
Successions of Bacterial and Fungal Communities in Biological Soil Crust under Sand-Fixation Plantation in Horqin Sandy Land, Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Site Description
2.2. Soil Sampling
2.3. Determination of Soil Properties
2.4. DNA Extraction, 16S rDNA and Internal Transcribed Spacer (ITS) rDNA Sequencing, and Data Processing and Analysis
2.5. Regression Analysis and Redundancy Analysis (RDA)
3. Results
3.1. Soil Property Improvement along BSC Development
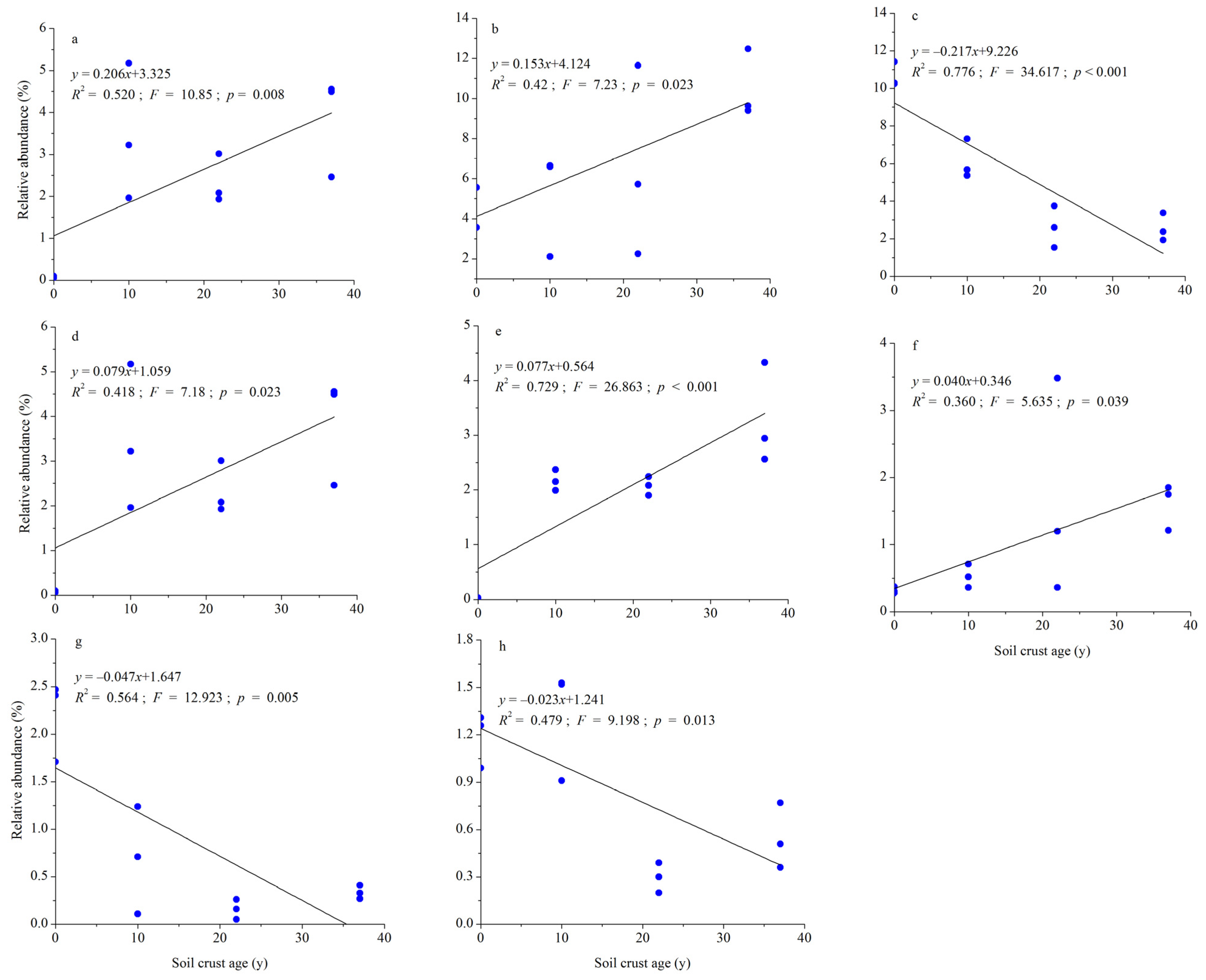
3.2. Response of Soil Bacterial Community to BSC Development
3.3. Response of Soil Fungal Community to BSC Development
3.4. Dependence of Bacterial and Fungal Community Structures on Soil Variables
4. Discussion
4.1. The Formation of BSC and Soil Property Improvement
4.2. Recovery and Succession of Soil Bacterial/Fungal Community in BSC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, L.N.; Liu, Y.B.; Wang, Z.R.; Yuan, S.W.; Qi, J.H.; Zhang, W.L.; Wang, Y.S.; Li, X.R. Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems. Plant Soil 2020, 447, 379–392. [Google Scholar] [CrossRef]
- Soule, T.; Anderson, I.J.; Johnson, S.L.; Bates, S.T.; Garcia-Pichel, F. Archaeal populations in biological soil crusts from arid lands in North America. Soil Biol. Biochem. 2009, 41, 2069–2074. [Google Scholar] [CrossRef]
- Weber, B.; Wu, D.M.; Tamm, A.; Ruckteschler, N.; Rodriguez-Caballero, E.; Steinkamp, J.; Meusel, H.; Elbert, W.; Behrendt, T.; Sorgel, M.; et al. Biological soil crusts accelerate the nitrogen cycle through large NO and HONO emissions in drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15384–15389. [Google Scholar] [CrossRef] [PubMed]
- Lan, S.B.; Wu, L.; Zhang, D.L.; Hu, C.X. Successional stages of biological soil crusts and their microstructure variability in Shapotou region (China). Environ. Earth Sci. 2012, 65, 77–88. [Google Scholar] [CrossRef]
- Zhao, X.X.; Zhang, Y.; Cui, Z.B.; Peng, L.; Cao, C.Y. Dynamics of phoD- and gcd-harboring microbial communities across an age sequence of biological soil crusts under sand-fixation plantation. Front. Microbiol. 2022, 13, 831888. [Google Scholar] [CrossRef] [PubMed]
- Büdel, B.; Williams, W.J.; Reichenberger, H. Annual net primary productivity of a cyanobacteria-dominated biological soil crust in the Gulf Savannah, Queensland, Australia. Biogeosciences 2018, 15, 491–505. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, C.Y.; Peng, M.; Xu, X.J.; Zhang, P.; Yu, Q.J.; Sun, T. Diversity of nitrogen-fixing, ammonia-oxidizing, and denitrifying bacteria in biological soil crusts of a revegetation area in Horqin Sandy Land, Northeast China. Ecol. Eng. 2014, 71, 71–79. [Google Scholar] [CrossRef]
- Belnap, J. Nitrogen fixation in biological soil crusts from Southeast Utah, USA. Biol. Fertil. Soils 2002, 35, 128–135. [Google Scholar] [CrossRef]
- Evans, R.D.; Johansen, J.R. Microbiotic crusts and ecosystem processes. Crit. Rev. Plant Sci. 1999, 18, 183–225. [Google Scholar] [CrossRef]
- Zhao, H.L.; Guo, Y.R.; Zhou, R.L.; Drake, S. Biological soil crust and surface soil properties in different vegetation types of Horqin sand land, China. Catena 2010, 82, 70–76. [Google Scholar] [CrossRef]
- Belnap, J.; Eldridge, D. Disturbance and recovery of biological soil crusts. In Biological Soil Crusts, Structure, Function, and Management; Belnap, J., Lange, O.L., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 363–383. [Google Scholar]
- Hawkes, C.V. Nitrogen cycling mediated by biological soil crusts and arbuscular mycorrhizal fungi. Ecology 2003, 84, 1553–1562. [Google Scholar] [CrossRef]
- Kidron, G.J.; Temina, M. The mycobiont role in crustose lichen expansion on cobbles in the Negev Desert. Geomicrobiol. J. 2008, 25, 95–100. [Google Scholar] [CrossRef]
- Smith, S.M.; Abed, R.M.M.; Garcia-Pichel, F. Biological soil crusts of sand dunes in Cape Cod National Seashore, Massachusetts, USA. Microb. Ecol. 2004, 48, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Zhou, R.L.; Su, Y.Z.; Zhang, H.; Zhao, L.Y.; Drake, S. Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia. Ecol. Eng. 2007, 31, 1–8. [Google Scholar] [CrossRef]
- Morillas, L.; Gallardo, A. Biological soil crusts and wetting events, effects on soil N and C cycles. Appl. Soil Ecol. 2015, 94, 1–6. [Google Scholar] [CrossRef]
- FAO. FAO/IUSS Working Group WRB, World reference base for soil resources. In World Soil Resources Reports 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Institute of Soil Science, Chinese Academy of Sciences (ISSCAS). Physical and Chemical Analysis Methods of Soils; Shanghai Science Technology Press: Shanghai, China, 1978; pp. 7–59. [Google Scholar]
- Ladd, J.N.; Butler, J.H.A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Perucci, P.; Casucc, C.; Dumontet, S. An improved method to evaluate the o-diphenol oxidase activity of soil. Soil Biol. Biochem. 2000, 32, 1927–1933. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis; Page, A.L., Millar, E.M., Keeney, D.R., Eds.; ASA and SSSA: Madison, WI, USA, 1982; pp. 501–538. [Google Scholar]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology, Springer Lab Manual; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Institute of Soil Science, Chinese Academy of Sciences (ISSCAS). Methods on Soil Microorganism Study; Science Press: Beijing, China, 1985; pp. 260–275. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Mayland, H.F.; McIntosh, T.H. Availability of biologically fixed atmospheric nitrogen-15 to higher plants. Nature 1966, 209, 421–422. [Google Scholar] [CrossRef]
- Song, G.; Li, X.R.; Hui, R. Biological soil crusts increase stability and invasion resistance of desert revegetation communities in northern China. Ecosphere 2020, 11, e03043. [Google Scholar] [CrossRef]
- Su, Y.Z.; Zhang, T.H.; Li, Y.L.; Wang, F. Changes in soil properties after establishment of Artemisia halodendron and Caragana microphylla on shifting sand dunes in semi-arid Horqin Sandy Land, Northern China. Environ. Manag. 2005, 36, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Li, S.G.; Harazono, Y.; Zhao, H.L.; He, Z.Y.; Chang, X.L.; Zhao, X.Y.; Zhang, T.H.; Oikawa, T. Micrometeorological changes following establishment of artificially established Artemisia vegetation on desertified sandy land in the Horqin sandy land, China and their implication on regional environmental change. J. Arid. Environ. 2002, 52, 101–119. [Google Scholar] [CrossRef]
- Liu, X.M.; Zhao, H.L. Comprehensive Strategy for Eco-Environmental Control in Horqin Sand Land; Gansu Science and Technology Publishing House: Lanzhou, China, 1993. [Google Scholar]
- Wang, F.; Zhang, Y.; Xia, Y.; Cui, Z.B.; Cao, C.Y. Soil microbial community succession based on phoD and gcd genes along a chronosequence of sand-fixation forest. Forests 2021, 12, 1707. [Google Scholar] [CrossRef]
- Liu, Y.M.; Yang, H.Y.; Li, X.; Xing, Z.S. Effects of biological soil crusts on soil enzyme activities in revegetated areas of the Tengger Desert. Appl. Soil Ecol. 2014, 80, 6–14. [Google Scholar] [CrossRef]
- Badiane, N.N.Y.; Chotte, J.L.; Pate, E.; Masse, D.; Rouland, C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol. 2001, 18, 229–238. [Google Scholar] [CrossRef]
- Ullah, S.; Ai, C.; Ding, W.C.; Jiang, R.; Zhao, S.C.; Zhang, J.J.; Zhou, W.; Hou, Y.P.; He, P. The response of soil fungal diversity and community composition to long-term fertilization. Appl. Soil Ecol. 2019, 140, 35–41. [Google Scholar] [CrossRef]
- Urbanova, M.; Snajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Miki, T.; Ushio, M.; Fukui, S.; Kondoh, M. Functional diversity of microbial decomposers facilitates plant coexistence in a plant microbe-soil feedback model. Proc. Natl. Acad. Sci. USA 2010, 107, 14251–14256. [Google Scholar] [CrossRef]
- Yang, C.Y.; Li, Y.; Zhou, B.; Zhou, Y.Y.; Zheng, W.; Tian, Y.; Van Nostrand, J.D.; Wu, L.Y.; He, Z.L.; Zhou, J.Z.; et al. Illumina sequencing based analysis of free-living bacterial community dynamics during an Akashiwo sanguine bloom in Xiamen Sea, China. Sci. Rep. 2015, 5, 8476. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Xia, B.C.; Huang, H.; Palumbo, A.V.; Tiedje, J.M. Microbial diversity and heterogeneity in sandy subsurface soils. Appl. Microbiol. Biotechnol. 2004, 70, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Drees, K.P.; Neilson, J.W.; Betancourt, J.L.; Quade, J.; Henderson, D.A.; Pryor, B.M.; Maier, R.M. Bacterial community structure in the hyperarid core of the Atacama Desert, Chile. Appl. Environ. Microbiol. 2006, 72, 7902–7908. [Google Scholar] [CrossRef]
- Delgado, E.F.; Valdez, A.T.; Covarrubias, S.A.; Tosi, S.; Nicola, L. Soil fungal diversity of the Aguarongo Andean Forest (Ecuador). Biology 2022, 10, 1289. [Google Scholar] [CrossRef] [PubMed]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Schmidt, T.M.; Coleman, D.C.; Whitman, W.B. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol. Biochem. 2011, 43, 2184–2193. [Google Scholar] [CrossRef]
- Suleiman, A.K.A.; Manoeli, L.; Boldo, J.T.; Pereira, M.G.; Roesch, L.F.W. Shifts in soil bacterial community after eight years of land-use change. Syst. Appl. Microbiol. 2013, 36, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kara, O.; Bolat, I.; Cakiroglu, K.; Ozturk, M. Plant canopy effects on litter accumulation and soil microbial biomass in two temperate forests. Biol. Fertil. Soils 2008, 45, 193–198. [Google Scholar] [CrossRef]
- Fierer, N.; Nemergut, D.; Knight, R.; Craine, J.M. Changes through time, integrating microorganisms into the study of succession. Res. Microbiol. 2010, 161, 635–642. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | MSD | SC10 | SC22 | SC37 | ANOVA in Response to Age | |||
|---|---|---|---|---|---|---|---|---|
| Regress Equation | R2 | F | p | |||||
| Soil moisture (%) | 0.25 ± 0.09 | 0.69 ± 0.15 | 0.87 ± 0.08 | 1.09 ± 0.146 | y = 0.021x+ 0.358 | 0.835 | 50.55 | <0.001 |
| pH | 6.64 ± 0.13 | 6.81 ± 0.06 | 6.93 ± 0.08 | 7.03 ± 0.05 | y = 0.010x + 6.676 | 0.783 | 36.02 | <0.001 |
| Soil organic matter (%) | 0.09 ± 0.01 | 0.43 ± 0.11 | 0.83 ± 0.10 | 1.10 ± 0.15 | y = 0.027x + 0.139 | 0.932 | 136.1 | <0.001 |
| Total N (%) | 0.018 ± 0.001 | 0.019 ± 0.002 | 0.022 ± 0.002 | 0.026 ± 0.003 | y = 0.001x + 0.007 | 0.731 | 27.21 | <0.001 |
| Total P (%) | 0.041 ± 0.001 | 0.022 ± 0.001 | 0.024 ± 0.004 | 0.029 ± 0.006 | y = 0.001x + 0.016 | 0.733 | 27.51 | <0.001 |
| Total K (%) | 2.24 ± 0.10 | 2.25 ± 0.12 | 2.23 ± 0.11 | 2.33 ± 0.11 | - | 0.091 | 1.006 | 0.340 |
| NH4-N (mg kg−1) | 1.50 ± 0.07 | 13.5 ± 0.73 | 18.9 ± 1.52 | 15.2 ± 1.08 | y = 0.348x + 6.272 | 0.535 | 11.51 | 0.007 |
| Available P (mg kg−1) | 0.55 ± 0.15 | 4.66 ± 0.79 | 6.82 ± 0.45 | 7.77 ± 0.93 | y = 0.187x + 1.723 | 0.832 | 49.36 | <0.001 |
| Available K (mg kg−1) | 43.8 ± 3.18 | 161.7 ± 4.68 | 178.3 ± 10.5 | 220.4 ± 13.1 | y = 4.288x + 77.07 | 0.808 | 41.96 | <0.001 |
| Nitrate reductase (μg g−1) | 0.006 ± 0.002 | 1.69 ± 0.14 | 1.64 ± 0.37 | 1.75 ± 0.10 | y = 0.042x + 0.468 | 0.657 | 19.14 | 0.001 |
| ALP (mg g−1 h−1) | 5.70 ± 2.75 | 147.4 ± 35.9 | 241.9 ± 85.8 | 408.0 ± 21.6 | y = 10.54x + 18.91 | 0.925 | 123.4 | <0.001 |
| POA (μmol g−1 10 min−1) | 0.34 ± 0.04 | 0.72 ± 0.18 | 0.80 ± 0.18 | 1.02 ± 0.15 | y = 0.017x + 0.430 | 0.724 | 26.22 | <0.001 |
| Dehydrogenase (mg kg−1) | 48.1 ± 1.58 | 183.3 ± 57.5 | 332.9 ± 63.4 | 572.4 ± 97.0 | y = 14.065x + 41.55 | 0.928 | 129.7 | <0.001 |
| Protease (μg g−1 2 h−1) | 4.89 ± 3.72 | 89.8 ± 51.7 | 133.5 ± 8.1 | 196.0 ± 26.7 | y = 4.942x + 20.793 | 0.862 | 62.34 | <0.001 |
| Urease (mg g−1 24 h−1) | 0.11 ± 0.01 | 1.52 ± 0.47 | 2.5 ± 0.42 | 3.42 ± 0.31 | y = 0.087x + 0.381 | 0.914 | 106.2 | <0.001 |
| Index | MSD | SC10 | SC22 | SC37 | R2 | p | |
|---|---|---|---|---|---|---|---|
| Observed OTUs | Bacteria | 398 ± 34 a | 650 ± 68 b | 691 ± 40 b | 736 ± 17 b | 0.691 | 0.001 |
| Fungi | 49 ± 26 a | 329 ± 18 b | 346 ± 56 b | 372 ± 14 b | 0.613 | 0.003 | |
| ACE | Bacteria | 466.6 ± 81.6 a | 720.7 ± 62.7 b | 761.1 ± 792.3 b | 792.3 ± 14.8 b | 0.635 | 0.002 |
| Fungi | 55.1 ± 27.2 a | 346.3 ± 14.0 b | 371.9 ± 46.8 bc | 415.0 ± 18.3 c | 0.669 | 0.001 | |
| Chao1 estimator | Bacteria | 469.2 ± 85.7 a | 729.5 ± 62.6 b | 771.5 ± 20.0 b | 805.3 ± 14.7 b | 0.643 | 0.002 |
| Fungi | 51.5 ± 26.1 a | 349.5 ± 11.7 b | 370.1 ± 45.0 bc | 416.9 ± 20.0 c | 0.666 | 0.010 | |
| Shannon–Wiener | Bacteria | 6.903 ± 0.336 a | 7.239 ± 0.533 b | 7.428 ± 0.163 b | 7.975 ± 0.059 b | 0.665 | 0.001 |
| Fungi | 3.828 ± 0.478 a | 5.343 ± 0.636 a | 4.677 ± 1.030 a | 5.121 ± 0.262 a | 0.180 | 0.170 | |
| Index | SM | pH | SOM | TN | TP | TK | NH4-N | AP | AK | |
|---|---|---|---|---|---|---|---|---|---|---|
| Observed OTUs | Bacteria | 0.907 ** | 0.857 ** | 0.851 ** | 0.941 ** | 0.794 ** | 0.274 | 0.909 ** | 0.950 ** | 0.951 ** |
| Fungi | 0.878 ** | 0.810 ** | 0.806 ** | 0.953 ** | 0.784 ** | 0.173 | 0.934 ** | 0.924 * | 0.948 ** | |
| ACE | Bacteria | 0.892 ** | 0.858 * | 0.821 ** | 0.929 ** | 0.772 ** | 0.278 | 0.901 ** | 0.925 ** | 0.924 ** |
| Fungi | 0.900 ** | 0.833 ** | 0.834 ** | 0.968 ** | 0.806 ** | 0.181 | 0.934 ** | 0.939 ** | 0.963 ** | |
| Chao1 estimator | Bacteria | 0.896 ** | 0.857 * | 0.826 ** | 0.933 ** | 0.778 ** | 0.264 | 0.900 ** | 0.926 ** | 0.926 ** |
| Fungi | 0.903 ** | 0.828 ** | 0.830 ** | 0.968 ** | 0.797 ** | 0.195 | 0.931 ** | 0.934 ** | 0.963 ** | |
| Shannon–Wiener | Bacteria | 0.823 ** | 0.626 * | 0.789 ** | 0.662 * | 0.646 * | 0.112 | 0.549 | 0.730 ** | 0.739 ** |
| Fungi | 0.546 | 0.499 | 0.440 | 0.581 * | 0.409 | 0.428 | 0.520 | 0.557 | 0.611 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, C.; Zhang, Y.; Cui, Z. Successions of Bacterial and Fungal Communities in Biological Soil Crust under Sand-Fixation Plantation in Horqin Sandy Land, Northeast China. Forests 2024, 15, 1631. https://doi.org/10.3390/f15091631
Cao C, Zhang Y, Cui Z. Successions of Bacterial and Fungal Communities in Biological Soil Crust under Sand-Fixation Plantation in Horqin Sandy Land, Northeast China. Forests. 2024; 15(9):1631. https://doi.org/10.3390/f15091631
Chicago/Turabian StyleCao, Chengyou, Ying Zhang, and Zhenbo Cui. 2024. "Successions of Bacterial and Fungal Communities in Biological Soil Crust under Sand-Fixation Plantation in Horqin Sandy Land, Northeast China" Forests 15, no. 9: 1631. https://doi.org/10.3390/f15091631