
Effects of Fertilization and Drip Irrigation on the Growth of Populus × canadensis ‘Zhongliao 1’ Plantation and on Soil Physicochemical Properties and Enzyme Activities

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Experiment Design
2.3. Measurement of Growth Traits
2.4. Soil Sample Collection
2.5. Determination of Soil Physicochemical Properties and Enzyme Activities
2.6. Data Analysis
3. Results
3.1. Tree Height and DBH Growth Characteristics of P. × canadensis ‘Zhongliao 1’
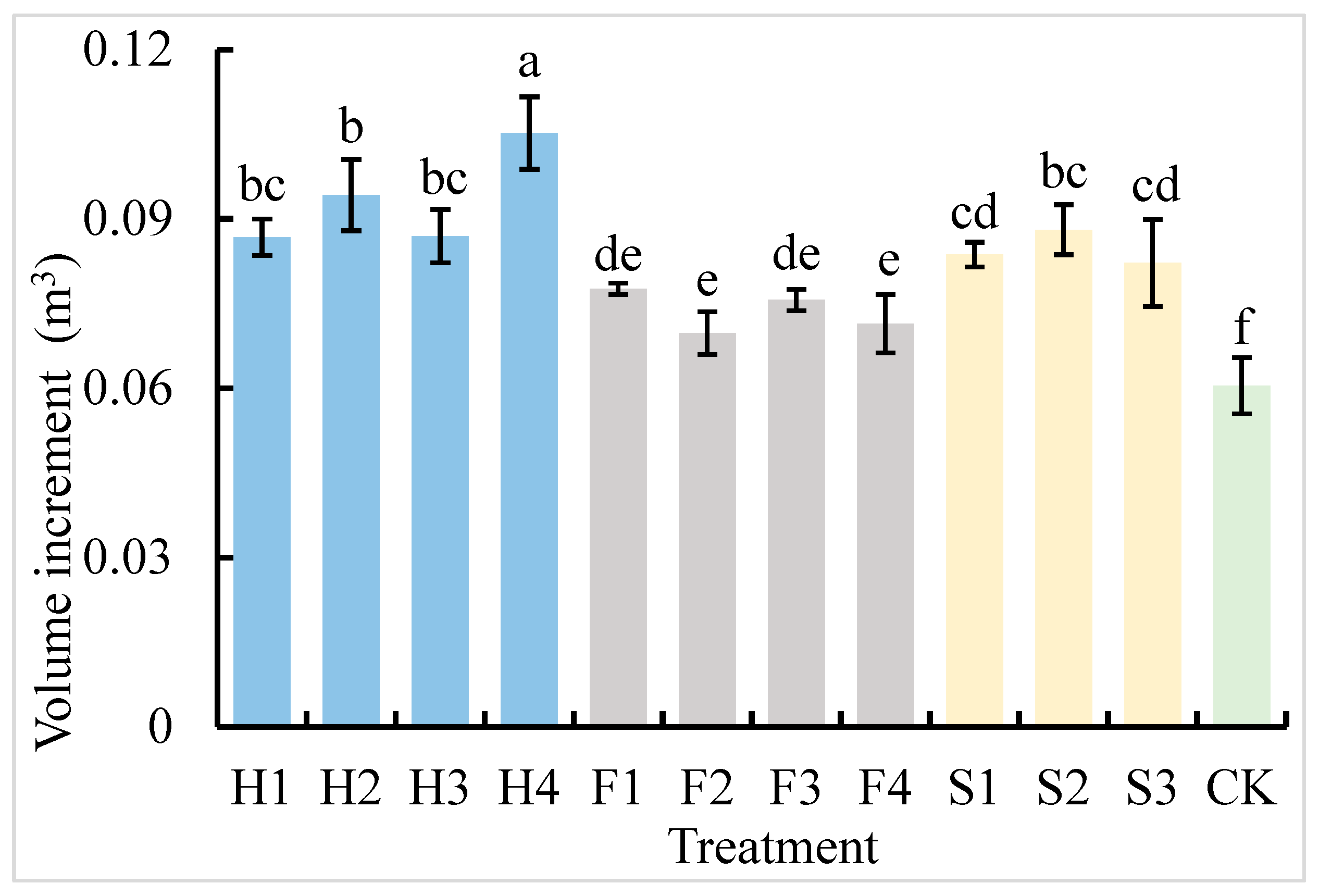
3.2. Volume Change in P. × canadensis ‘Zhongliao 1’
3.3. Soil pH and Physical Properties of P. × canadensis ‘Zhongliao 1’ Plantation
3.4. Soil Nutrient Contents of P. × canadensis ‘Zhongliao 1’ Plantation
3.5. Soil Enzyme Activity of P. × canadensis ‘Zhongliao 1’ Plantation
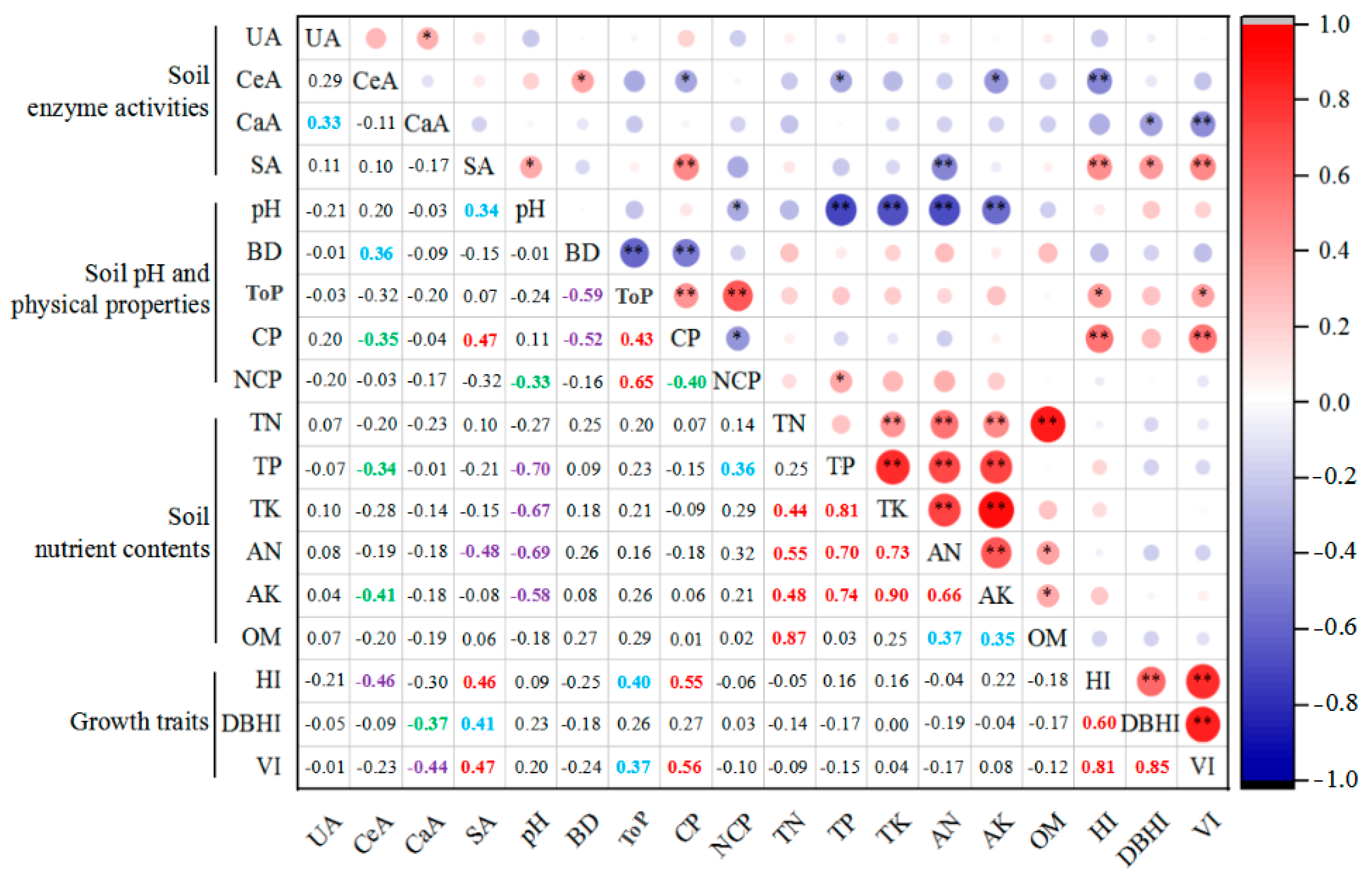
3.6. Correlation Analysis of Soil Physicochemical Properties and Plant Treatments
3.7. Comprehensive Evaluation of Different Treatments of Fertilization and Irrigation Regulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shi, T.; Zhang, X.; Hou, Y.; Jia, C.; Dan, X.; Zhang, Y.; Jiang, Y.; Lai, Q.; Feng, J.; Ma, T.; et al. The super-pangenome of Populus unveil genomic facets for its adaptation and diversification in widespread forest trees. Mol. Plant 2024, 17, 725–746. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Portillo-Estrada, M.; Ceulemans, R. Identifying the best plant water status indicator for bio-nergy poplar genotypes. GCB Bioenergy 2020, 12, 426–444. [Google Scholar] [CrossRef]
- Du, K.; Jiang, S.; Chen, H.; Xia, Y.; Guo, R.; Ling, A.; Liao, T.; Wu, W.; Kang, X. Spatiotemporal miRNA and transcriptomic network dynamically regulate the developmental and senescence processes of poplar leaves. Hortic. Res. 2023, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Bergante, S.; Barbetti, R.; Coaloa, D.; Facciotto, G. Nitrogen fertilization of ‘I-214’ poplar trees with urea and different slow-release fertilizers: Yield, economic and environmental aspects. Biomass Bioenergy 2023, 173, 106806. [Google Scholar] [CrossRef]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; Yin, W.; Xia, X. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. J. Exp. Bot. 2020, 71, 1969–1984. [Google Scholar] [CrossRef]
- Zhang, Q.; Tian, Y.; Wang, M.; Zhai, Z.; Zhou, S. Phenotypic traits differentiations and classifications of the F1 hybrid progenies of Populus deltoides × P. cathayana at the seeding stage. J. Nanjing For. Univ. (Nat. Sci.) 2022, 46, 40–48. (In Chinese) [Google Scholar]
- Guo, Q.; Zhu, Y.; Sun, F.; Korpelainen, H.; Niinemets, Ü.; Li, C. Male, female, and mixed-sex poplar plantations support divergent soil microbial communities. Glob. Chang. Biol. 2024, 30, e17198. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting density and site effects on stem dimensions, stand productivity, biomass partitioning, carbon stocks and soil nutrient supply in hybrid poplar plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef]
- Thomas, A.; Marron, N.; Bonal, D.; Piutti, S.; Dallé, E.; Priault, P. Leaf and tree water-use efficiencies of Populus deltoides × P. nigra in mixed forest and agroforestry plantations. Tree Physiol. 2022, 42, 2432–2445. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Arevalo, C.; Jones, H.; Bhatti, J.S.; Sidders, D. Carbon sequestration and water use of a young hybrid poplar plantation in north-central Alberta. Biomass Bioenergy 2013, 56, 323–333. [Google Scholar] [CrossRef]
- Jia, G.; Chen, L.; Yu, X.; Liu, Z. Soil water stress overrides the benefit of water-use efficiency from rising CO2 and temperature in a cold semi-arid poplar plantation. Plant Cell Environ. 2022, 45, 1172–1186. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.D.; Ning, Q.R.; Guo, J.J.; Gong, X.W.; Zhu, J.J.; Hao, G.Y. Hydraulic limitation underlies the dieback of Populus pseudo-simonii trees in water-limited areas of northern China. For. Ecol. Manag. 2021, 483, 118764. [Google Scholar] [CrossRef]
- Kang, S.; Eltahir, E.A. North China Plain threatened by deadly heatwaves due to climate change and irrigation. Nat. Commun. 2018, 9, 2894. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Tang, L.; Li, C.; Ren, J.; Zhang, L.; Li, X. Dynamics of soil organic carbon and nitrogen and their relations to hydrothermal variability in dryland. J. Environ. Manag. 2022, 319, 115751. [Google Scholar] [CrossRef]
- Chen, X.; Xing, H.; Liu, B.; Wang, Y.; Cui, N.; Wang, Z.; Zhang, Y. Changes induced by multi-stage water stress on maize growth, water and nitrogen utilization and hormone signaling under different nitrogen supplies. Agric. Water Manag. 2023, 290, 108570. [Google Scholar] [CrossRef]
- Surendran, U.; Chandran, K.M. Development and evaluation of drip irrigation and fertigation scheduling to improve water productivity and sustainable crop production using HYDRUS. Agric. Water Manag. 2022, 269, 107668. [Google Scholar] [CrossRef]
- Wang, N.; Zahng, T.; Cong, A.; Lian, J. Integrated application of fertilization and reduced irrigation improved maize (Zea mays L.) yield, crop water productivity and nitrogen use efficiency in a semi-arid region. Agric. Water Manag. 2023, 289, 108566. [Google Scholar] [CrossRef]
- Li, D.; Fernández, J.E.; Li, X.; Xi, B.; Jia, L.; Hernandez-Santana, V. Tree growth patterns and diagnosis of water status based on trunk diameter fluctuations in fast-growing Populus tomentosa plantations. Agric. Water Manag. 2020, 241, 106348. [Google Scholar] [CrossRef]
- Yan, X.L.; Dai, T.F.; Jia, L.M. Evaluation of the cumulative effect of drip irrigation and fertigation on productivity in a poplar plantation. Ann. For. Sci. 2018, 75, 5. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Qiang, Y.; Lu, R.; Che, C. Estimating non-productive water losses in irrigated Platycladus orientalis plantations in semi-arid mountainous: Based on stable isotopes. Sci. Total Environ. 2024, 934, 173279. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, F.; Zou, M.; Ji, S.; Li, M.; Kang, S. Quantifying the spatial water salinity threshold of saline water irrigation by applying distributed WAVES model. Agric. Syst. 2024, 214, 103839. [Google Scholar] [CrossRef]
- Souza, J.L.B.; Antonangelo, J.A.; Zhang, H.; Reed, V.; Finch, B.; Arnall, B. Impact of long-term fertilization in no-till on the stratification of soil acidity and related parameters. Soil Till. Res. 2023, 228, 105624. [Google Scholar] [CrossRef]
- Yang, T.; Li, D.; Clothier, B.; Wang, Y.; Duan, J.; Di, N.; Li, G.; Li, X.; Ji, L.; Xi, B.; et al. Where to monitor the soil-water potential for scheduling drip irrigation in Populus tomentosa plantations located on the North China Plain? For. Ecol. Manag. 2019, 437, 99–112. [Google Scholar] [CrossRef]
- Tuo, Y.; Wang, Z.; Zheng, Y.; Shi, X.; Liu, X.; Ding, M.; Yang, Q. Effect of water and fertilizer regulation on the soil microbial biomass carbon and nitrogen, enzyme activity, and saponin content of Panax notoginseng. Agric. Water Manag. 2023, 278, 108145. [Google Scholar] [CrossRef]
- Li, X.; Simunek, J.; Shi, H.; Yan, J.; Peng, Z.; Gong, X. Spatial distribution of soil water, soil temperature, and plant roots in a drip-irrigated intercropping field with plastic muich. Eur. J. Agron. 2017, 83, 47–56. [Google Scholar] [CrossRef]
- Yahgi, T.; Arslan, A.; Naoum, F. Cucumber (Cucumis sativus, L.) water use efficiency (WUE) under plastic mulch and drip irrigation. Agric. Water Manag. 2013, 115, 232–241. [Google Scholar]
- Abou Lila, T.S.; Berndtsson, R.; Persson, M.; Somaida, M.; Ei-Kiki, M.; Hamed, Y.; Mirdan, A. Numerical evaluation of subsurface trickle irrigation with brackish water. Irrig. Sci. 2013, 31, 1125–1137. [Google Scholar] [CrossRef]
- Cao, Y.; Cai, H.; Sun, S.; Gu, X.; Mu, Q.; Duan, W.; Zhao, Z. Effects of drip irrigation methods on yield and water productivity of maize in Northwest China. Agric. Water Manag. 2022, 259, 107227. [Google Scholar] [CrossRef]
- Skaggs, T.H.; Trout, T.J.; Rothfuss, Y. Drip irrigation water distribution patterns: Effects of emitter rate, pulsing, and antecedent water. Irrig. Sci. 2010, 74, 1886–1896. [Google Scholar] [CrossRef]
- Du, Y.; Liu, X.; Zhang, L.; Zhou, W. Drip irrigation in agricultural saline-alkali land controls soil salinity and improves crop yield: Evidence from a global meta-analysis. Sci. Total Environ. 2023, 880, 163226. [Google Scholar] [CrossRef]
- Patel, N.; Rajput, T.B.S. Dynamics and modeling of soil water under subsurface drip irrigated onion. Agric. Water Manag. 2008, 95, 1335–1349. [Google Scholar] [CrossRef]
- Chen, H.; Pang, X.; Zhang, R.; Zhang, Z.; Xu, Q.; Fang, C.; Li, J.; Lin, W. Effects of different irrigation and fertilizer application regimes on soil enzyme activities and microbial functional diversity in rhizosphere of ratooning rice. Acta Agron. Sin. 2017, 43, 1507–1517. (In Chinese) [Google Scholar] [CrossRef]
- He, Y.; Li, G.; Xi, B.; Zhao, H.; Jia, L. Fine root plasticity of young Populus tomentosa plantations under drip irrigation and nitrogen fertigation in the North China Plain. Agric. Water Manag. 2022, 261, 107341. [Google Scholar] [CrossRef]
- Yan, Z.; Chen, S.; Li, J.; Alva, A.; Chen, Q. Manure and nitrogen application enhances soil phosphorus mobility in calcareous soil in greenhouses. J. Environ. Manag. 2016, 181, 26–35. [Google Scholar] [CrossRef]
- He, Y.; Xi, B.; Bloomberg, M.; Jia, L.; Zhao, D. Effects of drip irrigation and nitrogen fertigation on stand growth and biomass allocation in young triploid Populus tomentosa plantations. For. Ecol. Manag. 2020, 461, 117937. [Google Scholar] [CrossRef]
- Paris, P.; Di Matteo, G.; Tarchi, M.; Tosi, L.; Spaccino, L.; Lauteri, M. Precision subsurface drip irrigation increases yield while sustaining water-use efficiency in Mediterranean poplar bioenergy plantations. For. Ecol. Manag. 2018, 409, 749–756. [Google Scholar] [CrossRef]
- Shock, C.C.; Feibert, E.B.G.; Seddigh, M.; Saunders, L.D. Water requirements and growth of irrigated hybrid poplar in a semi-arid environment in eastern Oregon. West. J. Appl. For. 2002, 17, 46–53. [Google Scholar] [CrossRef]
- Shrive, S.C.; McBride, R.A.; Gordon, A.M. Photosynthetic and growth responses of two broad-leaf tree species to irriga-tion with municipal landfill leachate. J. Environ. Qual. 1994, 23, 534–542. [Google Scholar] [CrossRef]
- Liu, J.; Li, D.; Fernández, J.E.; Coleman, M.; Hu, W.; Di, N. Variations in water-balance components and carbon stocks in poplar plantations with differing water inputs over a whole rotation: Implications for sustainable forest management under climate change. Agric. For. Meteorol. 2022, 320, 108958. [Google Scholar] [CrossRef]
- Mantel, S.; Dondeyne, S.; Deckers, S. World Reference Base for Soil Resources (WRB); Encyclopedia of Soils in the Environment; Academic Press: Oxford, UK, 2014; pp. 206–217. [Google Scholar]
- Lan, Z.; Lv, F.; Fu, J.; He, Y.; Bi, H.; Qin, X. The Invention Relates to a Water-Fertilizer Integrated Fertilization Method for Poplar Drip Irrigation Cultivatio. China Patent No. CN201911082097.4, 3 January 2020. [Google Scholar]
- Peng, P.H.; Kuo, C.H.; Wei, C.H.; Hsieh, Y.T.; Chen, J.C. The relationship between breast height form factor and form quotient of Liquidambar formosana in the eastern part of Taiwan. Forests 2022, 13, 1111. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, J.; Li, X.; Yang, Z.; Zheng, Y.; Wang, J.; Lin, W.; Xie, J.; Chen, Y.; Yang, Y. Are the combined effects of warming and drought on foliar C: N: P: K stoichiometry in a subtropical forest greater than their individual effects? For. Ecol. Manag. 2019, 448, 256–266. [Google Scholar] [CrossRef]
- Zha, T. Soil Physical and Chemical Analysis; Soil Physicochemical Analysis; China Forestry Publishing House: Beijing, China, 2017; pp. 62–78. (In Chinese) [Google Scholar]
- Gao, C.; Liu, C.; Chen, C.; Liu, N.; Liu, F.; Su, X.; Huang, Q. Genetic evaluation of water use efficiency and nutrient use efficiency in Populus deltoides Bartr. ex Marsh. seedlings in China. Plants 2024, 13, 2228. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Dybzinski, R.; Robinson, D. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, H.; Zhao, F.; Chen, C.; Liu, W.; Yang, B.; Zhang, W. Recognizing the role of plant species composition in the modification of soil nutrients and water in rubber agroforestry systems. Sci. Total Environ. 2020, 723, 138042. [Google Scholar] [CrossRef]
- Che, Z.; Wang, J.; Li, J. Modeling strategies to balance salt leaching and nitrogen loss for drip irrigation with saline water in arid regions. Agric. Water Manag. 2022, 274, 107943. [Google Scholar] [CrossRef]
- Ma, C.; Wang, J.; Li, J. Utilization of soil and fertilizer nitrogen supply under mulched drip irrigation with various water qualities in arid regions. Agric. Water Manag. 2023, 280, 108219. [Google Scholar] [CrossRef]
- Rogers, E.R.; Zalesny, R.S.; Lin, C.H.; Vinhal, R.A. Intrinsic and extrinsic factors influencing Populus water use: A literature review. J. Environ. Manag. 2023, 348, 119180. [Google Scholar] [CrossRef]
- Niu, M.X.; Feng, C.H.; He, F.; Zhang, H.; Bao, Y.; Liu, S.J.; Liu, X.; Su, Y.; Liu, C.; Wang, H.L.; et al. The miR6445-NAC029 module regulates drought tolerance by regulating the expression of glutathione S-transferase U23 and reactive oxygen species scavenging in Populus. New Phytol. 2024, 242, 2043–2058. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Clothier, B.; Coleman, M.; Duan, J.; Hu, W.; Li, D.; Di, N.; Liu, Y.; Fu, J.; Li, J.; et al. Irrigation management in poplar (Populus spp.) plantations: A review. For. Ecol. Manag. 2021, 494, 119330. [Google Scholar] [CrossRef]
- Sun, S.; He, C.; Qiu, L.; Li, C.; Zhang, J.; Meng, P. Stable isotope analysis reveals prolonged drought stress in poplar plantation mortality of the Three-North Shelter Forest in Northern China. Agric. For. Meteorol. 2018, 252, 39–48. [Google Scholar] [CrossRef]
- Wang, C.; Sun, Y.; Zou, X.; Chen, H.Y.Y.; Delgado-Baquerizo, M.; Yang, J.; Wang, G.; Liu, Y.; Ruan, H. Increased fine root production coupled with reduced aboveground production of plantations under a three-year experimental drought. Sci. Total Environ. 2024, 908, 168370. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Li, Z.; Xiang, Y.; Sun, T.; Liu, X.; Shi, H.; Li, W.; Huang, X.; Tang, Z.; Lu, J.; et al. Drip fertigation increases maize grain yield by affecting phenology, grain filling process, biomass accumulation and translocation: A 4-year field trial. Plants 2024, 13, 1903. [Google Scholar] [CrossRef]
- Dhayal, D.; Lal, K.; Khanna, M.; Sudhishri, S.; Brar, A.S.; Sindhu, V.K.; Singh, M.; Bhattacharyya, R.; Rajath, E.; Rosin, K.G.; et al. Performance of surface and subsurface drip fertigated wheat-moongbean-maize cropping system under different irrigation schedules and nutrient doses. Agric. Water Manag. 2023, 284, 108338. [Google Scholar] [CrossRef]
- Krumland, B.E.; Wensel, L.C. A generalized height-diameter equation for coastal California species. West. J. Appl. For. 1988, 3, 113–115. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Fang, H.; Shi, L.; Yue, X.; Yang, Q. Exploring the coupling mode of irrigation method and fertilization rate for improving growth and water-fertilizer use efficiency of young mango tree. Sci. Hortic. 2021, 286, 110211. [Google Scholar] [CrossRef]
- Fu, Y.; Ding, G.; Quan, W.; Zhao, X.; Tao, Q. Coupling optimization of water-fertilizer for coordinated development of the environment and growth of Pinus massoniana seedlings. Agric. Water Manag. 2024, 300, 108895. [Google Scholar] [CrossRef]
- Zhu, J.; Bo, H.; Li, X.; Wen, C.; Wang, J.; Nie, L.; Tian, J.; Song, L. Long term water-nitrogen coupling effect on stand volume of different clones of Populus tomentosa. Sci. Silv. Sin. 2019, 55, 27–35. (In Chinese) [Google Scholar]
- Yang, H.; Wang, Y.; Jia, L. Response of pulp plantation growth of Populus tomentosa S86 in short rotation period to coupling of water and fertilizer in furrow irrigation. J. Beijing For. Univ. 2023, 4, 68–78. (In Chinese) [Google Scholar]
- Coyle, D.R.; Coleman, M.D. Forest production responses to irrigation and fertilization are not explained by shifts in allocation. For. Ecol. Manag. 2005, 208, 137–152. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, X.; Liu, W.; Feng, B.; Lv, W.; Zhang, Z.; Yang, X.; Dong, Q. Plant biomass partitioning in alpine meadows under different herbivores asinfluenced by soil bulk density and available nutrients. Catena 2024, 240, 108017. [Google Scholar] [CrossRef]
- Liu, H.; Fu, X. Phytohormonal networks facilitate plant root developmental adaptations to environmental changes. Sci. Bull. 2024, 69, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xiao, G.; Bol, R.; Wang, L.; Zhuge, Y.; Wu, W.; Li, H.; Meng, F. Influences of irrigation and fertilization on soil N cycle and losses from Wheat-maize cropping system in northern China. Environ. Pollut. 2021, 278, 116852. [Google Scholar] [CrossRef] [PubMed]
- Zong, R.; Wang, Z.; Li, W.; Li, H.; Ayantobo, O.O. Effects of practicing long-term mulched drip irrigation on soil quality in Northwest China. Sci. Total Environ. 2023, 878, 163247. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.; Zhu, Q.; Zeng, M.; Shen, J.; Shi, X.; Liu, X.; de Vries, W. Impacts of nitrogen fertilizer type and application rate on soil acidification rate under a wheat-maize double cropping system. J. Environ. Manag. 2020, 270, 110888. [Google Scholar] [CrossRef]
- Wang, Y.K.; Zhang, Z.B.; Jiang, F.H.; Guo, Z.C.; Peng, X.H. Evaluating soil physical quality indicators of a Vertisol as affected by different tillage practices under wheat-maize system in the North China Plain. Soil Till. Res. 2021, 209, 101970. [Google Scholar] [CrossRef]
- Balakhnina, T.; Bennicelli, R.; Stêpinewska, Z.; Stêpniewski, W.; Borkowska, A.; Fomina1, I. Stress responses of spring rape plants to soil flooding. Int. Agrophys. 2012, 26, 347–353. [Google Scholar] [CrossRef]
- Wan, S.Z.; Gu, H.J.; Yang, Q.P.; Hu, X.F.; Fang, X.M.; Singh, A.N.; Chen, F.S. Long-term fertilization increases soil nutrient accumulations but decreases biological activity in navel orange orchards of subtropical China. J. Soil. Sediment. 2017, 17, 2346–2356. [Google Scholar] [CrossRef]
- Liu, G.; Wang, H.; Yan, G.; Wang, M.; Jiang, S.; Wang, X.; Xue, J.; Xu, M.; Xing, Y.; Wang, Q. Soil enzyme activities and microbial nutrient limitation during thesecondary succession of boreal forests. Catena 2023, 230, 107268. [Google Scholar] [CrossRef]
- Kotroczo, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Tóthmérész, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, S.; Hu, T.; Li, H.; Sun, J.; Sun, G.; Liu, J. Responses of soil reactive nitrogen pools and enzyme activities to water and nitrogen levels and their relationship with apple yield and quality under drip fertigation. Sci. Hortic. 2024, 324, 112632. [Google Scholar] [CrossRef]
- Xu, G.; Lu, D.; Sun, H.; Wang, H.; Li, Y. Effect of alternative wetting and drying irrigation and nitrogen coupling on rhizosphere environment of rice. Trans. CSAE 2017, 33, 186–194. [Google Scholar]
- Zuccarini, P.; Sardans, J.; Asensio, L.; Peñuelas, J. Altered activities of extracellular soil enzymes by the interacting global environmental changes. Glob. Chang. Biol. 2023, 29, 2067–2091. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.A.; Suir, E.; Duong, G.H.; Hulster, E.D.; Pronk, J.T.; Maris, A.J. Catalase overexpression reduces lactic acid induced oxidative stress in Saccharomyces cerevisiae. Appl. Environ. Microb. 2009, 75, 2320–2325. [Google Scholar] [CrossRef]
- Du, Y.D.; Zhang, Q.; Cui, B.J.; Sun, J.; Wang, Z.; Ma, L.H.; Niu, W.Q. Aerated irrigation improves tomato yield and nitrogen use efficiency while reducing nitrogen application rate. Agric. Water Manag. 2020, 235, 106152. [Google Scholar] [CrossRef]
- Liang, Y.; Ning, D.; Lu, Z.; Zhang, N.; Hale, L.; Wu, L.; Clark, I.M.; McGrath, S.P.; Storkey, J.; Hirsch, P.R.; et al. Century long fertilization reduces stochasticity controlling grassland microbial community succession. Soil Biol. Biochem. 2020, 151, 108023. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, S.; Hu, Y.; Chen, Y.; Jiang, H.; Duan, B.; Lu, X. Links between chemical composition of soil organic matter and soil enzyme activity in alpine grassland ecosystems of the Tibetan Plateau. Catena 2022, 218, 106565. [Google Scholar] [CrossRef]
- Flexas, J.; Carriquí, M.; Coopman, R.E.; Gago, J.; Galmés, J.; Martorell, S.; Morales, F.; Diaz-Espejo, A. Stomatal and mesophyll conductances to CO2 in different plant groups: Underrated factors for predicting leaf photosynthesis responses to climate change? Plant Sci. 2014, 226, 41–48. [Google Scholar] [CrossRef]
- Chenge, I.B. Height-diameter relationship of trees in Omo strict nature forest reserve, Nigeria. Trees For. People 2021, 3, 100051. [Google Scholar] [CrossRef]
- Zellweger, F.; De Frenne, P.; Lenoir, J.; Vangansbeke, P.; Verheyen, K.; Bernhardt-Romeramm, M.; Baeten, L.; Hedl, R.; Berki, I.; Brunet, J.; et al. Forest microclimate dynamics drive plant responses to warming. Science 2020, 368, 772–775. [Google Scholar] [CrossRef]
- Schärer, M.L.; Lüscher, A.; Kahmen, A. Post-drought compensatory growth in perennial grasslands is determined by legacy effects of the soil and not by plants. New Phytol. 2023, 240, 2265–2275. [Google Scholar] [CrossRef]
- Liu, J.; Han, C.; Zhao, Y.; Yang, J.; Cade-Menun, B.J.; Hu, Y.; Li, J.; Liu, H.; Sui, P.; Chen, Y.; et al. The chemical nature of soil phosphorus in response to long-term fertilization practices: Implications for sustainable phosphorus management. J. Clean. Prod. 2020, 272, 123093. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fertigation Amounts (g) | Drip Irrigation Thresholds | |||
|---|---|---|---|---|---|
| N | P | K | |||
| Drip irrigation and fertilization | H1 | 643.20 | 473.37 | 492.29 | 60% of field capacity |
| H2 | 643.20 | 473.37 | 492.29 | 80% of field capacity | |
| H3 | 1029.12 | 757.39 | 787.66 | 60% of field capacity | |
| H4 | 1029.12 | 757.39 | 787.66 | 80% of field capacity | |
| Only fertilization | F1 | 450.24 | 331.36 | 344.60 | non-irrigation |
| F2 | 643.20 | 473.37 | 492.29 | non-irrigation | |
| F3 | 836.16 | 615.38 | 639.97 | non-irrigation | |
| F4 | 1029.12 | 757.39 | 787.66 | non-irrigation | |
| Only drip irrigation | S1 | 0 | 0 | 0 | 80% of field capacity |
| S2 | 0 | 0 | 0 | 70% of field capacity | |
| S3 | 0 | 0 | 0 | 60% of field capacity | |
| Non-irrigation and non-fertigation | CK | 0 | 0 | 0 | non-irrigation |
| Organs | Biomass (kg) | Cumulative Absorption of Nutrient Elements (g) | Annual Absorption of Nutrient Elements (g) | ||||
|---|---|---|---|---|---|---|---|
| N | P | K | N | P | K | ||
| Trunk | 307.99 | 1187.74 | 803.17 | 839.66 | 643.20 | 473.37 | 492.29 |
| 70% of Annual Absorption of Nutrient Elements (g) | |||||||
| Branch | 124.65 | 779.95 | 474.27 | 588.24 | N | P | K |
| 450.24 | 331.36 | 344.60 | |||||
| Root | 54.58 | 371.40 | 296.06 | 414.55 | 130% of Annual Absorption of Nutrient Elements (g) | ||
| N | P | K | |||||
| 836.16 | 615.38 | 639.97 | |||||
| Leaf | 14.71 | 325.81 | 85.69 | 131.55 | 160% of Annual Absorption of Nutrient Elements (g) | ||
| Total | 1416.27 | 2264.92 | 1659.18 | 1973.82 | N | P | K |
| 1029.12 | 757.39 | 787.66 | |||||
| Treatments | H (m) | DBH (cm) | V (m3) | |||
|---|---|---|---|---|---|---|
| May | October | May | October | May | October | |
| H1 | 19.54 ± 0.30 | 20.93 ± 0.37 | 25.97 ± 0.53 | 27.40 ± 0.46 | 0.431 ± 0.03 | 0.518 ± 0.03 |
| H2 | 19.31 ± 0.13 | 20.86 ± 0.17 | 25.23 ± 0.34 | 26.88 ± 0.42 | 0.402 ± 0.01 | 0.496 ± 0.02 |
| H3 | 18.55 ± 0.09 | 20.00 ± 0.07 | 25.42 ± 0.17 | 26.95 ± 0.10 | 0.394 ± 0.01 | 0.481 ± 0.01 |
| H4 | 20.53 ± 0.38 | 22.18 ± 0.39 | 25.55 ± 0.49 | 27.28 ± 0.54 | 0.442 ± 0.02 | 0.547 ± 0.03 |
| F1 | 20.53 ± 0.38 | 21.99 ± 0.04 | 25.57 ± 0.16 | 26.86 ± 0.13 | 0.395 ± 0.01 | 0.473 ± 0.01 |
| F2 | 20.54 ± 0.42 | 21.75 ± 0.18 | 25.99 ± 0.44 | 27.20 ± 0.40 | 0.407 ± 0.02 | 0.477 ± 0.02 |
| F3 | 18.30 ± 0.23 | 19.74 ± 0.27 | 25.18 ± 0.28 | 26.53 ± 0.27 | 0.372 ± 0.01 | 0.447 ± 0.01 |
| F4 | 19.04 ± 0.22 | 20.34 ± 0.30 | 25.19 ± 0.30 | 26.45 ± 0.29 | 0.392 ± 0.01 | 0.464 ± 0.02 |
| S1 | 19.65 ± 0.13 | 21.07 ± 0.15 | 24.81 ± 0.29 | 26.34 ± 0.29 | 0.388 ± 0.01 | 0.472 ± 0.01 |
| S2 | 21.01 ± 0.40 | 22.52 ± 0.39 | 23.99 ± 0.27 | 25.55 ± 0.26 | 0.387 ± 0.01 | 0.476 ± 0.02 |
| S3 | 19.07 ± 0.24 | 20.37 ± 0.31 | 25.40 ± 1.62 | 26.78 ± 1.04 | 0.438 ± 0.03 | 0.520 ± 0.04 |
| CK | 19.55 ± 0.03 | 20.67 ± 0.19 | 24.99 ± 0.74 | 26.07 ± 0.15 | 0.392 ± 0.02 | 0.453 ± 0.03 |
| Treatments | pHH2O Valve | BD (g·cm−3) | ToP (%) | CP (%) | NCP (%) |
|---|---|---|---|---|---|
| H1 | 6.88 ± 0.17 abc | 1.58 ± 0.05 a | 38.98 ± 1.37 a | 35.24 ± 3.07 a | 3.74 ± 2.94 a |
| H2 | 7.52 ± 0.65 ab | 1.53 ± 0.01 a | 43.76 ± 2.94 a | 33.43 ± 5.28 a | 10.33 ± 3.25 a |
| H3 | 7.47 ± 0.72 ab | 1.61 ± 0.03 a | 38.62 ± 4.70 a | 31.44 ± 2.59 ab | 7.18 ± 2.81 a |
| H4 | 7.53 ± 0.63 a | 1.60 ± 0.09 a | 43.14 ± 1.24 a | 34.77 ± 0.34 a | 8.37 ± 0.96 a |
| F1 | 6.12 ± 0.30 cd | 1.58 ± 0.07 a | 41.30 ±2.94 a | 31.92 ± 1.97 ab | 9.38 ± 1.03 a |
| F2 | 6.71 ± 0.16 bc | 1.57 ± 0.04 a | 40.74 ± 3.11 a | 31.44 ± 1.76 ab | 9.31 ± 4.07 a |
| F3 | 7.37 ± 0.32 ab | 1.59 ± 0.02 a | 37.74 ±1.95 a | 33.23 ± 2.58 a | 4.51 ± 0.76 a |
| F4 | 5.44 ± 0.23 d | 1.62 ± 0.07 a | 44.05 ± 9.66 a | 30.14 ± 1.50 ab | 13.91 ± 8.91 a |
| S1 | 6.46 ± 0.34 c | 1.57 ± 0.08 a | 40.62 ± 2.99 a | 31.97 ± 0.21 ab | 8.66 ± 2.83 a |
| S2 | 6.53 ± 0.59 c | 1.66 ± 0.04 a | 38.42 ± 2.69 a | 27.79 ± 1.80 ab | 10.63 ± 4.25 a |
| S3 | 6.06 ± 0.38 cd | 1.62 ± 0.03 a | 40.29 ± 1.51 a | 32.24 ± 2.71 ab | 8.05 ± 4.15 a |
| CK | 7.55 ± 0.35 a | 1.68 ± 0.12 a | 34.14 ± 5.69 a | 24.62 ± 3.86 b | 9.52 ± 5.69 a |
| Treatments | TN (g·kg−1) | TP (g·kg−1) | TK (g·kg−1) | AN (mg·kg−1) | AK (mg·kg−1) | OM (g·kg−1) |
|---|---|---|---|---|---|---|
| H1 | 1.77 ± 0.38 abc | 0.37 ± 0.03 d | 10.39 ± 0.74 c | 94.17 ± 18.74 cd | 189.34 ± 35.62 c | 28.81 ± 6.17 b |
| H2 | 1.49 ± 0.08 c | 0.44 ± 0.08 cd | 10.74 ± 1.00 bc | 76.76 ± 19.33 d | 232.31 ± 42.43 bc | 24.25 ± 0.77 b |
| H3 | 1.55 ± 0.36 c | 0.49 ± 0.17 cd | 11.33 ± 2.20 bc | 107.45 ± 10.84 c | 333.30 ± 114.09 bc | 27.65 ± 7.24 b |
| H4 | 1.85 ± 0.30 abc | 0.51 ± 0.13 cd | 12.08 ± 0.84 bc | 95.24 ± 8.30 cd | 357.18 ± 120.63 bc | 30.08 ± 5.63 ab |
| F1 | 1.73 ± 0.20 abc | 0.81 ± 0.05 b | 13.18 ± 2.32 b | 115.53 ± 16.99 c | 416.29 ± 152.87 ab | 28.52 ± 1.89 b |
| F2 | 1.98 ± 0.27 ab | 0.45 ± 0.07 cd | 10.95 ± 1.78 bc | 111.64 ± 17.10 c | 342.75 ± 54.85 bc | 37.80 ± 7.43 a |
| F3 | 1.61 ± 0.08 bc | 0.52 ± 0.08 cd | 10.12 ± 0.27 c | 75.67 ± 18.65 d | 212.97 ± 32.26 c | 24.73 ± 3.88 b |
| F4 | 2.12 ± 0.06 a | 0.96 ± 0.07 a | 15.57 ± 2.28 a | 277.57 ± 18.05 a | 540.14 ± 140.81 a | 32.38 ± 0.54 ab |
| S1 | 1.67 ± 0.125 bc | 0.61 ± 0.09 c | 12.13 ± 0.84 bc | 97.16 ± 13.51 cd | 301.92 ± 49.94 bc | 26.81 ± 4.22 b |
| S2 | 1.51 ± 0.09 c | 0.52 ± 0.02 cd | 11.02 ± 0.98 bc | 118.37 ± 8.62 c | 244.32 ± 105.77 bc | 26.20 ± 3.55 b |
| S3 | 1.58 ± 0.07 bc | 0.49 ± 0.07 cd | 11.71 ± 0.36 bc | 145.67 ± 1.12 b | 287.62 ± 134.73 bc | 27.68 ± 3.11 b |
| CK | 1.47 ± 0.19 c | 0.49 ± 0.09 cd | 10.68 ± 0.61 bc | 95.77 ± 15.44 cd | 189.13 ± 73.49 c | 23.78 ± 4.32 b |
| Treatments | UA (mg·g−1) | CeA (U·g−1) | CaA (mg·g−1) | SA (mg·g−1) |
|---|---|---|---|---|
| H1 | 0.47 ± 0.03 abc | 16.87 ± 2.52 cd | 7.62 ± 0.32 bc | 52.23 ± 4.35 def |
| H2 | 0.43 ± 0.05 bcd | 15.54 ± 1.23 cde | 8.17 ± 0.24 ab | 56.69 ± 3.43 cde |
| H3 | 0.39 ± 0.03 cd | 13.02 ± 1.83 e | 7.79 ± 0.30 bc | 42.95 ± 3.06 gh |
| H4 | 0.40 ± 0.05 cd | 19.00 ± 2.25 bc | 5.12 ± 0.42 e | 79.11 ± 4.25 a |
| F1 | 0.40 ± 0.03 cd | 13.19 ± 2.09 e | 8.16 ± 0.32 ab | 58.32 ± 4.59 cd |
| F2 | 0.41 ± 0.05 cd | 15.70 ± 1.15 cde | 7.18 ± 0.33 cd | 51.73 ± 5.02 def |
| F3 | 0.42 ± 0.02 bcd | 18.19 ± 1.05 bc | 8.56 ± 0.39 a | 71.22 ± 3.90 b |
| F4 | 0.43 ± 0.04 bcd | 14.19 ± 1.84 de | 6.98 ± 0.44 d | 37.08 ± 3.24 h |
| S1 | 0.49 ± 0.05 ab | 16.72 ± 1.12 cd | 7.61 ± 0.39 bc | 63.37 ± 4.48 c |
| S2 | 0.35 ± 0.05 d | 17.59 ± 2.21 bcd | 6.59 ± 0.26 d | 48.34 ± 1.70 fg |
| S3 | 0.52 ± 0.05 a | 20.97 ± 1.86 ab | 7.65 ± 0.37 bc | 49.91 ± 2.86 ef |
| CK | 0.42 ± 0.06 bcd | 23.59 ± 2.46 a | 7.67 ± 0.33 bc | 40.52 ± 4.57 h |
| Treatments | Comprehensive Score | Ranking | Treatments | Comprehensive Score | Ranking |
|---|---|---|---|---|---|
| H1 | 0.297 | 11 | F3 | 0.319 | 8 |
| H2 | 0.313 | 9 | F4 | 0.706 | 1 |
| H3 | 0.330 | 7 | S1 | 0.426 | 4 |
| H4 | 0.518 | 2 | S2 | 0.301 | 10 |
| F1 | 0.485 | 3 | S3 | 0.416 | 5 |
| F2 | 0.363 | 6 | CK | 0.175 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, N.; Yang, L.; Liu, N.; Peng, R.; Yu, L.; Liu, F.; Wang, S.; Gao, C.; Ji, J.; et al. Effects of Fertilization and Drip Irrigation on the Growth of Populus × canadensis ‘Zhongliao 1’ Plantation and on Soil Physicochemical Properties and Enzyme Activities. Forests 2024, 15, 1651. https://doi.org/10.3390/f15091651
Zhang Y, Wang N, Yang L, Liu N, Peng R, Yu L, Liu F, Wang S, Gao C, Ji J, et al. Effects of Fertilization and Drip Irrigation on the Growth of Populus × canadensis ‘Zhongliao 1’ Plantation and on Soil Physicochemical Properties and Enzyme Activities. Forests. 2024; 15(9):1651. https://doi.org/10.3390/f15091651
Chicago/Turabian StyleZhang, Yan, Nairui Wang, Lingyu Yang, Ning Liu, Rusheng Peng, Lei Yu, Fenfen Liu, Shiqi Wang, Chengcheng Gao, Jiabao Ji, and et al. 2024. "Effects of Fertilization and Drip Irrigation on the Growth of Populus × canadensis ‘Zhongliao 1’ Plantation and on Soil Physicochemical Properties and Enzyme Activities" Forests 15, no. 9: 1651. https://doi.org/10.3390/f15091651
APA StyleZhang, Y., Wang, N., Yang, L., Liu, N., Peng, R., Yu, L., Liu, F., Wang, S., Gao, C., Ji, J., Liu, C., & Liang, D. (2024). Effects of Fertilization and Drip Irrigation on the Growth of Populus × canadensis ‘Zhongliao 1’ Plantation and on Soil Physicochemical Properties and Enzyme Activities. Forests, 15(9), 1651. https://doi.org/10.3390/f15091651