.; Kessler, M.; Gradstein, S.R. A comparison of alpha and beta diversity patterns of ferns, bryophytes and macrolichens in tropical montane forests of southern Ecuador. Biodivers. Conserv. 2010, 19, 2359–2369. [Google Scholar] [CrossRef]
- Werner, F.A.; Gradstein, S.R. Diversity of epiphytes of tropical dry forest along a gradient of human disturbance in the Andes of Ecuador. J. Veg. Sci. 2009, 20, 59–68. [Google Scholar] [CrossRef]
- Nash, T.H. (Ed.) Lichen Biology; Cambridge University Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef] [PubMed]
- Higuera, D.; Wolf, J.H. Vascular epiphytes in dry oak forests show resilience to anthropogenic disturbance, Cordillera Oriental, Colombia. Caldasia 2010, 32, 161–174. [Google Scholar]
- De la Rosa-Manzano, E.; Andrade, J.L.; Zotz, G.; Reyes-García, C. Epiphytic orchids in tropical dry forests of Yucatan, Mexico–Species occurrence, abundance and correlations with host tree characteristics and environmental conditions. Flora Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 100–109. [Google Scholar] [CrossRef]
- Wolseley, P.A.; Aguirre-Hudson, B. Fire in tropical dry forests: Corticolous lichens as indicators of recent ecological changes in Thailand. J. Biogeogr. 1997, 24, 345–362. [Google Scholar] [CrossRef]
- Werner, F.A.; Homeier, J.; Gradstein, S.R. Diversity of vascular epiphytes on isolated remnant trees in the montane forest belt of southern Ecuador. Ecotropica 2005, 11, 21–40. [Google Scholar]
- Gradstein, S.R. Epiphytes of tropical montane forests-impact of deforestation and climate change. Göttingen Centre for Biodiversity and Ecology. Biodivers. Mar. Ecol. Prog. Ser. 2008, 2, 51–65. [Google Scholar]
- McGee, G.G.; Kimmerer, R.W. Forest age and management effects on epiphytic bryophyte communities in Adirondack northern hardwood forests, New York, USA. Can. J. For. Res. 2002, 32, 1562–1576. [Google Scholar] [CrossRef]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef]
- Cáceres, M.E.S. Corticolous crustose and microfoliose lichens of northeastern Brazil. Libri Botanici. 2007, 22, 1–168. [Google Scholar]
- Benítez, Á.; Prieto, M.; Aragón, G. Large trees and dense canopies: Key factors for maintaining high epiphytic diversity on trunk bases (bryophytes and lichens) in tropical montane forests. Int. J. For. Res. 2015, 88, 521–527. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Camarero, J.J.; Espinosa, C.I. Intra-annual stem increment patterns and climatic responses in five tree species from an Ecuadorian tropical dry forest. J. Tree Sci. 2017, 31, 1057–1067. [Google Scholar] [CrossRef]
- Cueva Ortiz, J.; Chalán, L.A. Cobertura Vegetal y Uso Actual del Suelo de la Provincia de Loja: Informe Técnico; Gráficas Amazonas: Cuenca, Ecuador, 2010. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; C.R.C. Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Permanova+ for Primer: Guide to Software and Statistical Methods; The University of Chicago Press: Plymouth, MI, USA, 2008. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Okasanen, M.J. Vegan: Community Ecology Package; R CRAN: Helsinki, Finland, 2018. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Nöske, N.M.; Hilt, N.; Werner, F.A.; Brehm, G.; Fiedler, K.; Sipman, H.J.; Gradstein, S.R. Disturbance effects on diversity of epiphytes and moths in a montane forest in Ecuador. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- García-Martínez, S.; Mercado-Gómez, J.D. Contribution to lichen microbiota of the colombian tropical dry forest (Montes de María, Sucre). Cienc. Desarro. 2020, 11, 43–52. [Google Scholar]
- Cueva Ortiz, J.; Espinosa, C.I.; Quiroz Dahik, C.; Aguirre Mendoza, Z.; Cueva Ortiz, E.; Gusmán, E.; Hildebrandt, P. Influence of anthropogenic factors on the diversity and structure of a dry forest in the Central Part of the Tumbesian Region (Ecuador–Perú). Forests 2019, 10, 31. [Google Scholar] [CrossRef]
- Déleg, J.; Gradstein, S.R.; Aragón, G.; Giordani, P.; Benítez, Á. Cryptogamic epiphytes as indicators of successional changes in megadiverse lowland rain forests of western Amazonia. Ecol. Indic. 2021, 129, 107890. [Google Scholar] [CrossRef]
- Büdel, B.; Scheidegger, C. Thallus morphology and anatomy. In Lichen Biology, 2nd ed.; Nash, T.H., III, Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 40–68. [Google Scholar]
- Aragón, G.; Martínez, I.; Hurtado, P.; Benítez, Á.; Rodríguez, C.; Prieto, M. Using growth forms to predict epiphytic lichen abundance in a wide variety of forest types. Diversity 2019, 11, 51. [Google Scholar] [CrossRef]
- Cocchietto, M.; Skert, N.; Nimis, P.L.; Sava, G. A review on usnic acid, an interesting natural compound. Naturwissenschaften 2002, 89, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Benítez, A.; Aragón, G.; González, Y.; Prieto, M. Functional traits of epiphytic lichens in response to forest disturbance and as predictors of total richness and diversity. Ecol. Indic. 2018, 86, 18–26. [Google Scholar] [CrossRef]
- Gradstein, S.R.; Churchill, S.P.; Salazar-Allen, N. Guide to the Bryophytes of Tropical America; New York Botanical Garden Press: New York, NY, USA, 2001; Volume 86. [Google Scholar]
- Jiménez, M.S.; Suarez, G.M.; Cabral, R.A. New records of mosses from the Dry Chaco forest of Santiago del Estero, Argentina. Bol. Soc. Argent. Bot. 2020, 55, 547–555. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).