
Integrated Transcriptome and Metabolome Analyses Provided Insight into the Variation in Quality of Green Bamboo (Bambusa oldhamii) Shoots at Different Growth Stages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Materials
2.2. Determination of Lignin, Cellulose, Hemicellulose and Total Flavonoid Content
2.3. Metabolite Extraction and Analysis
2.4. RNA Extraction, Library Construction, and Transcriptome Analysis
2.5. Quantitative Real-Time-PCR Validation
2.6. Statistical Analysis
3. Results
3.1. Changes of Lignin, Cellulose, Hemicellulose and Flavonoid Contents of Green Bamboo Shoots at Three Growth Stages
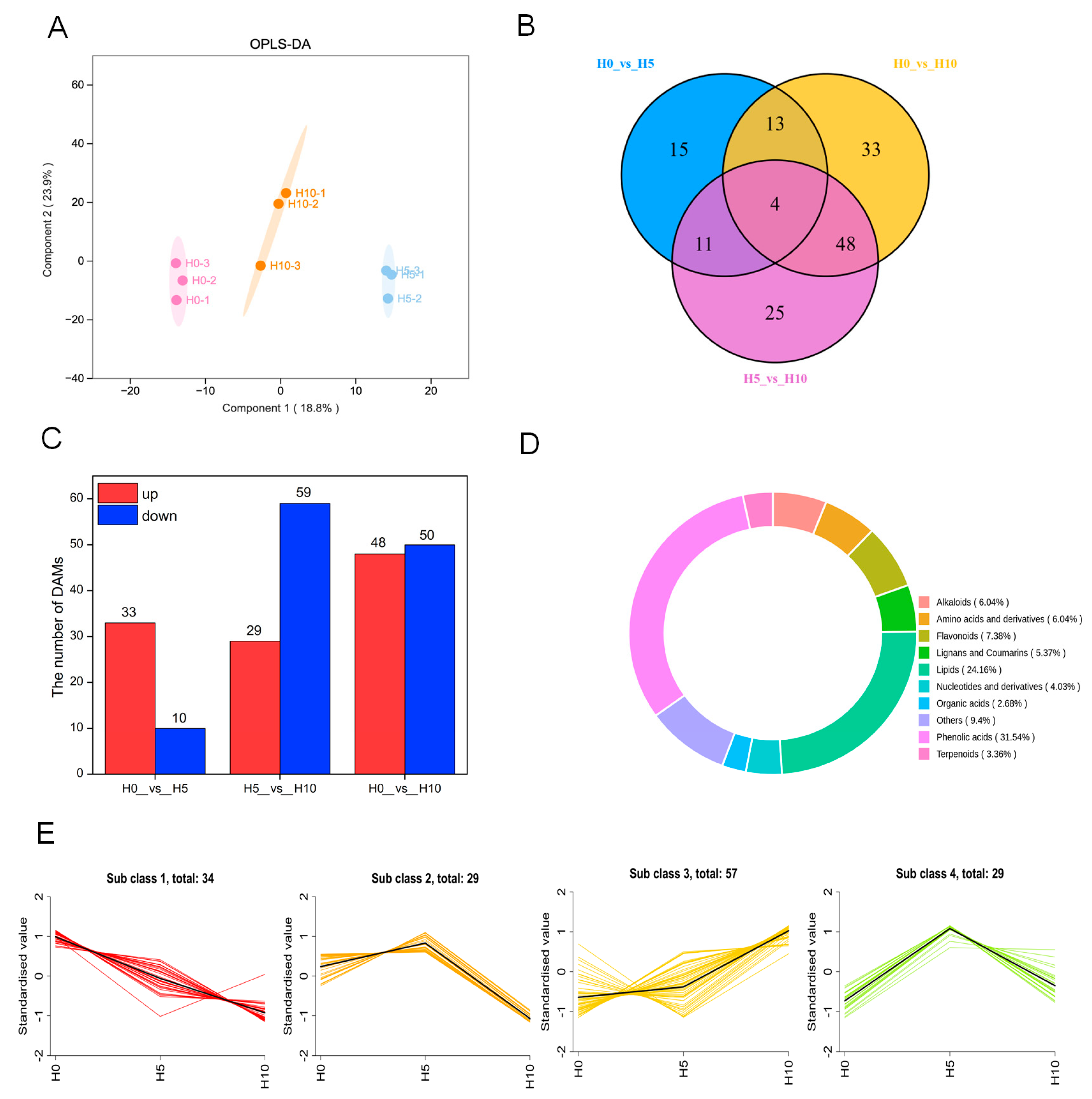
3.2. Metabolomic Changes at Three Growth Stages
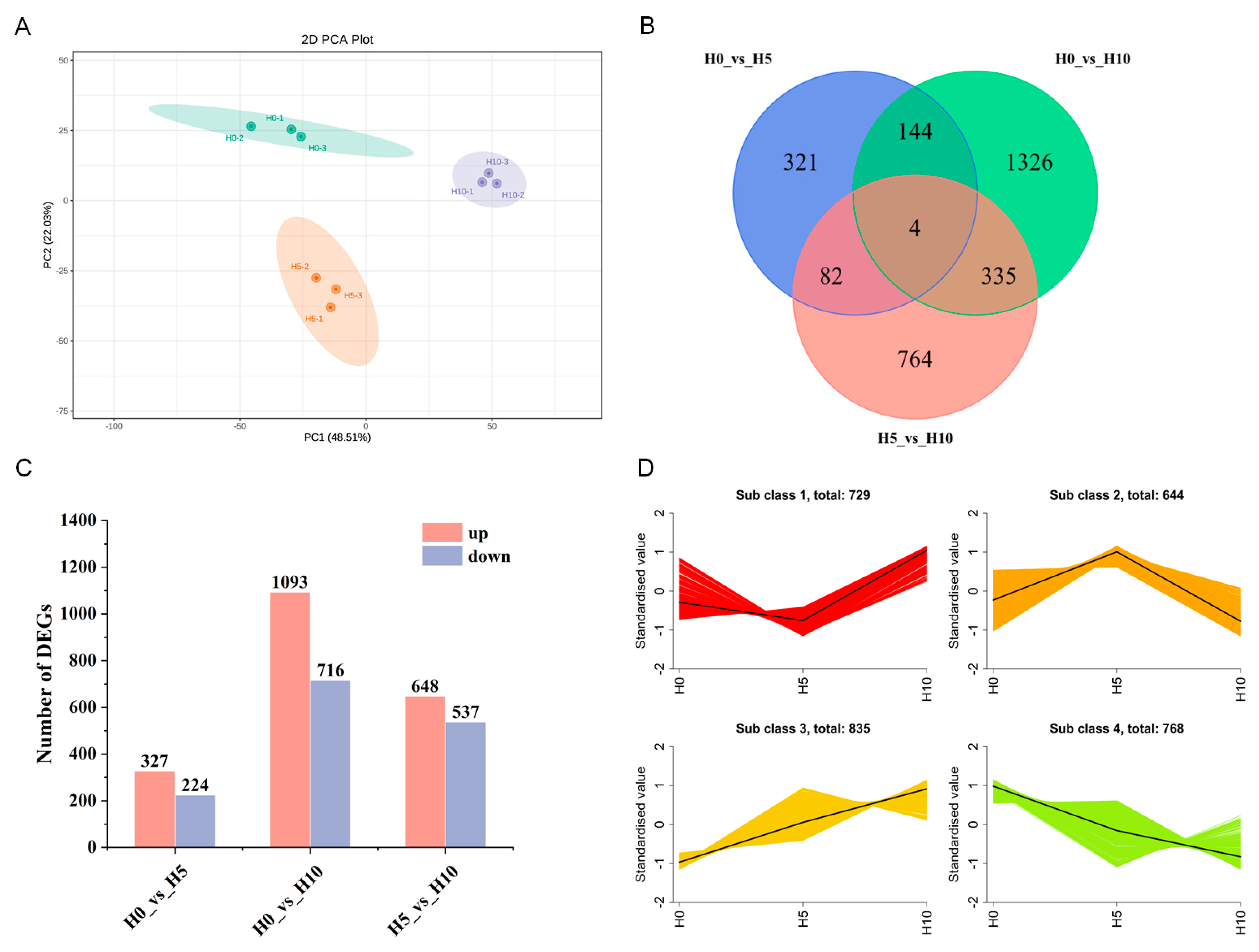
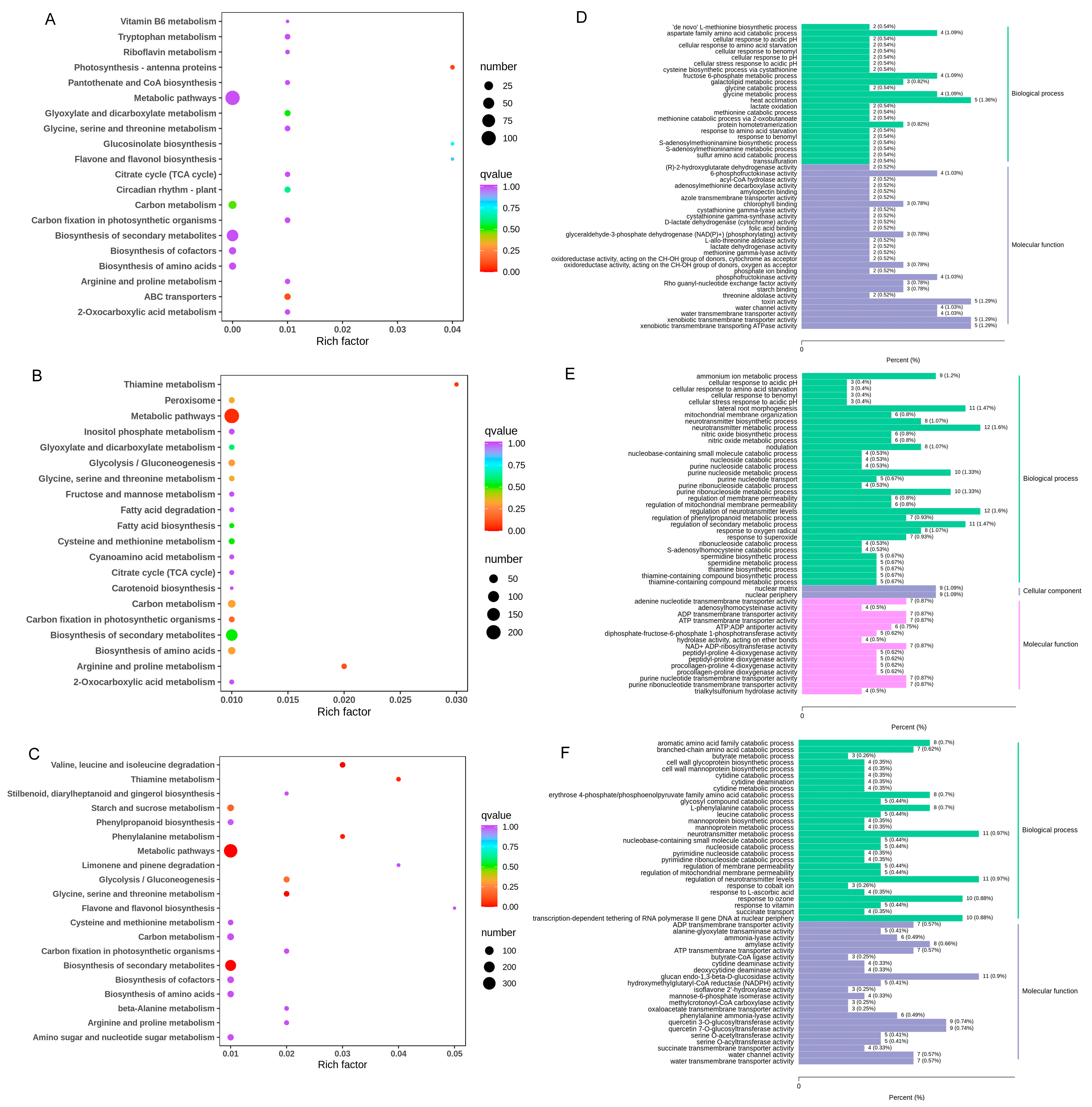
3.3. Transcriptome Analysis and Identification of DEGs of Three Growth Stages
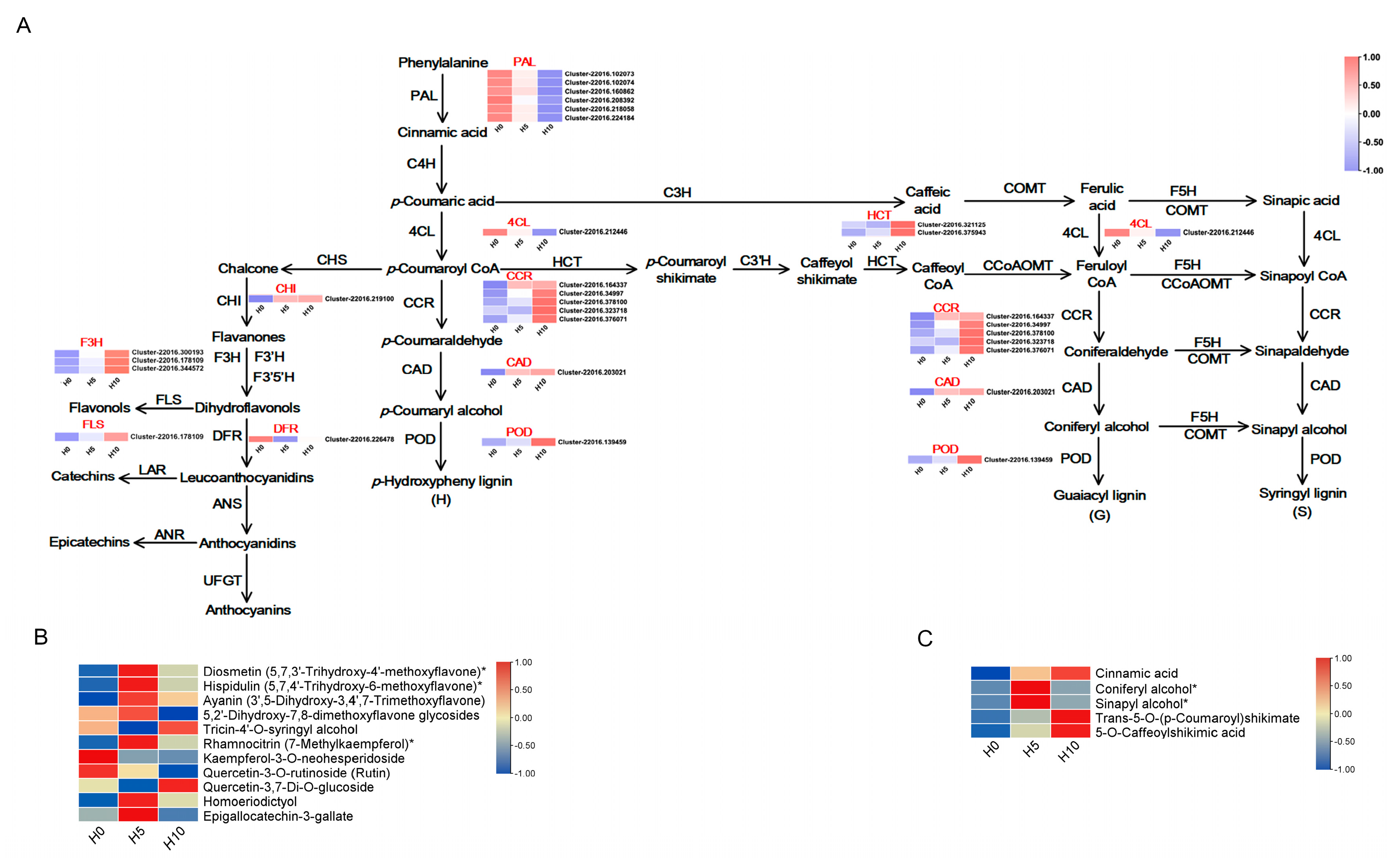
3.4. DEGs and DAMs Related to Lignin and Flavonoid Synthesis
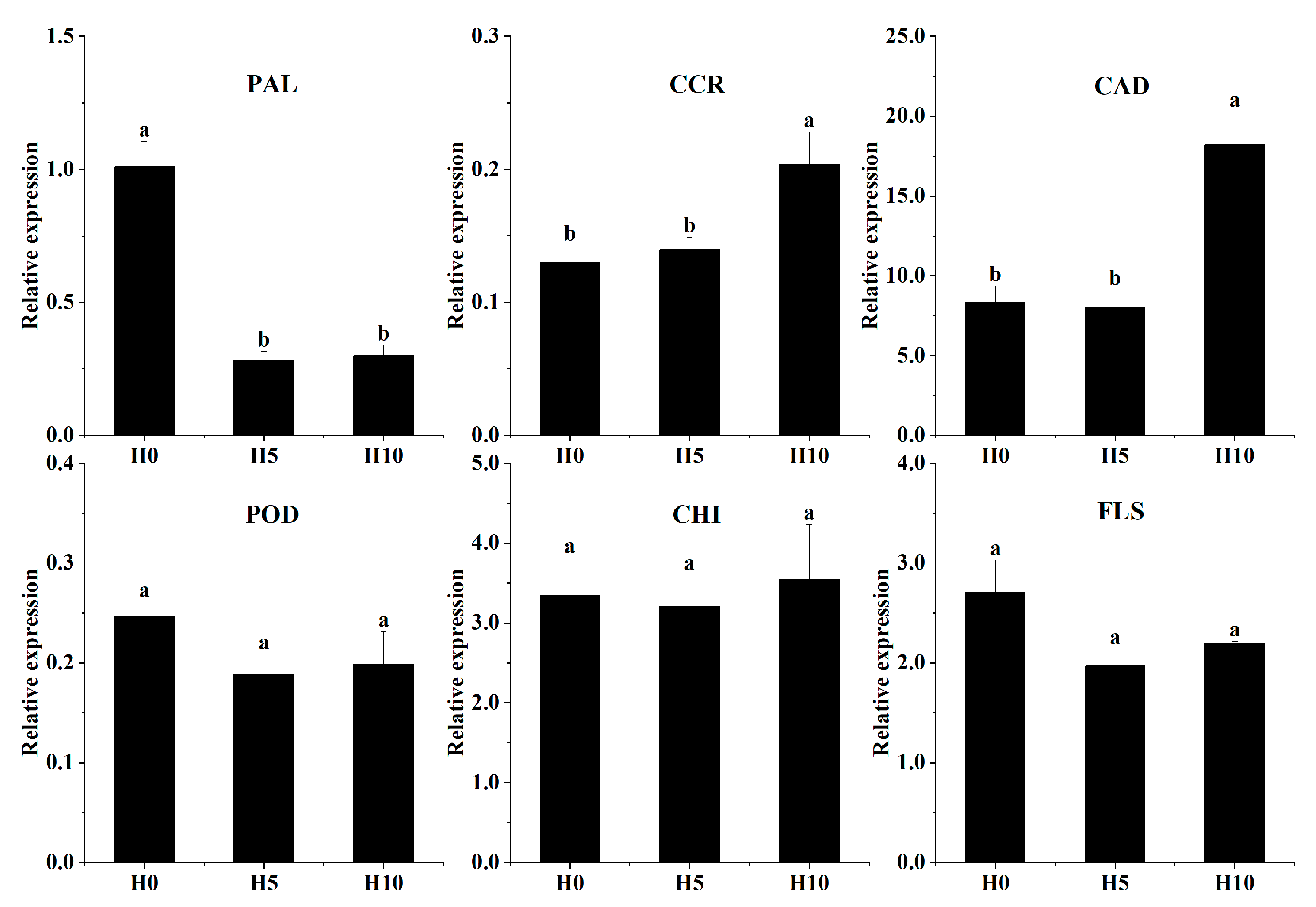
3.5. Validation of Gene Expression in Green Bamboo Shoots via qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S. Bamboo shoots: A novel source of nutrition and medicine. Crit. Rev. Food Sci. Nutr. 2013, 53, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, H.K.; Santosh, O.; Koul, A.; Bisht, M.; Nirmala, C. Quantitative determination of macroelement and microelement content of fresh and processed bamboo shoots by wavelength dispersive X-ray fluorescence spectrometry. X-ray Spectrom. 2019, 48, 637–643. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Wang, D.; Ye, F.; He, Y.; Hu, Z.; Zhao, G. A systematic review on the composition, storage, processing of bamboo shoots: Focusing the nutritional and functional benefits. J. Funct. Foods 2020, 71, 104015. [Google Scholar] [CrossRef]
- Jiao, Y. Transcriptomic and metabolomic analyses reveal the flavor of bitterness in the tip shoots of Bambusa oldhamii Munro. Sci. Rep. 2023, 13, 14853. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Hua, K.; Chen, C.; Yue, J.; Yuan, J. Metabolomic and transcriptomic analysis of bitter compounds in Dendrocalamopsis oldhamii shoots. J. Food Compos. Anal. 2024, 130, 106140. [Google Scholar] [CrossRef]
- Tong, L.; Zhang, L.; Li, B.; Geng, Y.; Wang, L.; Chen, L. Influence of soil-covered cultivation on quality and palatability of Dendrocalamopsis oldhami shoot. Acta Agric. Univ. Jiangxiensis 2018, 40, 487–493. [Google Scholar]
- Liu, K.; Lin, C.; Lin, T.; Tsao, S.; Lo, H. Growth and shoot emergence of green bamboo (Bambusa oldhamii Munro) under different temperatures. In Proceedings of the XXVII International Horticultural Congress-IHC2006: International Symposium on Asian Plants with Unique Horticultural 769, Seoul, Republic of Korea, 13 August 2006; pp. 83–90. [Google Scholar]
- Shi, J.; Zhang, C.; Chen, S.; Gu, R.; Guo, Z.; Ye, H.; Sun, P.; Jiang, Z. Temporal variation of appearance, nutrition and eating quality of Phyllostachys prominens shoots after unearthed. For. Res. 2019, 32, 137–143. [Google Scholar] [CrossRef]
- Wang, X.; Yang, L.; Geng, X.; Shi, W.; Chen, Y.; Lu, C. Integrative analysis of metabolome and transcriptome reveals the different metabolite biosynthesis profiles related to palatability in winter and spring shoot in moso bamboo. Plant Physiol. Biochem. 2023, 202, 107973. [Google Scholar] [CrossRef]
- Bal, L.M.; Singhal, P.; Satya, S.; Naik, S.; Kar, A. Bamboo shoot preservation for enhancing its business potential and local economy: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 804–814. [Google Scholar] [CrossRef]
- Luo, Z.; Feng, S.; Pang, J.; Mao, L.; Shou, H.; Xie, J. Effect of heat treatment on lignification of postharvest bamboo shoots (Phyllostachys praecox f. prevernalis.). Food Chem. 2012, 135, 2182–2187. [Google Scholar] [CrossRef]
- Li, C.; Suo, J.; Xuan, L.; Ding, M.; Zhang, H.; Song, L.; Ying, Y. Bamboo shoot-lignification delay by melatonin during low temperature storage. Postharvest Biol. Technol. 2019, 156, 110933. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, C.; Zhang, H.; Ying, Y.; Hu, Y.; Song, L. Comparative analysis of the lignification process of two bamboo shoots stored at room temperature. Plants 2020, 9, 1399. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Lu, H.; Zhao, Z.; Pei, J.; Yang, H.; Wu, A.; Yu, X.; Lin, X. Integrative transcriptomic and metabolomic data provide insights into gene networks associated with lignification in postharvest Lei bamboo shoots under low temperature. Food Chem. 2022, 368, 130822. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, X.; Yang, K.; Zhu, C.; Liu, Y.; Gao, Z. Multifaceted analyses reveal carbohydrate metabolism mainly affecting the quality of postharvest bamboo shoots. Front. Plant Sci. 2022, 13, 1021161. [Google Scholar] [CrossRef] [PubMed]
- Mack, C.; Wefers, D.; Schuster, P.; Weinert, C.H.; Egert, B.; Bliedung, S.; Trierweiler, B.; Muhle-Goll, C.; Bunzel, M.; Luy, B. Untargeted multi-platform analysis of the metabolome and the non-starch polysaccharides of kiwifruit during postharvest ripening. Postharvest Biol. Technol. 2017, 125, 65–76. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhang, K.; Chen, W.; Li, X.; Zhu, X. Transcriptomic and metabolomic analyses reveal key factors regulating chilling stress-induced softening disorder in papaya fruit. Postharvest Biol. Technol. 2023, 205, 112534. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Zhang, J.; Ge, H.; Li, S.; Li, X.; Yin, X.; Grierson, D.; Chen, K. EjMYB8 transcriptionally regulates flesh lignification in loquat fruit. PLoS ONE 2016, 11, e0154399. [Google Scholar] [CrossRef]
- Zeng, J.K.; Li, X.; Zhang, J.; Ge, H.; Yin, X.R.; Chen, K.S. Regulation of loquat fruit low temperature response and lignification involves interaction of heat shock factors and genes associated with lignin biosynthesis. Plant Cell Environ. 2016, 39, 1780–1789. [Google Scholar] [CrossRef]
- Huang, W.; Zhu, N.; Zhu, C.; Wu, D.; Chen, K. Morphology and cell wall composition changes in lignified cells from loquat fruit during postharvest storage. Postharvest Biol. Technol. 2019, 157, 110975. [Google Scholar] [CrossRef]
- Zhu, N.; Zhao, C.; Wei, Y.; Sun, C.; Wu, D.; Chen, K. Biosynthetic labeling with 3-O-propargylcaffeyl alcohol reveals in vivo cell-specific patterned lignification in loquat fruits during development and postharvest storage. Hortic. Res. 2021, 8, 61. [Google Scholar] [CrossRef]
- Li, C.; Xuan, L.; He, Y.; Wang, J.; Zhang, H.; Ying, Y.; Wu, A.; Bacic, A.; Zeng, W.; Song, L. Molecular Mechanism of Xylogenesis in Moso Bamboo (Phyllostachys edulis) Shoots during Cold Storage. Polymers 2018, 11, 38. [Google Scholar] [CrossRef]
- Boonsiriwit, A.; Lee, M.; Kim, M.; Itkor, P.; Lee, Y.S. Exogenous melatonin reduces lignification and retains quality of green asparagus (Asparagus officinalis L.). Foods 2021, 10, 2111. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, J.; Wagner, S.; Trierweiler, B.; Bunzel, M. Characterization of cell wall components and their modifications during postharvest storage of Asparagus officinalis L.: Storage-related changes in dietary fiber composition. J. Agric. Food Chem. 2016, 64, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Campbell, L.; Turner, S. Secondary cell walls: Biosynthesis and manipulation. J. Exp. Bot. 2016, 67, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Lu, S. Biosynthesis and regulation of phenylpropanoids in plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Gao, Q.; Jiang, H.; Tang, F.; Cao, H.; Wu, X.; Qi, F.; Sun, J.; Yang, J. Evaluation of the bitter components of bamboo shoots using a metabolomics approach. Food Funct. 2019, 10, 90–98. [Google Scholar] [CrossRef]
- Zhang, Z.; Ding, X.; Cui, F.; Cai, H. Impact of avoiding light on bitterness and astringency, tannin content, morphology and distribution of Dendrocalamus latiflorus. For. Res. 2016, 29, 770–777. [Google Scholar]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef]
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health. Biotechnol. J. Healthc. Nutr. Technol. 2007, 2, 1214–1234. [Google Scholar] [CrossRef]
- Chen, M.; Guo, L.; Ramakrishnan, M.; Fei, Z.; Vinod, K.K.; Ding, Y.; Jiao, C.; Gao, Z.; Zha, R.; Wang, C.; et al. Rapid growth of Moso bamboo (Phyllostachys edulis): Cellular roadmaps, transcriptome dynamics, and environmental factors. Plant Cell 2022, 34, 3577–3610. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Z.; Liao, Z.; Liu, X.; Li, Y.; Zhou, C.; Sun, C.; Wang, Y.; Cao, J.; Sun, C. Integrated metabolomic–transcriptomic analyses of flavonoid accumulation in citrus fruit under exogenous melatonin treatment. Int. J. Mol. Sci. 2024, 25, 6632. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Walker, A.R. Biosynthesis and regulation of flavonoids in buckwheat. Breed. Sci. 2020, 70, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Qiu, X.; Pan, X.; Li, C. Transcriptional factor-mediated regulation of active component biosynthesis in medicinal plants. Curr. Pharm. Biotechnol. 2021, 22, 848–866. [Google Scholar] [CrossRef]
- Yan, Q.; Zhang, G.; Zhang, X.; Huang, L. A review of transcriptomics and metabolomics in plant quality and environmental response: From bibliometric analysis to science mapping and future trends. Metabolites 2024, 14, 272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, J.; Li, H.; Chiang, V.L.; Fu, Y. Cooperative regulation of flavonoid and lignin biosynthesis in plants. Crit. Rev. Plant Sci. 2021, 40, 109–126. [Google Scholar] [CrossRef]
- Wang, J. The Study of Expression and Analysis of Different Expression Genes in Moso Bamboo Shoot Transcriptome Associated with the Lignification. Master’s Thesis, Nanjing Agriculture University, Nanjing, China, 2016. [Google Scholar]
- Miao, N.; Yun, C.; Han, S.; Shi, Y.; Gao, Y.; Wu, S.; Zhao, Z.; Wang, H.; Wang, W. Postharvest UV-A radiation affects flavonoid content, composition, and bioactivity of Scutellaria baicalensis root. Postharvest Biol. Technol. 2022, 189, 111933. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Nongdam, P.; Tikendra, L. The nutritional facts of bamboo shoots and their usage as important traditional foods of northeast India. Int. Sch. Res. Not. 2014, 2014, 679073. [Google Scholar] [CrossRef]
- Guo, M.; He, Y.; Pan, K.; Bao, F.; Ying, Y. Analysis of differential metabolites of Phyllostachys edulis shoots at different growth stages by ultra-high performance liquid chromatography-tandem mass spectrometry. Food Sci. 2023, 44, 283–291. [Google Scholar]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food sources, bioavailability, metabolism, and bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Sahu, J.K.; Sharma, G. Biochemistry of bitterness in bamboo shoots. Assam Univ. J. Sci. Technol. 2010, 6, 105–111. [Google Scholar]
- Yang, J.; Li, B.; Shi, W.; Gong, Z.; Chen, L.; Hou, Z. Transcriptional activation of anthocyanin biosynthesis in developing fruit of blueberries (Vaccinium corymbosum L.) by preharvest and postharvest UV irradiation. J. Agric. Food Chem. 2018, 66, 10931–10942. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shi, W.; Li, B.; Bai, Y.; Hou, Z. Preharvest and postharvest UV radiation affected flavonoid metabolism and antioxidant capacity differently in developing blueberries (Vaccinium corymbosum L.). Food Chem. 2019, 301, 125248. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Postharvest UV-C irradiation increased the flavonoids and anthocyanins accumulation, phenylpropanoid pathway gene expression, and antioxidant activity in sweet cherries (Prunus avium L.). Postharvest Biol. Technol. 2021, 175, 111490. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Cao, X.; Pan, K.; Shi, W.; Ying, Y. Integrated Transcriptome and Metabolome Analyses Provided Insight into the Variation in Quality of Green Bamboo (Bambusa oldhamii) Shoots at Different Growth Stages. Forests 2024, 15, 1582. https://doi.org/10.3390/f15091582
He Y, Cao X, Pan K, Shi W, Ying Y. Integrated Transcriptome and Metabolome Analyses Provided Insight into the Variation in Quality of Green Bamboo (Bambusa oldhamii) Shoots at Different Growth Stages. Forests. 2024; 15(9):1582. https://doi.org/10.3390/f15091582
Chicago/Turabian StyleHe, Yuelin, Xiangmei Cao, Kaiting Pan, Wenhui Shi, and Yeqing Ying. 2024. "Integrated Transcriptome and Metabolome Analyses Provided Insight into the Variation in Quality of Green Bamboo (Bambusa oldhamii) Shoots at Different Growth Stages" Forests 15, no. 9: 1582. https://doi.org/10.3390/f15091582
APA StyleHe, Y., Cao, X., Pan, K., Shi, W., & Ying, Y. (2024). Integrated Transcriptome and Metabolome Analyses Provided Insight into the Variation in Quality of Green Bamboo (Bambusa oldhamii) Shoots at Different Growth Stages. Forests, 15(9), 1582. https://doi.org/10.3390/f15091582