
Vegetation Health in China Is Severely Compromised by Drought, Wet and Heat Stress Events

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data
2.3. Methods
2.3.1. Construction of the Vegetation Health Assessment System
2.3.2. Model Effectiveness Verification
2.3.3. Trend Analysis
2.3.4. Identification of Stress Events
2.3.5. Sensitivity Analysis
2.3.6. Modified Residual Analysis
2.3.7. Statistical Methods
3. Results
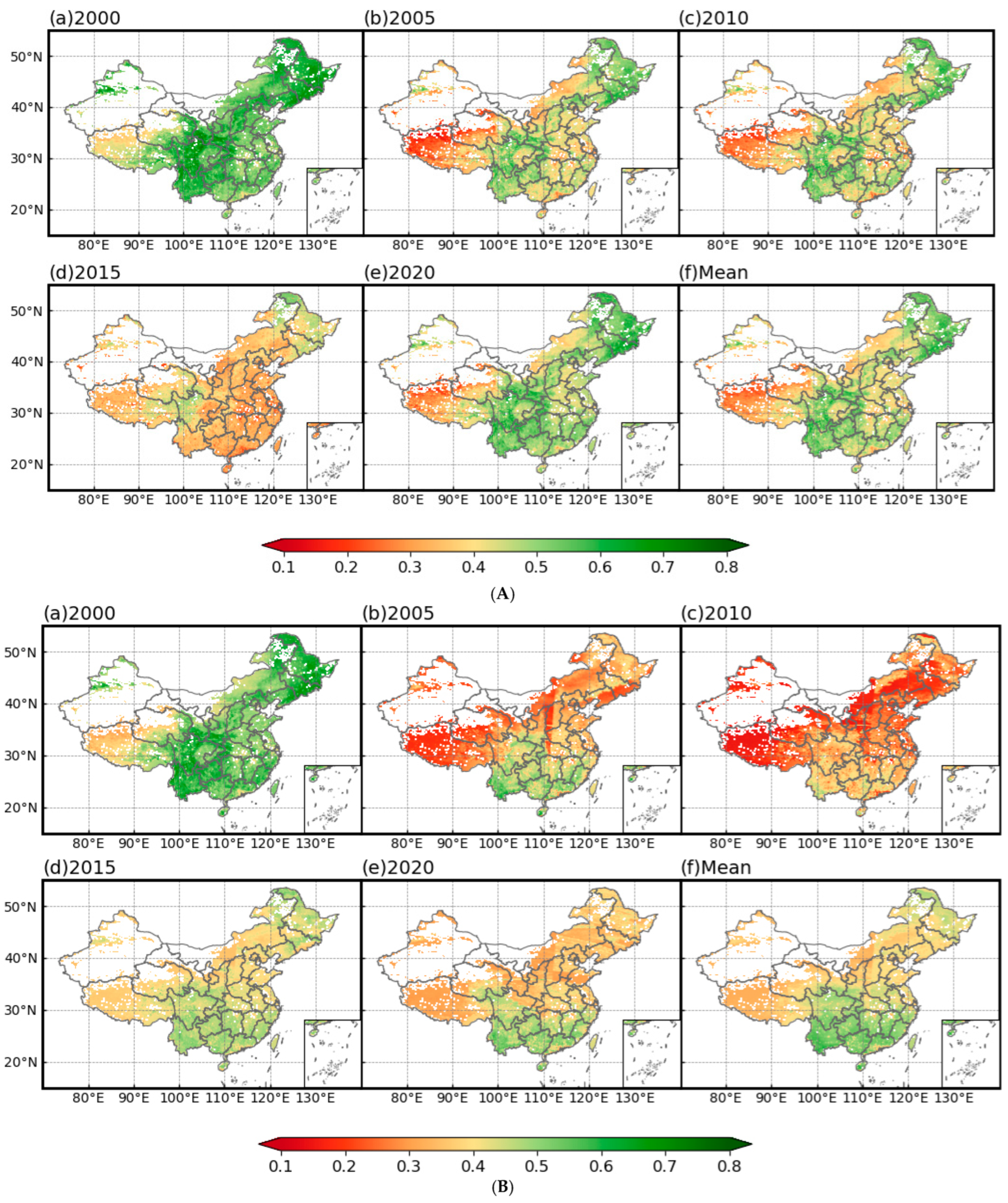
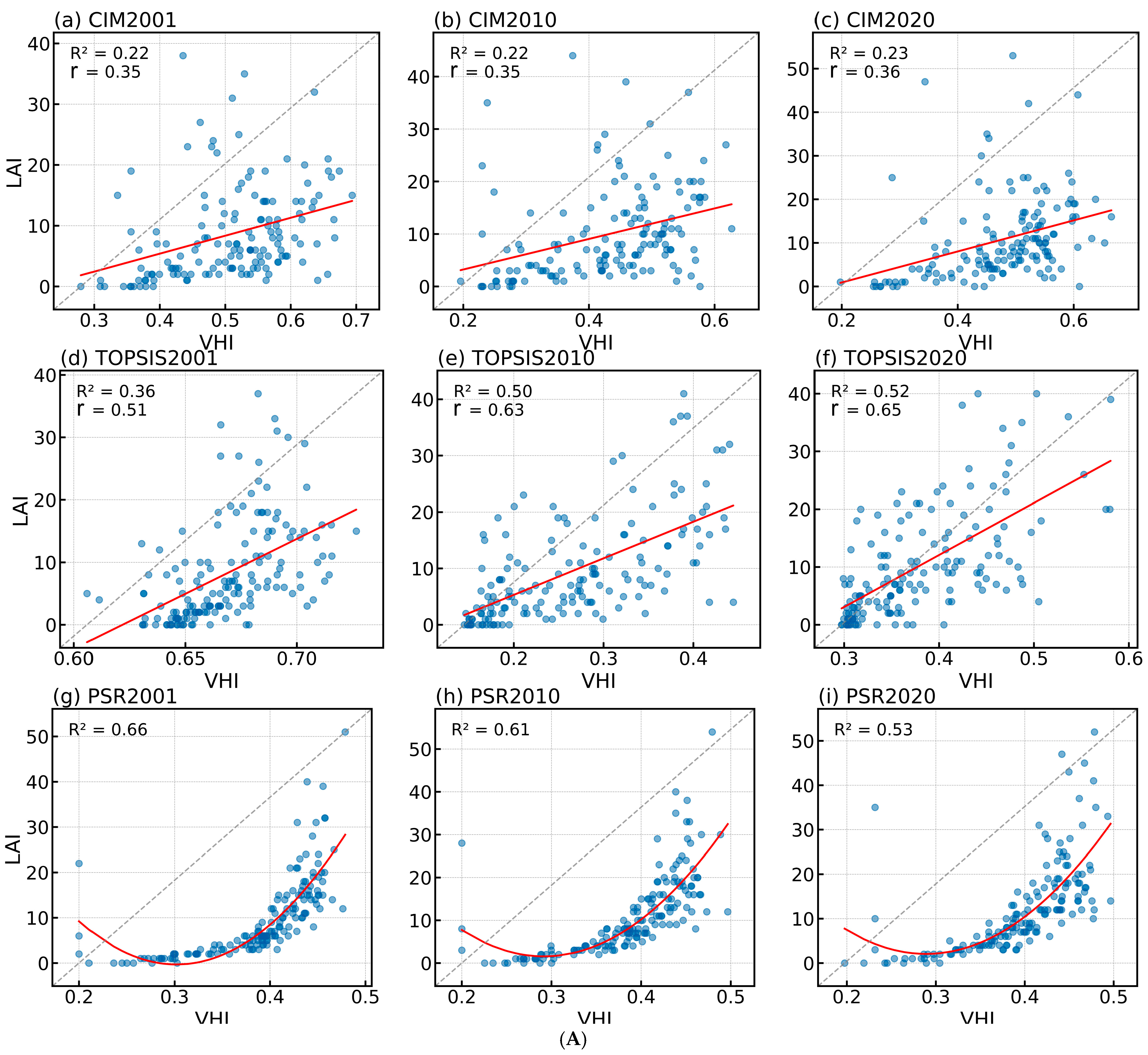
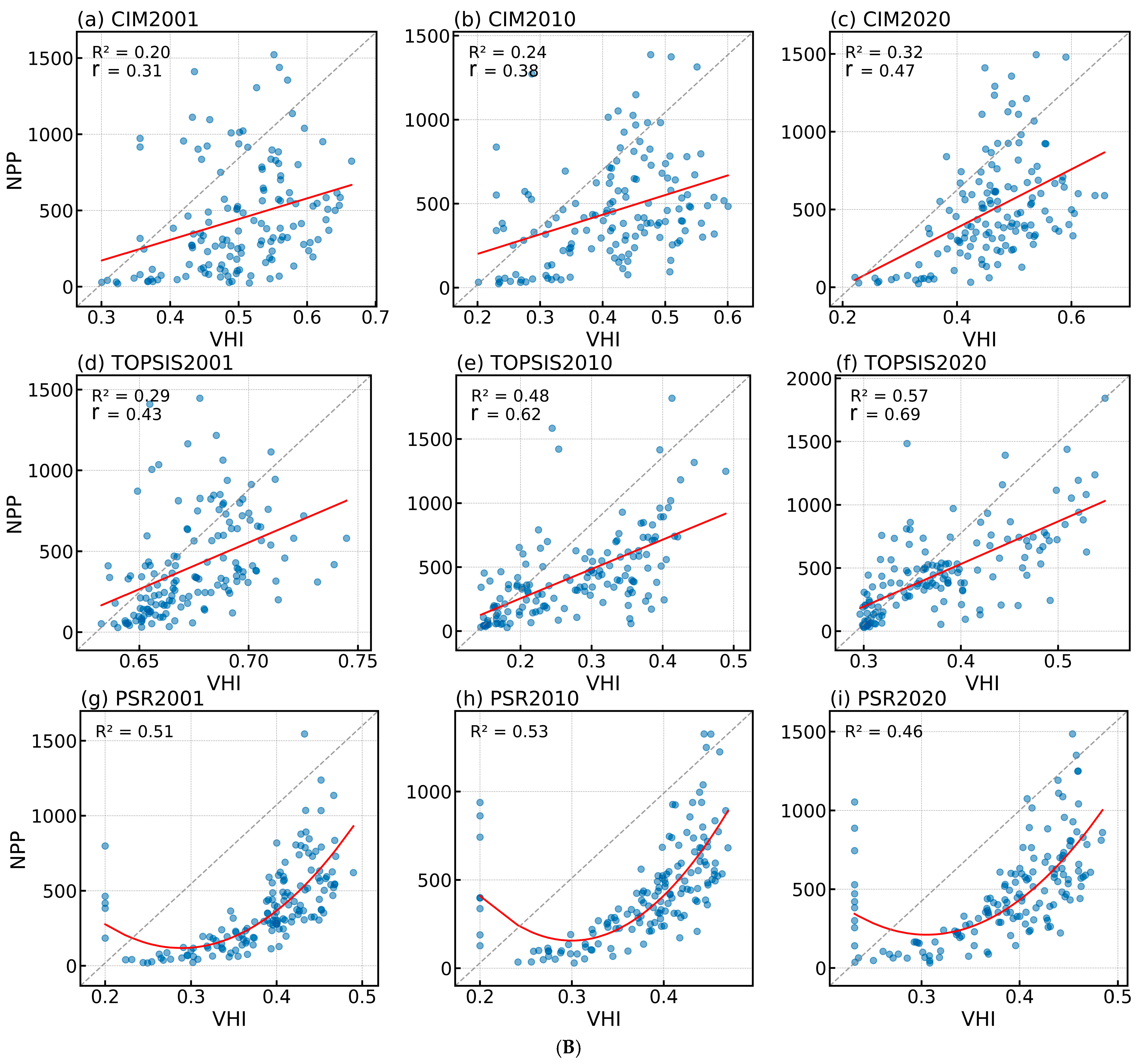
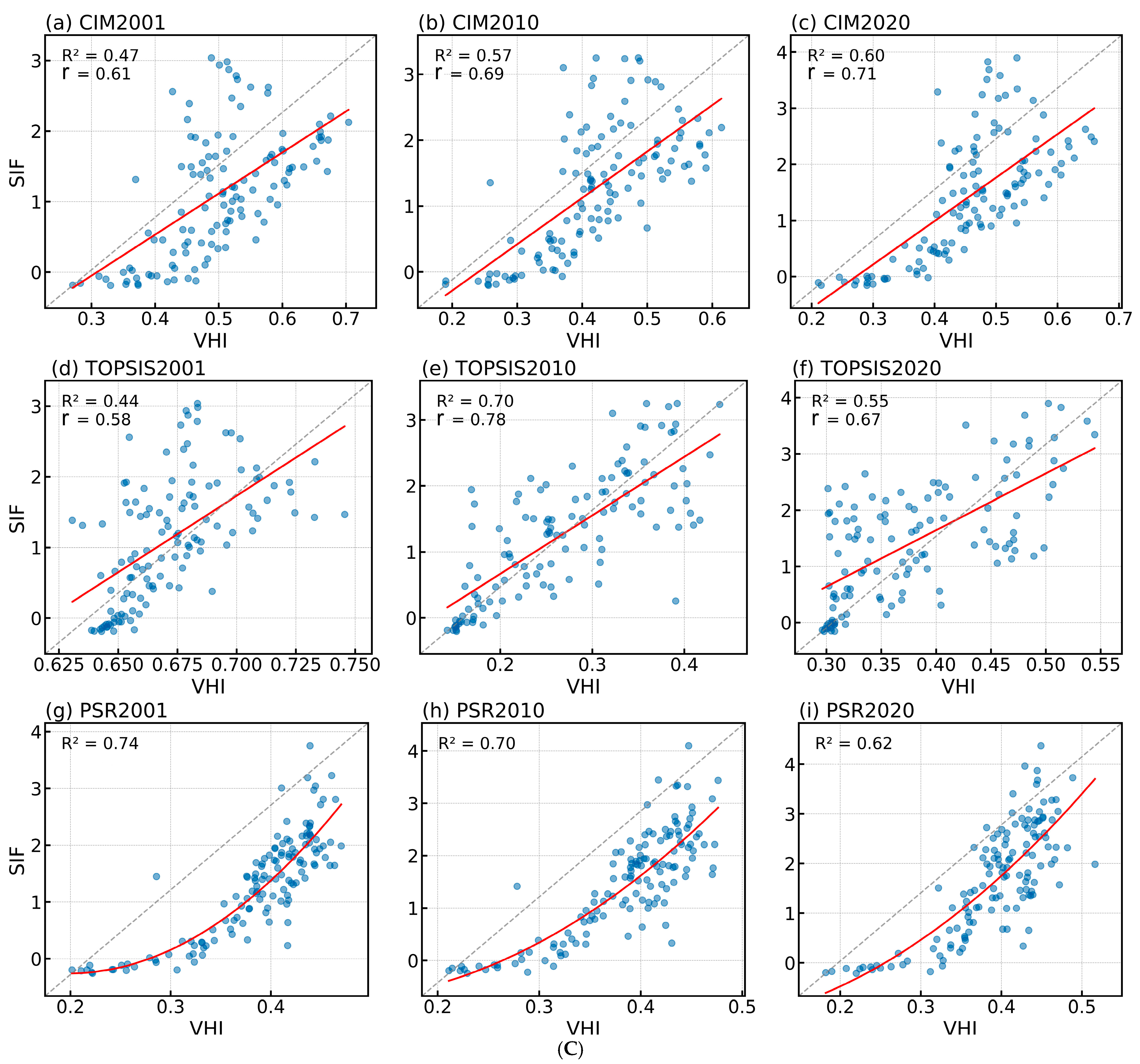
3.1. PSR Models Most Accurately Reflect Vegetation Health
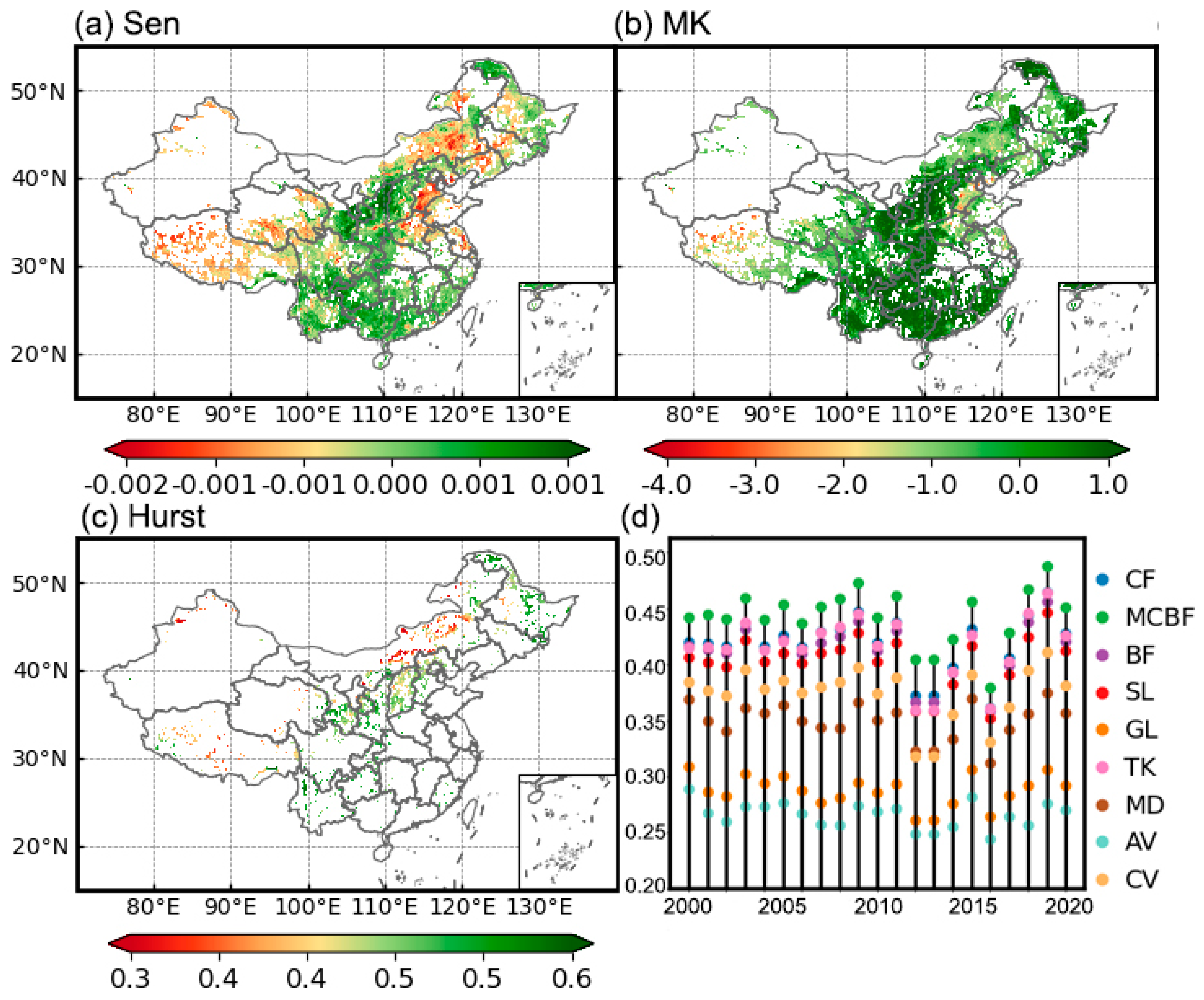
3.1.1. Temporal Trends of Vegetation Health
3.1.2. Evaluation of the Vegetation Health Assessment System Construction Effectiveness
3.1.3. Trends in Vegetation Health
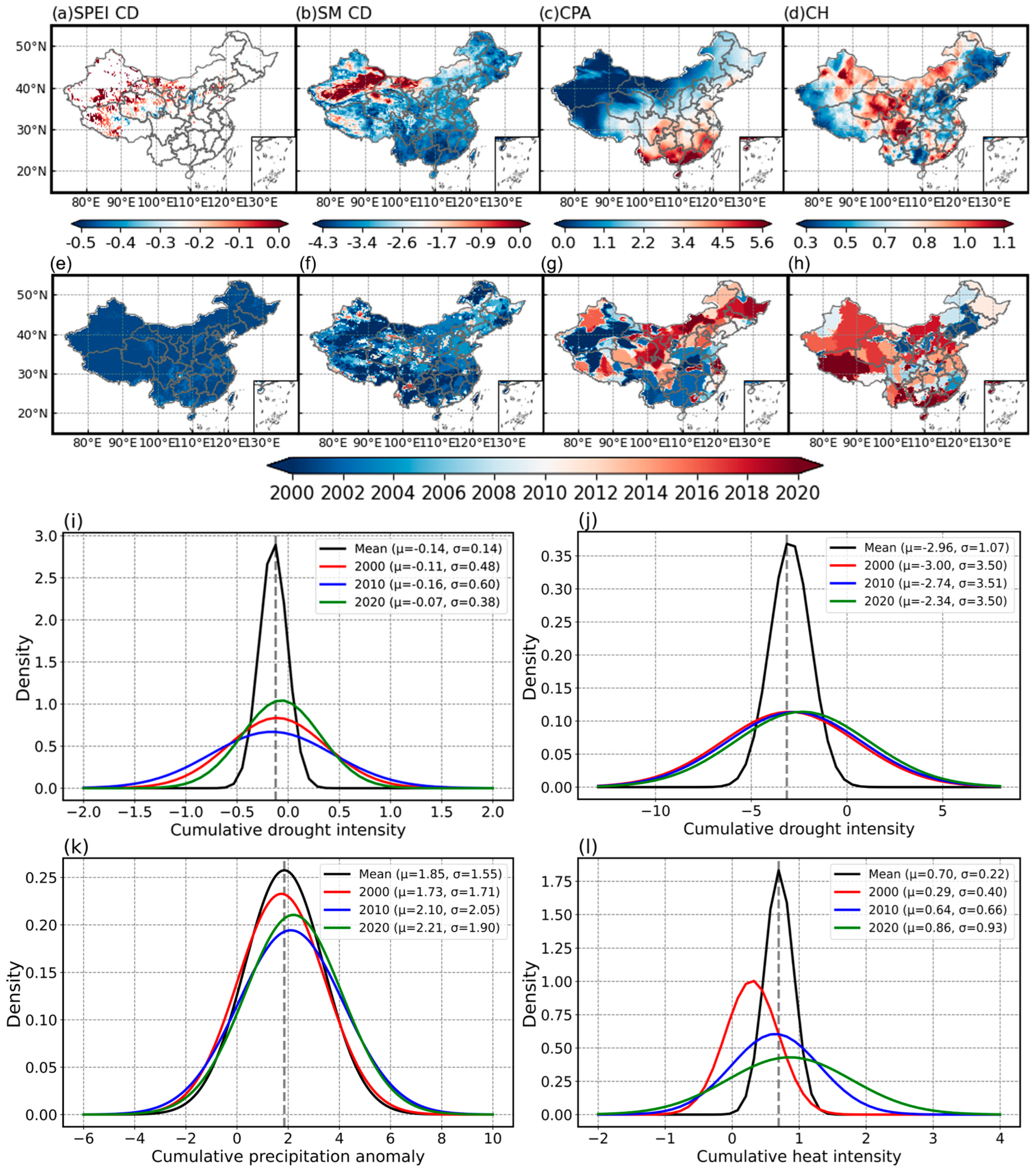
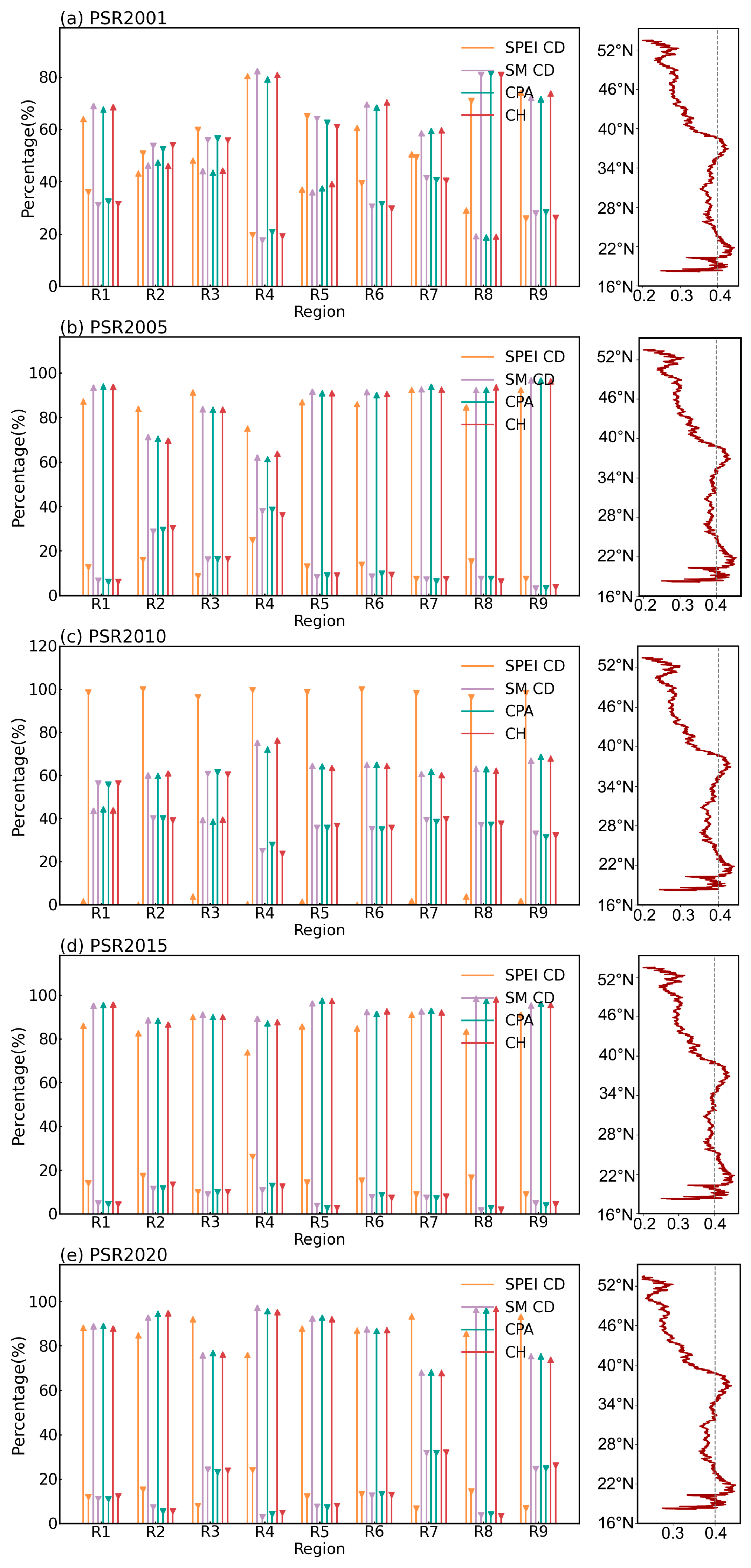
3.2. Characteristics of Stress Events
3.3. Responses of Vegetation Health to Stress Events
3.3.1. Anomalies in Vegetation Health Due to Stress Events
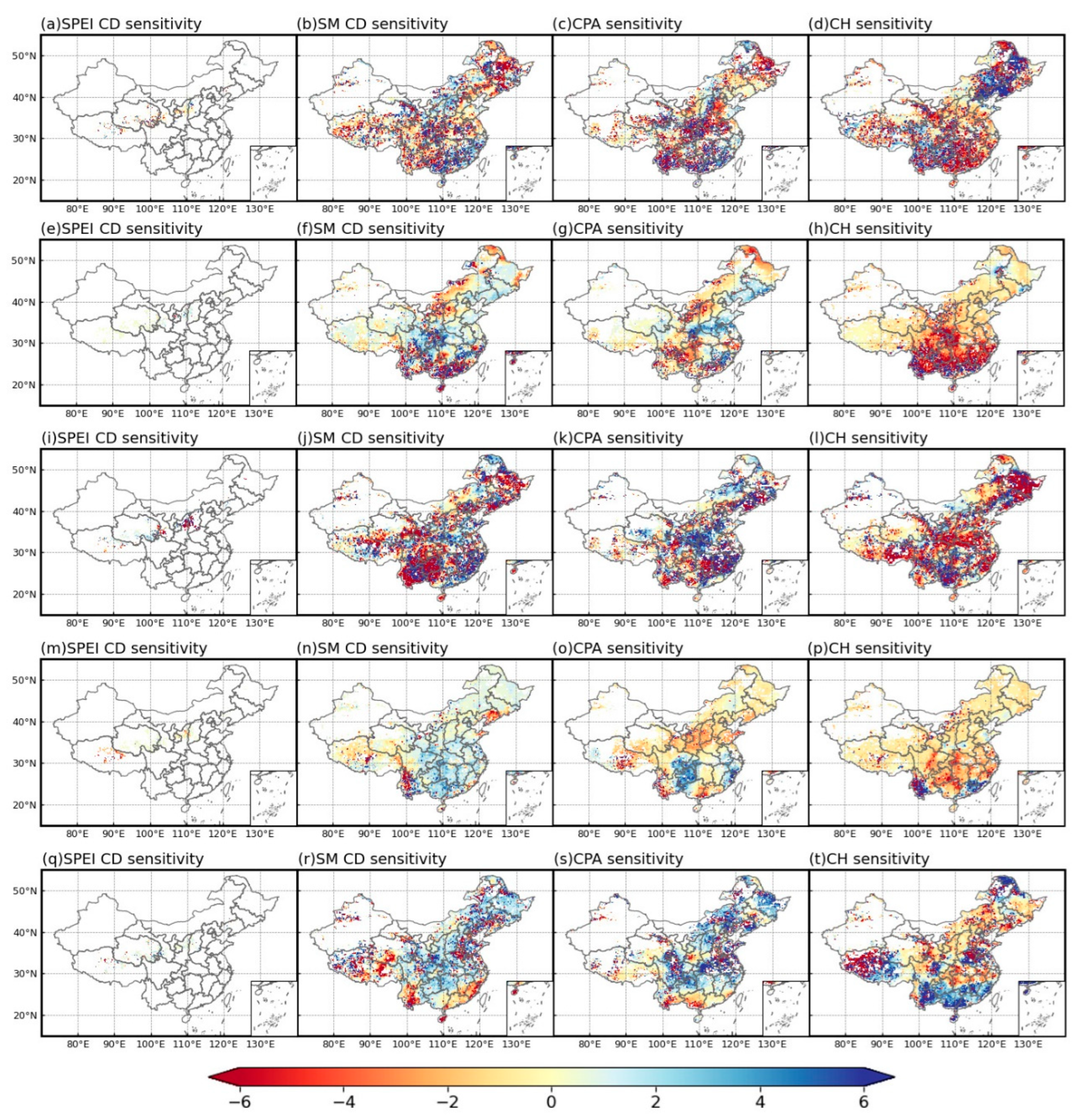
3.3.2. Vegetation Health Dominated by Negative Sensitivity to Stress Events
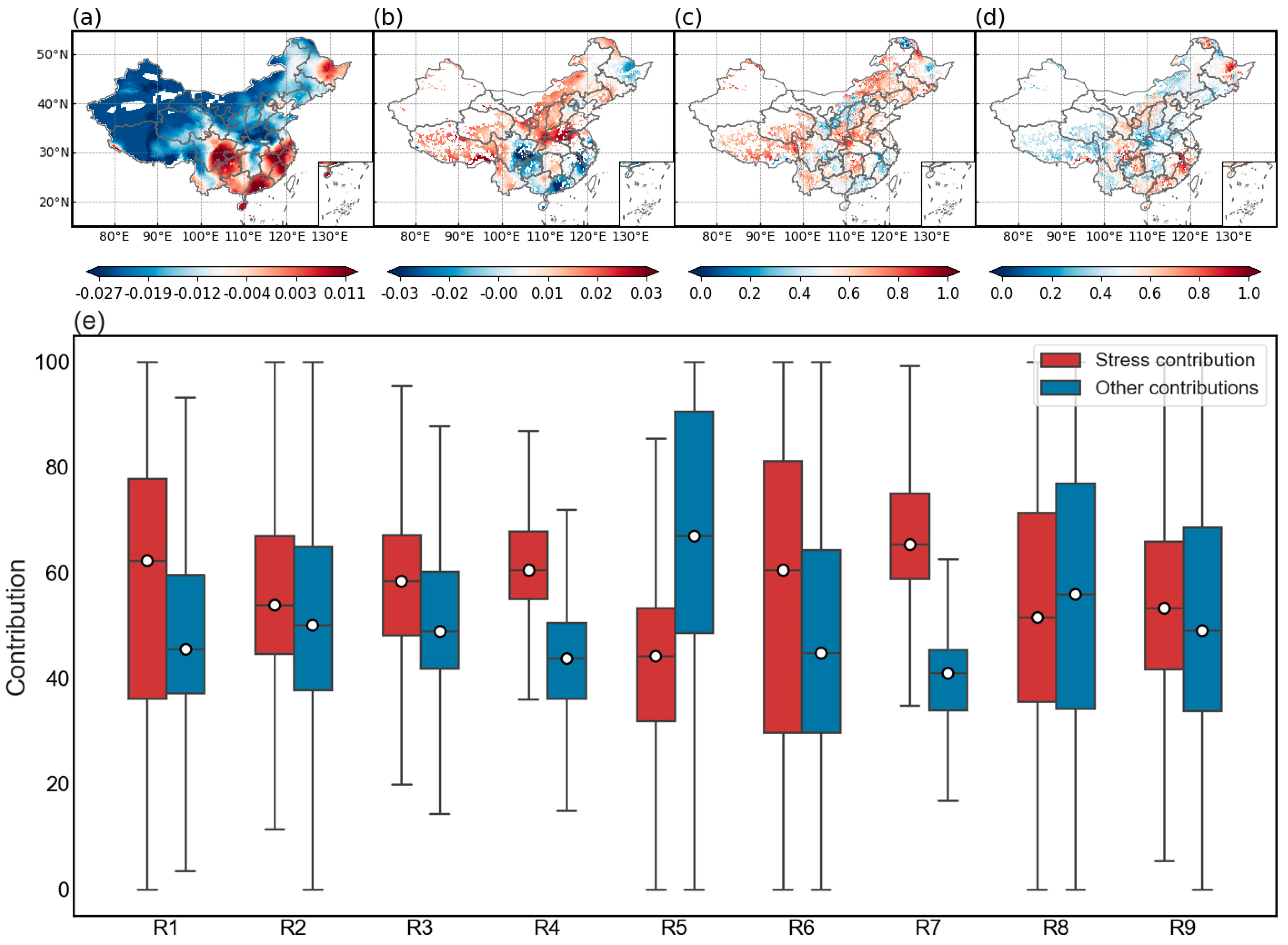
3.4. Stress Events Have a Greater Impact on Vegetation Health Compared to Other Factors
4. Discussion
4.1. Evaluation of the Effectiveness and Trend Changes of Vegetation Health Assessment Systems
4.2. Characteristics of Stress Events and Their Impact on Vegetation Health
4.3. Uncertainty Analysis
5. Conclusions
- (1)
- VHI levels in eastern China are higher than in the western regions, and they exhibit a significant upward trend as a whole. The frequency of drought stress occurring alongside heat stress is gradually increasing, but there is a trend toward a warmer and wetter climate with climate change becoming increasingly extreme.
- (2)
- Among the vegetation types, meadows, grasslands, and cultivated vegetation are most sensitive to stress events with CH having a greater impact on vegetation health than other stress events. Regionally, vegetation health is most affected by stress events in areas R4 and R8 with a significant decline in vegetation health occurring at latitudes greater than 35°.
- (3)
- The contribution of stress events to vegetation health is predominant and may continue to increase in the future.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kogan, F.; Yang, B.; Wei, G.; Pei, Z.; Jiao, X. Modelling Corn Production in China Using AVHRR-Based Vegetation Health Indices. Int. J. Remote Sens. 2005, 26, 2325–2336. [Google Scholar] [CrossRef]
- Pei, F.; Wu, C.; Liu, X.; Xia, L.; Yang, K.; Zhou, Y.; Wang, K.; Xu, L.; Xia, G. Monitoring the Vegetation Activity in China Using Vegetation Health Indices. Agric. For. Meteorol. 2018, 248, 215–227. [Google Scholar] [CrossRef]
- Calvin, K.; Lee, H.; Romero, J.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.; Trisos, C.; Aldunce, P.; Barret, K.; et al. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023. [Google Scholar] [CrossRef]
- Perkins-Kirkpatrick, S.E.; Lewis, S.C. Increasing Trends in Regional Heatwaves. Nat. Commun. 2020, 11, 3357. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, X.; Wang, K.; Ciais, P.; Tang, S.; Jin, L.; Li, L.; Piao, S. Responses of Vegetation Greenness and Carbon Cycle to Extreme Droughts in China. Agric. For. Meteorol. 2021, 298–299, 108307. [Google Scholar] [CrossRef]
- Ghanbari, M.; Arabi, M.; Georgescu, M.; Broadbent, A.M. The Role of Climate Change and Urban Development on Compound Dry-Hot Extremes across US Cities. Nat. Commun. 2023, 14, 3509. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Piao, S.; Huntingford, C.; Peñuelas, J.; Yang, H.; Xu, H.; Chen, A.; Friedlingstein, P.; Keenan, T.F.; Sitch, S.; et al. Global Variations in Critical Drought Thresholds That Impact Vegetation. Natl. Sci. Rev. 2023, 10, nwad049. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, D.; Chen, G.; Yang, J.; Yang, Z.; Guo, X.J.; Wang, C.; Zhu, S.; Huang, Y.; Chen, H.; et al. Responses of Leaf Functional Traits to Different Hydrological Regimes and Leaf Economics Spectrum in the Water Level Fluctuation Zone of Three Gorges Reservoir, China. Front. Plant Sci. 2022, 13, 939452. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Zhang, X.; Chen, A.; Liu, Q.; Lian, X.; Wang, X.; Peng, S.; Wu, X. The Impacts of Climate Extremes on the Terrestrial Carbon Cycle: A Review. Sci. China Earth Sci. 2019, 62, 1551–1563. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, Z.; Wang, L.; Jin, B. Plant Responses to Heat Stress: Physiology, Transcription, Noncoding RNAs, and Epigenetics. Int. J. Mol. Sci. 2020, 22, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, D. Dynamic Assessment of Alpine Pasture Grassland Ecosystem Health: A Case Study from the Gannan Region. Pratacultural Sci. 2017, 34, 16–29. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Zhou, G.; Luo, T.; Yakir, D.; Zhou, L.; Lv, X. Variation of Net Primary Productivity and Its Drivers in China’s Forests during 2000–2018. For. Ecosyst. 2020, 7, 15. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Yang, H.; Zhao, Y. Study of the Temporal and Spatial Patterns of Drought in the Yellow River Basin Based on SPEI. Sci. China Earth Sci. 2018, 61, 1098–1111. [Google Scholar] [CrossRef]
- Li, X.; Meng, X.; Ji, X.; Zhou, J.; Pan, C.; Gao, N. Zoning Technology for the Management of Ecological and Clean Small-Watersheds via k-Means Clustering and Entropy-Weighted TOPSIS: A Case Study in Beijing. J. Clean. Prod. 2023, 397, 136449. [Google Scholar] [CrossRef]
- Fathi-Taperasht, A.; Shafizadeh-Moghadam, H.; Kouchakzadeh, M. MODIS-based evaluation of agricultural drought, water use efficiency, and post-drought in Iran: Considering the influence of heterogeneous climatic regions. J. Clean. Prod. 2022, 374, 133836. [Google Scholar] [CrossRef]
- Funk, C.; Dettinger, M.D.; Michaelsen, J.C.; Verdin, J.P.; Brown, M.E.; Barlow, M.; Hoell, A. Warming of the Indian Ocean Threatens Eastern and Southern African Food Security but Could Be Mitigated by Agricultural Development. Proc. Natl. Acad. Sci. USA 2008, 105, 11081–11086. [Google Scholar] [CrossRef] [PubMed]
- Rowhani, P.; Lobell, D.B.; Linderman, M.; Ramankutty, N. Climate Variability and Crop Production in Tanzania. Agric. For. Meteorol. 2011, 151, 449–460. [Google Scholar] [CrossRef]
- Sutanto, S.J.; Vitolo, C.; Di Napoli, C.; D’Andrea, M.; Van Lanen, H.A.J. Heatwaves, Droughts, and Fires: Exploring Compound and Cascading Dry Hazards at the Pan-European Scale. Environ. Int. 2020, 134, 105276. [Google Scholar] [CrossRef]
- Sauter, C.; Fowler, H.J.; Westra, S.; Ali, H.; Peleg, N.; White, C.J. Compound Extreme Hourly Rainfall Preconditioned by Heatwaves Most Likely in the Mid-Latitudes. Weather. Clim. Extrem. 2023, 40, 100563. [Google Scholar] [CrossRef]
- Schweiger, R.; Maidel, A.-M.; Renziehausen, T.; Schmidt-Schippers, R.; Müller, C. Effects of Drought, Subsequent Waterlogging and Redrying on Growth, Physiology and Metabolism of Wheat. Physiol. Plant. 2023, 175, e13874. [Google Scholar] [CrossRef] [PubMed]
- Marengo, J.A.; Ambrizzi, T.; Alves, L.M.; Barreto, N.J.C.; Reboita, M.S.; Ramos, A.M. Changing Trends in Rainfall Extremes in the Metropolitan Area of São Paulo: Causes and Impacts. Front. Clim. 2020, 2, 3. [Google Scholar] [CrossRef]
- Yu, F.; Price, K.P.; Ellis, J.; Shi, P. Response of Seasonal Vegetation Development to Climatic Variations in Eastern Central Asia. Remote Sens. Environ. 2003, 87, 42–54. [Google Scholar] [CrossRef]
- Yu, Y.; You, Q.; Zuo, Z.; Zhang, Y.; Cai, Z.; Li, W.; Jiang, Z.; Ullah, S.; Tang, X.; Zhang, R.; et al. Compound Climate Extremes in China: Trends, Causes, and Projections. Atmos. Res. 2023, 286, 106675. [Google Scholar] [CrossRef]
- Smith, T.; Traxl, D.; Boers, N. Empirical Evidence for Recent Global Shifts in Vegetation Resilience. Nat. Clim. Chang. 2022, 12, 477–484. [Google Scholar] [CrossRef]
- The State Council of the People’s Republic of China. Notice from the General Office of the State Council on Printing and Distributing the “14th Five-Year Plan” Employment Promotion Plan. The Central People’s Government of the People’s Republic of China. Available online: https://www.gov.cn/xinwen/2021-08/19/content_5632124.htm (accessed on 6 May 2024).
- Wu, C.; Chen, W. Indicator System Construction and Health Assessment of Wetland Ecosystem—Taking Hongze Lake Wetland, China as an Example. Ecol. Indic. 2020, 112, 106164. [Google Scholar] [CrossRef]
- Zhu, X.; Jiao, L.; Wu, X.; Du, D.; Wu, J.; Zhang, P. Ecosystem Health Assessment and Comparison of Natural and Constructed Wetlands in the Arid Zone of Northwest China. Ecol. Indic. 2023, 154, 110576. [Google Scholar] [CrossRef]
- Guan, Y.; Lu, H.; Yin, C.; Xue, Y.; Jiang, Y.; Kang, Y.; He, L.; Heiskanen, J. Vegetation Response to Climate Zone Dynamics and Its Impacts on Surface Soil Water Content and Albedo in China. Sci. Total Environ. 2020, 747, 141537. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, G.; Sharifzadeh, M.S.; Sklenička, P.; Azadi, H. Adaptive Capacity of Farming Systems to Climate Change in Iran: Application of Composite Index Approach. Agric. Syst. 2023, 204, 103537. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, J.; Guan, J.; Han, W.; Liu, Y. Grassland Cover Dynamics and Their Relationship with Climatic Factors in China from 1982 to 2021. Sci. Total Environ. 2023, 905, 167067. [Google Scholar] [CrossRef]
- Qu, L.; Chen, J.; Dong, G.; Jiang, S.; Li, L.; Guo, J.; Shao, C. Heat Waves Reduce Ecosystem Carbon Sink Strength in a Eurasian Meadow Steppe. Environ. Res. 2016, 144, 39–48. [Google Scholar] [CrossRef]
- Sen, P.K. Estimates of the Regression Coefficient Based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric Tests against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, X.; Zhou, T.; Zhang, W.; Hu, S.; Clark, R. Understanding and Attribution of Extreme Heat and Drought Events in 2022: Current Situation and Future Challenges. Adv. Atmos. Sci. 2023, 40, 1941–1951. [Google Scholar] [CrossRef]
- Leng, S.; Huete, A.; Cleverly, J.; Lu, X.; Ma, X.; Gao, S.; Yu, Q. Response of Dryland Vegetation under Extreme Wet Events with Satellite Measures of Greenness and Fluorescence. Sci. Total Environ. 2022, 842, 156860. [Google Scholar] [CrossRef]
- Xu, W.; Yuan, W.; Wu, D.; Zhang, Y.; Shen, R.; Xia, X.; Ciais, P.; Liu, J. Impacts of Record-Breaking Compound Heatwave and Drought Events in 2022 China on Vegetation Growth. Agric. For. Meteorol. 2024, 344, 109799. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Q.; Liu, R.; Zhao, Y.; Zhang, D. Effects of Climate Change and Human Activities on Vegetation Coverage Change in Northern China Considering Extreme Climate and Time-Lag and -Accumulation Effects. Sci. Total Environ. 2023, 860, 160527. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K. Regression, Heredity, and Panmixia. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 1896, 187, 253–318. [Google Scholar] [CrossRef]
- Qi, J.; Kerr, Y.H.; Moran, M.S.; Weltz, M.; Huete, A.R.; Sorooshian, S.; Bryant, R. Leaf Area Index Estimates Using Remotely Sensed Data and BRDF Models in a Semiarid Region. Remote Sens. Environ. 2000, 73, 18–30. [Google Scholar] [CrossRef]
- Fang, H.; Baret, F.; Plummer, S.; Schaepman-Strub, G. An overview of global leaf area index (LAI): Methods, products, validation, and applications. Rev. Geophys. 2019, 57, 739–799. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Zhang, K.; Cheng, X.; Band, L.E.; Zhang, Q. Effects of Land Use/Land Cover and Climate Changes on Terrestrial Net Primary Productivity in the Yangtze River Basin, China, from 2001 to 2010. J. Geophys. Res. Biogeosci. 2014, 119, 1092–1109. [Google Scholar] [CrossRef]
- Mohammed, G.H.; Colombo, R.; Middleton, E.M.; Rascher, U.; van der Tol, C.; Nedbal, L.; Goulas, Y.; Pérez-Priego, O.; Damm, A.; Meroni, M.; et al. Remote sensing of solar-induced chlorophyll fluorescence (SIF) in vegetation: 50 years of progress. Remote Sens. Environ. 2019, 231, 111177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ren, Y.; Cao, L.; Wu, J.; Zhang, S.; Hu, C.; Zhujie, S. Characteristics of Dry-Wet Climate Change in China during the Past 60 Years and Its Trends Projection. Atmosphere 2022, 13, 275. [Google Scholar] [CrossRef]
- Bakala, H.S.; Devi, J.; Singh, G.; Singh, I. Drought and Heat Stress: Insights into Tolerance Mechanisms and Breeding Strategies for Pigeonpea Improvement. Planta 2024, 259, 123. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, Q.; Chen, Y.; Zhou, H.; Xiang, Y.; Liu, Z.; Hou, Y. Vegetation Change and Eco-Environmental Quality Evaluation in the Loess Plateau of China from 2000 to 2020. Remote Sens. 2023, 15, 424. [Google Scholar] [CrossRef]
- Lesk, C.; Anderson, W.; Rigden, A.; Coast, O.; Jägermeyr, J.; McDermid, S.; Davis, K.F.; Konar, M. Compound Heat and Moisture Extreme Impacts on Global Crop Yields under Climate Change. Nat. Rev. Earth Environ. 2022, 3, 872–889. [Google Scholar] [CrossRef]
- Alizadeh, M.R.; Adamowsk, J.; Nikoo, M.R.; AghaKouchak, A.; Dennison, P.; Sadegh, M. A Century of Observations Reveals Increasing Likelihood of Continental-Scale Compound Dry-Hot Extremes. Sci. Adv. 2020, 6, eaaz4571. [Google Scholar] [CrossRef] [PubMed]
- Fowler, H.J.; Lenderink, G.; Prein, A.F.; Westra, S.; Allan, R.P.; Ban, N.; Barbero, R.; Berg, P.; Blenkinsop, S.; Do, H.X.; et al. Anthropogenic Intensification of Short-Duration Rainfall Extremes. Nat. Rev. Earth Environ. 2021, 2, 107–122. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Y.; Hu, T.; Yin, H. The 2022 Record-Breaking Heat Event over the Middle and Lower Reaches of the Yangtze River: The Role of Anthropogenic Forcing and Atmospheric Circulation. Bull. Am. Meteorol. Soc. 2024, 105, E200–E205. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding Plant Responses to Drought—From Genes to the Whole Plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.; Chen, M.; Xu, X.; Wang, Z.; Wang, C.; Piao, S.; Lin, W.; Miao, G.; Deng, M.; et al. Responses of Terrestrial Ecosystems to Temperature and Precipitation Variability in the Northern Hemisphere. Glob. Chang. Biol. 2018, 24, 5093–5106. [Google Scholar] [CrossRef]
- Grossiord, C.; Granier, A.; Ratcliffe, S.; Bouriaud, O.; Bruelheide, H.; Chećko, E.; Forrester, D.I.; Dawud, S.M.; Finér, L.; Pollastrini, M.; et al. Tree Diversity Does Not Always Improve Resistance of Forest Ecosystems to Drought. Proc. Natl. Acad. Sci. USA 2014, 111, 14812–14817. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, C.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368, eaaz7005. [Google Scholar] [CrossRef] [PubMed]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, Y.; Fu, C.; Chen, F.; Fu, Q.; Dai, A.; Shinoda, M.; Ma, Z.; Guo, W.; Li, Z.; et al. Dryland climate change: Recent progress and challenges. Rev. Geophys. 2017, 55, 719–778. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, G.; Zheng, Z.; Zhu, Y.; Zhu, J.; Di, Y.; Gao, J.; Cai, M.; Zhang, Y. Wetting-warming climate increases ecosystem carbon use efficiency of Chinese arid and semi-arid ecosystem. Agric. For. Meteorol. 2024, 351, 110018. [Google Scholar] [CrossRef]
- Su, J.; Zhang, Y.; Xu, F. Divergent responses of grassland productivity and plant diversity to intra-annual precipitation variability across climate regions: A global synthesis. J. Ecol. 2023, 111, 1921–1934. [Google Scholar] [CrossRef]
- Verrall, B.; Pickering, C.M. Alpine vegetation in the context of climate change: A global review of past research and future directions. Sci. Total Environ. 2020, 748, 141344. [Google Scholar] [CrossRef]
- Zahra, N.; Hafeez, M.B.; Ghaffar, A.; Kausar, A.; Al Zeidi, M.; Siddique, K.H.M.; Farooq, M. Plant photosynthesis under heat stress: Effects and management. Environ. Exp. Bot. 2023, 206, 105178. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and Landscape Dynamics in a Changing World. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Wang, L. Differential Responses of Vegetation Phenology to Climatic Elements during Extreme Events on the Chinese Loess Plateau. Sci. Total Environ. 2024, 933, 173146. [Google Scholar] [CrossRef]
- Oswald-Spring, D.Ú. Climate Change 2014: Impacts, Adaptation, and Vulnerability; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.; Austrheim, G.; Speed, J.D.M.; Zhao, W.; Cherubini, F. Disentangling the Effects of Climate Variability and Herbivory on Spatial and Temporal Changes in Northern Vegetation Greening. Ecol. Indic. 2024, 159, 111700. [Google Scholar] [CrossRef]
- Fathi-Taperasht, A.; Shafizadeh-Moghadam, H.; Sadian, A.; Xu, T.; Nikoo, M.R. Drought-induced vulnerability and resilience of different land use types using time series of MODIS-based indices. Int. J. Disaster Risk Reduct. 2023, 91, 103703. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Type | Spatial Resolution | Temporal Resolution | Source |
|---|---|---|---|
| Temperature | 0.0083333° | Yearly | http://www.geodata.cn/data/datadetails.html?dataguid=67669514169502&docId=4 (accessed on 12 March 2024) |
| Precipitation | https://www.geodata.cn/data/datadetails.html?dataguid=113786088533256&docId=6 (accessed on 12 March 2024) | ||
| 2 m temperature | 0.1° | Monthly | https://cds.climate.copernicus.eu/datasets/reanalysis-era5-land-monthly-means?tab=overview (accessed on 12 March 2024) |
| GPP | 500 m | 8 day | https://lpdaac.usgs.gov/products/mod17a2hv061/ (accessed on 6 May 2023) |
| Land Surface Temperature, SR, RH | 0.1° | Monthly | https://cds.climate.copernicus.eu/#!/home (accessed on 15 March 2024) |
| ET, SM | 0.25° | Yearly | https://www.gleam.eu/ (accessed on 17 March 2024) |
| Slope, DEM | 1 km | Yearly | https://www.gscloud.cn/ (accessed on 18 March 2024) |
| PM2.5 | 1 km | Monthly | https://data.tpdc.ac.cn/ (accessed on 18 March 2024) |
| PDSI | 1 km | Monthly | https://crudata.uea.ac.uk/cru/data/drought/#global (accessed on 18 March 2024) |
| Nighttime Light | 500 m | Yearly | http://nnu.geodata.cn/data/datadetails.html?dataguid=8213124601985&docid=0 (accessed on 20 March 2024) |
| Soil properties | 1 km | Yearly | https://data.tpdc.ac.cn/zh-hans/data/8ba0a731-5b0b-4e2f-8b95-8b29cc3c0f3a (accessed on 20 March 2024) |
| GDP, Tertiary industry added value | 1 km | Yearly | https://github.com/thestarlab/ChinaGDP (accessed on 25 March 2024) |
| Population density | 1 km | Yearly | https://landscan.ornl.gov (accessed on 25 March 2024) |
| NDVI | 1 km | Yearly | https://www.geodata.cn/main/#/face_science_detail?gugu=197351408897313 (accessed on 30 March 2024) |
| LAI | 0.05° | Yearly | https://doi.org/10.3974/geodb.2023.10.03.V1 (accessed on 30 March 2024) |
| SIF | 0.05° | 4 day | https://cstr.cn/18406.11.Ecolo.tpdc.271751 (accessed on 3 April 2024) |
| NPP | 500 m | Yearly | https://lpdaac.usgs.gov/product_search/?view=listhttpl://lpdaac.usgs.gov/product_search/?view=list (accessed on 3 April 2024) |
| 1:1,000,000 vegetation types in China | 1 km | https://www.plantplus.cn (accessed on 5 April 2024) |
| System | Subsystem | Primary Indicator Layer | Secondary Indicator Layer | Attributes |
|---|---|---|---|---|
| China Vegetation Health System | Pressure | Human activity pressure | Population Density | − |
| Climate pressure | Relative Humidity | + | ||
| Temperature | − | |||
| Precipitation | + | |||
| PDSI | − | |||
| SM | + | |||
| State | Vegetation growth state | NDVI | + | |
| LAI | + | |||
| Soil attributes state | PH | − | ||
| SOM | + | |||
| Total Nitrogen | + | |||
| Total Phosphorus | + | |||
| Terrain state | Slope | − | ||
| DEM | − | |||
| Response | Vegetation response | FVC | + | |
| GPP | + | |||
| Transpiration | − | |||
| Production activity response | Tertiary Industry Value Added | − | ||
| Nighttime Lights | − | |||
| PM2.5 | − | |||
| GDP | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, P.; Peng, J.; Zheng, J.; Liu, L.; Yu, X.; Li, W. Vegetation Health in China Is Severely Compromised by Drought, Wet and Heat Stress Events. Forests 2025, 16, 38. https://doi.org/10.3390/f16010038
Ma P, Peng J, Zheng J, Liu L, Yu X, Li W. Vegetation Health in China Is Severely Compromised by Drought, Wet and Heat Stress Events. Forests. 2025; 16(1):38. https://doi.org/10.3390/f16010038
Chicago/Turabian StyleMa, Ping, Jian Peng, Jianghua Zheng, Liang Liu, Xiaojing Yu, and Wei Li. 2025. "Vegetation Health in China Is Severely Compromised by Drought, Wet and Heat Stress Events" Forests 16, no. 1: 38. https://doi.org/10.3390/f16010038
APA StyleMa, P., Peng, J., Zheng, J., Liu, L., Yu, X., & Li, W. (2025). Vegetation Health in China Is Severely Compromised by Drought, Wet and Heat Stress Events. Forests, 16(1), 38. https://doi.org/10.3390/f16010038